
Leptin—A Potential Bridge between Fat Metabolism and the Brain’s Vulnerability to Neuropsychiatric Disorders: A Systematic Review



Abstract
:1. Introduction
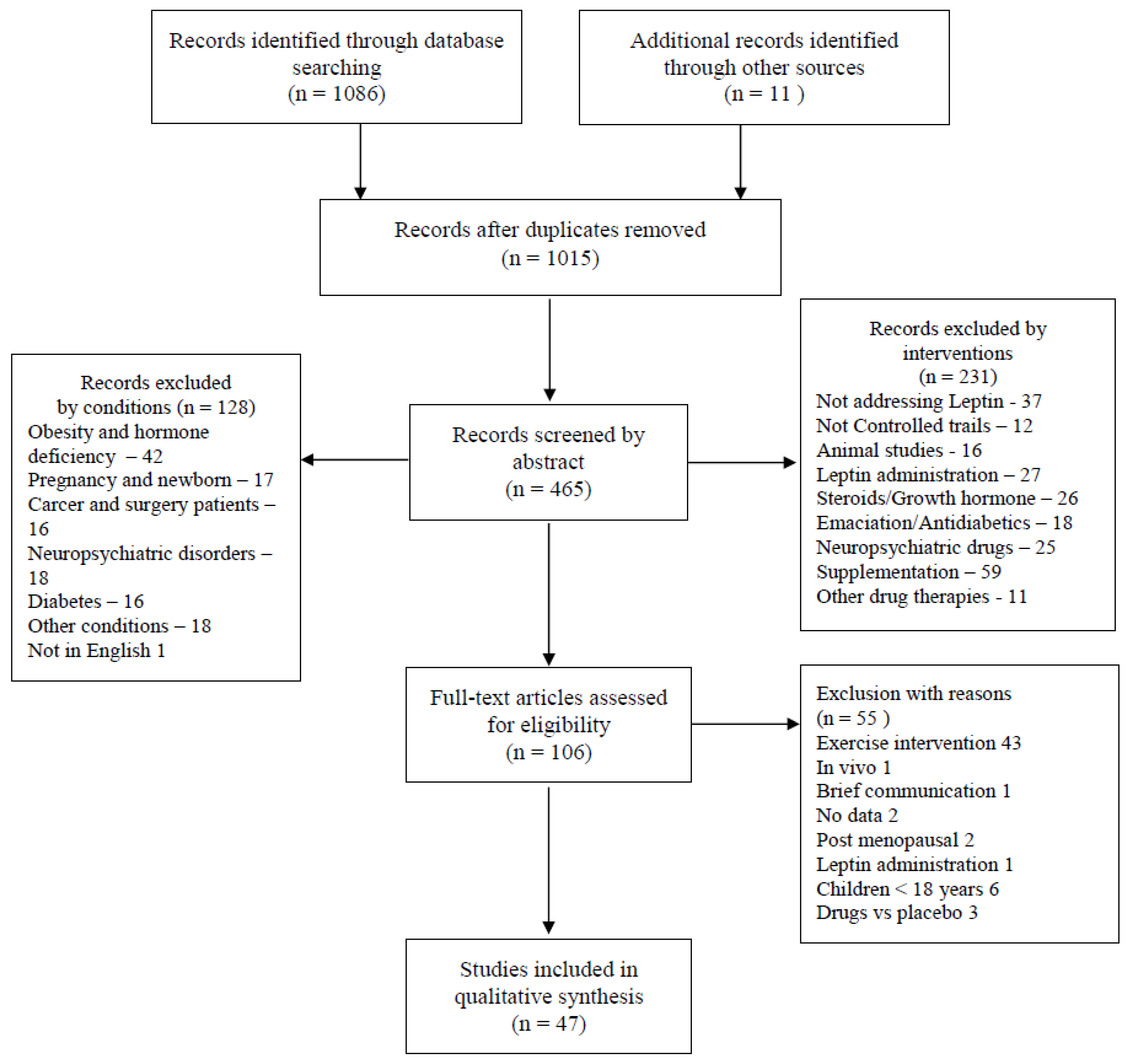
2. Methods
3. Results
4. Discussion
5. Final Considerations
Author Contributions
Funding
Conflicts of Interest
References
- Freed, C. Psychic factors in the development and treatment of obesity. J. Am. Med. Assoc. 1947, 133, 369–373. [Google Scholar] [CrossRef]
- Bouchard, C. Gene-environment interactions in the etiology of obesity: Defining the fundamentals. Obesity 2008, 16, 5–10. [Google Scholar] [CrossRef]
- Chung, W.K.; Leibel, R.L. Considerations regarding the genetics of obesity. Obesity 2008, 16, 33–39. [Google Scholar] [CrossRef] [Green Version]
- Hinney, A.; Hebebrand, J. Polygenic obesity in humans. Obes. Facts 2008, 1, 35–42. [Google Scholar] [CrossRef]
- Kleinendorst, L.; van Haelst, M.M.; van den Akker, E.L.T. Genetics of Obesity. Exp. Suppl. 2019, 111, 419–441. [Google Scholar] [CrossRef]
- Shuldiner, A.R. Obesity genes and gene-environment-behavior interactions: Recommendations for a way forward. Obesity 2008, 16, 79–81. [Google Scholar] [CrossRef] [Green Version]
- Wareham, N.J.; Young, E.H.; Loos, R.J.F. Epidemiological study designs to investigate gene-behavior interactions in the context of human obesity. Obesity 2008, 16, 66–71. [Google Scholar] [CrossRef] [Green Version]
- Leibel, R.L. Energy In, Energy Out, and the Effects of Obesity-Related Genes. N. Engl. J. Med. 2008, 359, 2603–2604. [Google Scholar] [CrossRef]
- Warden, C.H.; Fisler, J.S. Gene-nutrient and gene-physical activity summary-genetics viewpoint. Obesity 2008, 16, 55–59. [Google Scholar] [CrossRef]
- Capers, P.L.; Fobian, A.D.; Kaiser, K.A.; Borah, R.; Allison, D.B. A systemic review and meta-analysis of randomized controlled trials of the impact of sleep duration on adiposity and components of energy balance. Obes. Rev. 2015, 16, 771–782. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Considine, R.V. Leptin and obesity in humans. Eat. Weight. Disord. Stud. Anorex. Bulim. Obes. 1997, 2, 61–66. [Google Scholar] [CrossRef]
- Friedman, J.M.; Leibel, R.L.; Siegel, D.S.; Walsh, J.; Bahary, N. Molecular mapping of the mouse ob mutation. Genomics 1991, 11, 1054–1062. [Google Scholar] [CrossRef]
- St-Onge, M.P. The Role of Sleep Duration in the Regulation of Energy balance: Effects on Energy intakes and Expenditure. J. Clin. Sleep Med. 2013, 9, 73–80. [Google Scholar] [CrossRef]
- Harter, C.J.L.; Kavanagh, G.S.; Smith, J.T. The role of kisspeptin neurons in reproduction and metabolism. J. Endocrinol. 2018, 238, 173–183. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- La Cava, A. Leptin in inflammation and autoimmunity. Cytokine 2017, 98, 51–58. [Google Scholar] [CrossRef] [PubMed]
- Mishra, S.; Harris, T.B.; Hsueh, W.C.; Hue, T.; Leak, T.S.; Li, R.; Mehta, M.; Vaisse, C.; Sahyoun, N.R. The association of serum leptin with mortality in older adults. PLoS ONE 2015, 10, e0140763. [Google Scholar] [CrossRef] [Green Version]
- Catteau, A.; Caillon, H.; Barrière, P.; Denis, M.G.; Masson, D.; Fréour, T. Leptin and its potential interest in assisted reproduction cycles. Hum. Reprod. Update 2016, 22, 320–341. [Google Scholar] [CrossRef] [Green Version]
- Chou, S.H.; Mantzoros, C. Role of leptin in human reproductive disorders. J. Endocrinol. 2014, 223, T49–T62. [Google Scholar] [CrossRef]
- He, S.; Le, N.A.; Ramirez-Zea, M.; Martorell, R.; Narayan, K.M.V.; Stein, A.D. Leptin partially mediates the association between early-life nutritional supplementation and long-term glycemic status among women in a Guatemalan longitudinal cohort. Am. J. Clin. Nutr. 2020, 111, 804–813. [Google Scholar] [CrossRef] [PubMed]
- Wauters, M.; Considine, R.; Löfgren, A.; Van Broeckhoven, C.; Van der Auwera, J.C.; De Leeuw, I.; Van Gaal, L. Associations of leptin with body fat distribution and metabolic parameters in non-insulin-dependent diabetic patients: No effect of apolipoprotein E polymorphism. Metab. Clin. Exp. 2000, 49, 724–730. [Google Scholar] [CrossRef]
- Da Silva, A.A.; do Carmo, J.M.; Hall, J.E. CNS Regulation of Glucose Homeostasis: Role of the Leptin-Melanocortin System. Curr. Diabetes Rep. 2020, 20, 29. [Google Scholar] [CrossRef]
- Fernández-Formoso, G.; Pérez-Sieira, S.; González-Touceda, D.; Dieguez, C.; Tovar, S. Leptin, 20 years of searching for glucose homeostasis. Life Sci. 2015, 140, 4–9. [Google Scholar] [CrossRef] [PubMed]
- Tartaglia, L.A. The leptin receptor. J. Biol. Chem. 1997, 272, 6093–6096. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caldeira, R.S.; Panissa VL, G.; Inoue, D.S.; Campos, E.Z.; Monteiro, P.A.; Giglio, B.d.M.; Pimentel, G.D.; Hofmann, P.; Lira, F.S. Impact to short-term high intensity intermittent training on different storages of body fat, leptin and soluble leptin receptor levels in physically active non-obese men: A pilot investigation. Clin. Nutr. ESPEN 2018, 28, 186–192. [Google Scholar] [CrossRef] [PubMed]
- Boutari, C.; Pappas, P.D.; Mintziori, G.; Nigdelis, M.P.; Athanasiadis, L.; Goulis, D.G.; Mantzoros, C.S. The effect of underweight on female and male reproduction. Metab. Clin. Exp. 2020, 107, 154–229. [Google Scholar] [CrossRef] [PubMed]
- Marazziti, D.; Rutigliano, G.; Baroni, S.; Landi, P.; Dell’Osso, L. Metabolic syndrome and major depression. CNS Spectrums. 2014, 19, 293–304. [Google Scholar] [CrossRef] [PubMed]
- Mintziori, G.; Nigdelis, M.P.; Mathew, H.; Mousiolis, A.; Goulis, D.G.; Mantzoros, C.S. The effect of excess body fat on female and male reproduction. Metab. Clin. Exp. 2020, 107, 154–193. [Google Scholar] [CrossRef] [PubMed]
- Stieg, M.R.; Sievers, C.; Farr, O.; Stalla, G.K.; Mantzoros, C.S. Leptin: A hormone linking activation of neuroendocrine axes with neuropathology. Psychoneuroendocrinology 2015, 51, 47–57. [Google Scholar] [CrossRef]
- Ángeles Martínez-García, M.; Moncayo, S.; Insenser, M.; Álvarez-Blasco, F.; Luque-Ramírez, M.; Escobar-Morreale, H.F. Postprandial responses of circulating energy homeostasis mediators to single macronutrient challenges: Influence of obesity and sex hormones. Food Funct. 2021, 12, 1051–1062. [Google Scholar] [CrossRef] [PubMed]
- Hamilton, K.; Harvey, J. The neuronal actions of leptin and the implications for treating alzheimer’s disease. Pharmaceuticals 2021, 14, 52. [Google Scholar] [CrossRef]
- Voigtmann, F.; Wolf, P.; Landgraf, K.; Stein, R.; Kratzsch, J.; Schmitz, S.; Abou Jamra, R.; Blüher, M.; Meiler, J.; Beck-Sickinger, A.G.; et al. Identification of a novel leptin receptor (LEPR) variant and proof of functional relevance directing treatment decisions in patients with morbid obesity. Metab. Clin. Exp. 2021, 116, 154438. [Google Scholar] [CrossRef]
- Javor, E.D.; Cochran, E.K.; Musso, C.; Young, J.R.; DePaoli, A.M.; Gorden, P. Long-term efficacy of leptin replacement in patients with generalized lipodystrophy. Diabetes 2005, 54, 1994–2002. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oral, E.A.; Simha, V.; Ruiz, E.; Andewelt, A.; Premkumar, A.; Snell, P.; Wagner, A.J.; DePaoli, A.M.; Reitman, M.L.; Taylor, S.I.; et al. Leptin-Replacement Therapy for Lipodystrophy. N. Engl. J. Med. 2002, 346, 570–578. [Google Scholar] [CrossRef] [PubMed]
- Cao, B.; Chen, Y.; Brietzke, E.; Cha, D.; Shaukat, A.; Pan, Z.; Park, C.; Subramaniapillai, M.; Zuckerman, H.; Grant, K.; et al. Leptin and adiponectin levels in major depressive disorder: A systematic review and meta-analysis. J. Affect. Disord. 2018, 238, 101–110. [Google Scholar] [CrossRef]
- Carvalho, A.F.; Rocha, D.Q.C.; McIntyre, R.S.; Mesquita, L.M.; Köhler, C.A.; Hyphantis, T.N.; Sales, P.M.G.; Machado-Vieira, R.; Berk, M. Adipokines as emerging depression biomarkers: A systematic review and meta-analysis. J. Psychiatr. Res. 2014, 59, 28–37. [Google Scholar] [CrossRef] [PubMed]
- Chan, J.L.; Heist, K.; DePaoli, A.M.; Veldhuis, J.D.; Mantzoros, C.S. The role of falling leptin levels in the neuroendocrine and metabolic adaptation to short-term starvation in healthy men. J. Clin. Investig. 2003, 111, 1409–1421. [Google Scholar] [CrossRef] [Green Version]
- Kiernan, K.; MacIver, N.J. The Role of the Adipokine Leptin in Immune Cell Function in Health and Disease. Front. Immunol. 2021, 11, 3656. [Google Scholar] [CrossRef] [PubMed]
- Moher, D.; Liberati, A.; Tetzlaff, J.; Altman, D.G.; Altman, D.; Antes, G.; Atkins, D.; Barbour, V.; Barrowman, N.; Berlin, J.A.; et al. Preferred reporting items for systematic reviews and meta-analyses: The PRISMA statement. PLoS Med. 2009, 6, e1000097. [Google Scholar] [CrossRef] [Green Version]
- Vgontzas, A.N.; Papanicolaou, D.A.; Bixler, E.O.; Hopper, K.; Lotsikas, A.; Lin, H.M.; Kales, A.; Chrousos, G.P. Sleep apnea and daytime sleepiness and fatigue: Relation to visceral obesity, insulin resistance, and hypercytokinemia. J. Clin. Endocrinol. Metab. 2000, 85, 1151–1158. [Google Scholar] [CrossRef]
- Guerci, B.; Hadjadj, S.; Quilliot, D.; Ziegler, O.; Drouin, P. No acute response of leptin to an oral fat load in obese patients and during circadian rhythm in healthy controls. Eur. J. Endocrinol. 2000, 143, 649–655. [Google Scholar] [CrossRef] [Green Version]
- Assali, A.R.; Ganor, A.; Beigel, Y.; Shafer, Z.; Hershcovici, T.; Fainaru, M. Insulin resistance in obesity: Body-weight or energy balance? J. Endocrinol. 2001, 171, 293–298. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsai, A.C.; Sandretto, A.; Chung, Y.C. Dieting is more effective in reducing weight but exercise is more effective in reducing fat during the early phase of a weight-reducing program in healthy humans. J. Nutr. Biochem. 2003, 14, 541–549. [Google Scholar] [CrossRef]
- Tentolouris, N.; Tsigos, C.; Perea, D.; Koukou, E.; Kyriaki, D.; Kitsou, E.; Daskas, S.; Daifotis, Z.; Makrilakis, K.; Raptis, S.A.; et al. Differential effects of high-fat and high-carbohydrate isoenergetic meals on cardiac autonomic nervous system activity in lean and obese women. Metab. Clin. Exp. 2003, 52, 1426–1432. [Google Scholar] [CrossRef]
- Matsumoto, T.; Miyatsuji, A.; Miyawaki, T.; Yanagimoto, Y.; Moritani, T. Potential association between endogenous leptin and sympatho-vagal activities in young obese Japanese women. Am. J. Hum. Biol. 2003, 15, 8–15. [Google Scholar] [CrossRef] [PubMed]
- Kassab, S.E.; Abdul-Ghaffar, T.; Nagalla, D.S.; Sachdeva, U.; Nayar, U. Interactions between leptin, neuropeptide-Y and insulin with chronic diurnal fasting during Ramadan. Ann. Saudi Med. 2004, 24, 345–349. [Google Scholar] [CrossRef]
- Eikelis, N.; Lambert, G.; Wiesner, G.; Kaye, D.; Schlaich, M.; Morris, M.; Hastings, J.; Socratous, F.; Esler, M. Extra-adipocyte leptin release in human obesity and its relation to sympathoadrenal function. Am. J. Physiol. Endocrinol. Metab. 2004, 286, E744–E752. [Google Scholar] [CrossRef]
- Baratta, R.; Amato, S.; Degano, C.; Farina, M.G.; Patanè, G.; Vigneri, R.; Frittitta, L. Adiponectin relationship with lipid metabolism is independent of body fat mass: Evidence from both cross-sectional and intervention studies. J. Clin. Endocrinol. Metab. 2004, 89, 2665–2671. [Google Scholar] [CrossRef] [Green Version]
- Weigle, D.S.; Breen, P.A.; Matthys, C.C.; Callahan, H.S.; Meeuws, K.E.; Burden, V.R.; Purnell, J.Q. A high-protein diet induces sustained reductions in appetite, ad libitum caloric intake, and body weight despite compensatory changes in diurnal plasma leptin and ghrelin concentrations. Am. J. Clin. Nutr. 2005, 82, 41–48. [Google Scholar] [CrossRef]
- Kiviniemi, T.O.; Snapir, A.; Saraste, M.; Toikka, J.O.; Raitakari, O.T.; Ahotupa, M.; Hartiala, J.J.; Scheinin, M.; Koskenvuo, J.W. Determinants of coronary flow velocity reserve in healthy young men. Am. J. Physiol. Heart Circ. Physiol. 2006, 291, 564–569. [Google Scholar] [CrossRef] [Green Version]
- Joosen, A.M.C.P.; Bakker, A.H.F.; Zorenc, A.H.G.; Kersten, S.; Schrauwen, P.; Westerterp, K.R. PPARγ activity in subcutaneous abdominal fat tissue and fat mass gain during short-term overfeeding. Int. J. Obes. 2006, 30, 302–307. [Google Scholar] [CrossRef] [Green Version]
- Chapelot, D.; Marmonier, C.; Aubert, R.; Allègre, C.; Gausseres, N.; Fantino, M.; Louis-Sylvestre, J. Consequence of omitting or adding a meal in man on body composition, food intake, and metabolism. Obesity 2006, 14, 215–227. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bouhlel, E.; Denguezli, M.; Zaouali, M.; Tabka, Z.; Shephard, R.J. Ramadan fasting’s effect on plasma leptin, adiponectin concentrations, and body composition in trained young men. Int. J. Sport Nutr. Exerc. Metab. 2008, 18, 617–627. [Google Scholar] [CrossRef]
- Bhutani, S.; Klempel, M.C.; Berger, R.A.; Varady, K.A. Improvements in coronary heart disease risk indicators by alternate-day fasting involve adipose tissue modulations. Obesity 2010, 18, 2152–2159. [Google Scholar] [CrossRef]
- Bergouignan, A.; Momken, I.; Schoeller, D.A.; Normand, S.; Zahariev, A.; Lescure, B.; Simon, C.; Blanc, S. Regulation of energy balance during long-term physical inactivity induced by bed rest with and without exercise training. J. Clin. Endocrinol. Metab. 2010, 95, 1045–1053. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, N.; Wang, Q.P.; Li, H.; Wu, X.P.; Sun, Z.Q.; Luo, X.H. Erratum to Relationships between serum adiponectin, leptin concentrations and bone mineral density and bone biochemical markers in Chinese. Clin. Chim. Acta 2010, 411, 1159. [Google Scholar] [CrossRef]
- Benedict, C.; Shostak, A.; Lange, T.; Brooks, S.J.; Schiöth, H.B.; Schultes, B.; Born, J.; Oster, H.; Hallschmid, M. Diurnal rhythm of circulating nicotinamide phosphoribosyltransferase (Nampt/visfatin/PBEF): Impact of sleep loss and relation to glucose metabolism. J. Clin. Endocrinol. Metab. 2012, 97, E218–E222. [Google Scholar] [CrossRef] [Green Version]
- Cooper, J.N.; Fried, L.; Tepper, P.; Barinas-Mitchell, E.; Conroy, M.B.; Evans, R.W.; Mori Brooks, M.; Woodard, G.A.; Sutton-Tyrrell, K. Changes in serum aldosterone are associated with changes in obesity-related factors in normotensive overweight and obese young adults. Hypertens. Res. 2013, 36, 895–901. [Google Scholar] [CrossRef] [Green Version]
- Dos Santos Moraes, A.; Pisani, L.P.; Corgosinho, F.C.; Testa Carvalho, L.O.; Masquio, D.C.L.; Jamar, G.; Sanches, R.B.; Oyama, L.M.; Dâmaso, A.R.; Belote, C.; et al. The role of leptinemia state as a mediator of inflammation in obese adults. Horm. Metab. Res. 2013, 45, 605–610. [Google Scholar] [CrossRef]
- Müller, M.J.; Enderle, J.; Pourhassan, M.; Braun, W.; Eggeling, B.; Lagerpusch, M.; Glüer, C.C.; Kehayias, J.J.; Kiosz, D.; Bosy-Westphal, A. Metabolic adaptation to caloric restriction and subsequent refeeding: The Minnesota Starvation Experiment revisited. Am. J. Clin. Nutr. 2015, 102, 807–819. [Google Scholar] [CrossRef] [PubMed]
- Williams, R.L.; Wood, L.G.; Collins, C.E.; Morgan, P.J.; Callister, R. Energy homeostasis and appetite regulating hormones as predictors of weight loss in men and women. Appetite 2016, 101, 1–7. [Google Scholar] [CrossRef]
- Thong, F.S.L.; Hudson, R.; Ross, R.; Janssen, I.A.N.; Graham, T.E. Plasma leptin in moderately obese men: Independent effects of weight loss and aerobic exercise. Am. J. Physiol. Endocrinol. Metab. 2000, 279, 307–313. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chin-Chance, C.; Polonsky, K.S.; Schoeller, D.A. Twenty-Four-Hour Leptin Levels Respond to Cumulative Short-Term Energy Imbalance and Predict Subsequent Intake1. J. Clin. Endocrinol. Metab. 2000, 85, 2685–2691. [Google Scholar] [CrossRef] [PubMed]
- Herrmann, T.S.; Bean, M.L.; Black, T.M.; Wang, P.; Coleman, R.A. High glycemic index carbohydrate diet alters the diurnal rhythm of leptin but not insulin concentrations. Exp. Biol. Med. 2001, 226, 1037–1044. [Google Scholar] [CrossRef]
- Poppitt, S.D.; Leahy, F.E.; Keogh, G.F.; Wang, Y.; Mulvey, T.B.; Stojkovic, M.; Chan, Y.K.; Choong, Y.S.; McArdle, B.H.; Cooper, G.J.S. Effect of high-fat meals and fatty acid saturation on postprandial levels of the hormones ghrelin and leptin in healthy men. Eur. J. Clin. Nutr. 2006, 60, 77–84. [Google Scholar] [CrossRef]
- Bray, G.A.; Most, M.; Rood, J.; Redmann, S.; Smith, S.R. Hormonal responses to a fast-food meal compared with nutritionally comparable meals of different composition. Ann. Nutr. Metab. 2007, 51, 163–171. [Google Scholar] [CrossRef]
- Carlson, O.; Martin, B.; Stote, K.S.; Golden, E.; Maudsley, S.; Najjar, S.S.; Ferrucci, L.; Ingram, D.K.; Longo, D.L.; Rumpler, W.V.; et al. Impact of reduced meal frequency without caloric restriction on glucose regulation in healthy, normal-weight middle-aged men and women. Metab. Clin. Exp. 2007, 56, 1729–1734. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abete, I.; Parra, D.; Crujeiras, A.B.; Goyenechea, E.; Martínez, J.A. Specific insulin sensitivity and leptin responses to a nutritional treatment of obesity via a combination of energy restriction and fatty fish intake. J. Hum. Nutr. Diet. 2008, 21, 591–600. [Google Scholar] [CrossRef]
- Nedeltcheva, A.V.; Kilkus, J.M.; Imperial, J.; Kasza, K.; Schoeller, D.A.; Penev, P.D. Sleep curtailment is accompanied by increased intake of calories from snacks. Am. J. Clin. Nutr. 2009, 89, 126–133. [Google Scholar] [CrossRef]
- Bortolotti, M.; Kreis, R.; Debard, C.; Cariou, B.; Faeh, D.; Chetiveaux, M.; Ith, M.; Vermathen, P.; Stefanoni, N.; Lê, K.A.; et al. High protein intake reduces intrahepatocellular lipid deposition in humans. Am. J. Clin. Nutr. 2009, 90, 1002–1010. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ratliff, J.; Mutungi, G.; Puglisi, M.J.; Volek, J.S.; Fernandez, M.L. Carbohydrate restriction (with or without additional dietary cholesterol provided by eggs) reduces insulin resistance and plasma leptin without modifying appetite hormones in adult men. Nutr. Res. 2009, 29, 262–268. [Google Scholar] [CrossRef] [PubMed]
- Cooper, J.A.; Polonsky, K.S.; Schoeller, D.A. Serum leptin levels in obese males during over-and underfeeding. Obesity 2009, 17, 2149–2154. [Google Scholar] [CrossRef]
- Adachi, T.; Sert-Kuniyoshi, F.H.; Calvin, A.D.; Singh, P.; Romero-Corral, A.; Van Der Walt, C.; Davison, D.E.; Bukartyk, J.; Konecny, T.; Pusalavidyasagar, S.; et al. Effect of weight gain on cardiac autonomic control during wakefulness and sleep. Hypertension 2011, 57, 723–730. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lecoultre, V.; Ravussin, E.; Redman, L.M. The fall in leptin concentration is a major determinant of the metabolic adaptation induced by caloric restriction independently of the changes in leptin circadian rhythms. J. Clin. Endocrinol. Metab. 2011, 96, E1512–E1516. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Varady, K.A.; Bhutani, S.; Klempel, M.C.; Phillips, S.A. Improvements in vascular health by a low-fat diet, but not a high-fat diet, are mediated by changes in adipocyte biology. Nutr. J. 2011, 10, 8. [Google Scholar] [CrossRef] [Green Version]
- Ebbeling, C.B.; Swain, J.F.; Feldman, H.A.; Wong, W.W.; Hachey, D.L.; Garcia-Lago, E.; Ludwig, D.S. Effects of dietary composition on energy expenditure during weight-loss maintenance. JAMA J. Am. Med. Assoc. 2012, 307, 2627–2634. [Google Scholar] [CrossRef] [Green Version]
- Chang, K.T.; Lampe, J.W.; Schwarz, Y.; Breymeyer, K.L.; Noar, K.A.; Song, X.; Neuhouser, M.L. Low glycemic load experimental diet more satiating than high glycemic load diet. Nutr. Cancer 2012, 64, 666–673. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Varady, K.A.; Bhutani, S.; Klempel, M.C.; Kroeger, C.M.; Trepanowski, J.F.; Haus, J.M.; Hoddy, K.K.; Calvo, Y. Alternate day fasting for weight loss in normal weight and overweight subjects: A randomized controlled trial. Nutr. J. 2013, 12, 146. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Calvin, A.D.; Carter, R.E.; Adachi, T.; MacEdo, P.G.; Albuquerque, F.N.; Van Der Walt, C.; Bukartyk, J.; Davison, D.E.; Levine, J.A.; Somers, V.K. Effects of experimental sleep restriction on caloric intake and activity energy expenditure. Chest 2013, 144, 79–86. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mendham, A.E.; Duffield, R.; Marino, F.; Coutts, A.J. Small-sided games training reduces CRP, IL-6 and leptin in sedentary, middle-aged men. Eur. J. Appl. Physiol. 2014, 114, 2289–2297. [Google Scholar] [CrossRef]
- Singh, P.; Sharma, P.; Sahakyan, K.R.; Davison, D.E.; Sert-Kuniyoshi, F.H.; Romero-Corral, A.; Swain, J.M.; Jensen, M.D.; Lopez-Jimenez, F.; Kara, T.; et al. Differential effects of leptin on adiponectin expression with weight gain versus obesity. Int. J. Obes. 2016, 40, 266–274. [Google Scholar] [CrossRef] [Green Version]
- Perrigue, M.M.; Drewnowski, A.; Wang, C.Y.; Song, X.; Kratz, M.; Neuhouser, M.L. Randomized Trial Testing the Effects of Eating Frequency on Two Hormonal Biomarkers of Metabolism and Energy Balance. Nutr. Cancer 2017, 69, 56–63. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, G.; Liang, L.; Bray, G.A.; Qi, L.; Hu, F.B.; Rood, J.; Sacks, F.M.; Sun, Q. Thyroid hormones and changes in body weight and metabolic parameters in response to weight loss diets: The POUNDS LOST trial. Int. J. Obes. 2017, 41, 878–886. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kessler, K.; Hornemann, S.; Petzke, K.J.; Kemper, M.; Markova, M.; Rudovich, N.; Grune, T.; Kramer, A.; Pfeiffer, A.F.H.; Pivovarova-Ramich, O. Diurnal distribution of carbohydrates and fat affects substrate oxidation and adipokine secretion in humans. Am. J. Clin. Nutr. 2018, 108, 1209–1219. [Google Scholar] [CrossRef] [PubMed]
- Trepanowski, J.F.; Kroeger, C.M.; Barnosky, A.; Klempel, M.; Bhutani, S.; Hoddy, K.K.; Rood, J.; Ravussin, E.; Varady, K.A. Effects of alternate-day fasting or daily calorie restriction on body composition, fat distribution, and circulating adipokines: Secondary analysis of a randomized controlled trial. Clin. Nutr. 2018, 37, 1871–1878. [Google Scholar] [CrossRef] [PubMed]
- Hołowko, J.; Michalczyk, M.M.; Zajac, A.; Czerwińska-Rogowska, M.; Ryterska, K.; Banaszczak, M.; Jakubczyk, K.; Stachowska, E. Six weeks of calorie restriction improves body composition and lipid profile in obese and overweight former athletes. Nutrients 2019, 11, 1461. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Benbaibeche, H.; Bounihi, A.; Koceir, E.A. Leptin level as a biomarker of uncontrolled eating in obesity and overweight. Ir. J. Med. Sci. 2020, 190, 155–161. [Google Scholar] [CrossRef]
- Poetsch, M.S.; Strano, A.; Guan, K. Role of Leptin in Cardiovascular Diseases. Front. Endocrinol. 2020, 11, 354. [Google Scholar] [CrossRef] [PubMed]
- Reyes-Barrera, J.; Sainz-Escárrega, V.H.; Medina-Urritia, A.X.; Jorge-Galarza, E.; Osorio-Alonso, H.; Torres-Tamayo, M.; Leal-Escobar, G.; Posadas-Romero, C.; Torre-Villalvazo, I.; Juárez-Rojas, J.G. Dysfunctional adiposity index as a marker of adipose tissue morpho-functional abnormalities and metabolic disorders in apparently healthy subjects. Adipocyte 2021, 10, 142–152. [Google Scholar] [CrossRef] [PubMed]
- Trinh, T.; Broxmeyer, H.E. Role for Leptin and Leptin Receptors in Stem Cells During Health and Diseases. Stem Cell Rev. Rep. 2021, 17, 511–522. [Google Scholar] [CrossRef] [PubMed]
- Thackray, A.E.; Willis, S.A.; Clayton, D.J.; Broom, D.R.; Finlayson, G.; Goltz, F.R.; Sargeant, J.A.; Woods, R.M.; Stensel, D.J.; King, J.A. Influence of short-term hyperenergetic, high-fat feeding on appetite, appetite-related hormones, and food reward in healthy men. Nutrients 2020, 12, 2635. [Google Scholar] [CrossRef] [PubMed]
- Fantuzzi, G. Adiponectin in inflammatory and immune-mediated diseases. Cytokine 2013, 64, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ouchi, N.; Walsh, K. Adiponectin as an anti-inflammatory factor. Clin. Chim. Acta 2007, 380, 24–30. [Google Scholar] [CrossRef] [Green Version]
- Arch, J.R.S. The discovery of drugs for obesity, the metabolic effects of leptin and variable receptor pharmacology: Perspectives from β3- adrenoceptor agonists. Naunyn Schmiedebergs Arch. Pharmacol. 2008, 378, 225–240. [Google Scholar] [CrossRef] [PubMed]
- Zanella, M.T.; Filho, F.F.R. Novos fármacos para o tratamento da obesidade. Arq. Bras. Endocrinol. Metabol. 2009, 53, 271–280. [Google Scholar] [CrossRef] [Green Version]
- Cassioli, E.; Rossi, E.; Squecco, R.; Baccari, M.C.; Maggi, M.; Vignozzi, L.; Comeglio, P.; Gironi, V.; Lelli, L.; Rotella, F.; et al. Reward and psychopathological correlates of eating disorders: The explanatory role of leptin. Psychiatry Res. 2020, 290, 113071. [Google Scholar] [CrossRef] [PubMed]
- Platzer, M.; Fellendorf, F.T.; Bengesser, S.A.; Birner, A.; Dalkner, N.; Hamm, C.; Lenger, M.; Maget, A.; Pilz, R.; Queissner, R.; et al. The relationship between food craving, appetite-related hormones and clinical parameters in bipolar disorder. Nutrients 2020, 13, 76. [Google Scholar] [CrossRef]
- Steiner, J.; Bernstein, H.G.; Guest, P.C.; Summergrad, P.; Oxenkrug, G. Plasma leptin correlates with anthranilic acid in schizophrenia but not in major depressive disorder. Eur. Neuropsychopharmacol. 2020, 41, 167–168. [Google Scholar] [CrossRef] [PubMed]
- Tural, R.; Altan, N.; Irkec, C.; Sahin, D.; Batur Caglayan, H.Z. The probable role of insulin resistance and SIRT1 proteins in the Alzheimer’s disease. Bratisl. Lek. Listy 2020, 121, 812–816. [Google Scholar] [CrossRef] [PubMed]
- Guzmán, A.; Hernández-Coronado, C.G.; Rosales-Torres, A.M.; Hernández-Medrano, J.H. Leptin regulates neuropeptides associated with food intake and GnRH secretion. Ann. Endocrinol. 2019, 80, 38–46. [Google Scholar] [CrossRef]
- Marcos, P.; Coveñas, R. Neuropeptidergic control of feeding: Focus on the galanin family of peptides. Int. J. Mol. Sci. 2021, 22, 2544. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Non-Randomized Controlled Trials | Aim | Methods | Main Outcomes | Age Range | Fem (%) | BMI > 25 | BMI < 25 |
|---|---|---|---|---|---|---|---|
| Sleep apnea and daytime sleepiness and fatigue: relation to visceral obesity, insulin resistance, and hypercyt [39]. | To test whether sleep apnea contributes to changes of tumor necrosis factor-α, interleukin-6, and leptin levels. | Obese, sleep apneic, and control subjects were investigated for leptin levels and other biochemical factors, and were monitored in the sleep laboratory for 4 consecutive nights. | Sleep-apneic men had higher plasma concentrations of leptin than non-apneic obese men or lean men, who had the lowest values. | 38–49 | 0 | 11 | 12 |
| No acute response of leptin to an oral fat load in obese patients and during circadian rhythm in healthy controls [40]. | To elucidate the relationship between postprandial leptin and obesity, and the possible influence of the circadian rhythm on the dynamic leptin response | Leptin and insulin responses to an oral fat load test were measured according to the time of fat load ingestion: 0700 h (diurnal (D) test) or 2200 h (nocturnal (N) test) in 9 normal-weight healthy men. | Leptin concentrations were higher in non-obese women than men, and in obese subjects than in controls. No correlation was found between postprandial leptin and insulin. Leptin concentrations were not acutely influenced by a high fat intake load. | 19–25 | 75 | 16 | 0 |
| Insulin resistance in obesity: body weight or energy balance? [41]. | To investigate the impact of diet on amelioration of the metabolic abnormalities associated with insulin resistance in obese subjects. | Twelve healthy obese subjects signed for a 6-week hypo-energetic diet, then a 4-week iso-energetic diet, then another 6-week hypo-energetic diet again. Insulin sensitivity, GLU tolerance and leptin levels were assessed. | One-third of the weight losses were achieved within the first week of diets. During the iso-energetic weight-maintaining period, insulin resistance decreased by 43%. Serum levels of leptin and triglyceride followed a similar pattern, but to a lesser extent. | 38–57 | 50 | 12 | 0 |
| Dieting is more effective in reducing weight but exercise is more effective in reducing fat during the early phase of a weight-reducing program in healthy humans [42]. | To compare the effectiveness of food restriction and physical activity on body weight, composition and concentration of serum leptin in female subjects. | Thirteen non-obese subjects signed up for a two-phase crossover study including 9-day hypo-energetic treatments, either by food restriction or increased physical activity, with a 14-day washout in between and a 5-day follow-up energy repletion period. Parameters were established before and after the two phases. | Blood leptin levels were reduced by 27.02% and 32.27% after 9 days of hypo- energetic treatment—diet and exercise, respectively. The follow-up energy repletion increased the serum leptin concentration by ~45% in both cases. | 22–55 | 100 | 0 | 13 |
| Differential effects of high-fat and high-carbohydrate isoenergetic meals on cardiac autonomic nervous system activity in lean and obese women [43]. | To compare the effects of two different isoenergetic meals on the sympathetic nervous system activity in lean and obese women. | Obese and nonobese subjects were examined after a CHO-rich and a fat-rich test meal. Blood pressure, heart rate, resting energy expenditure, plasma GLU, lipids, insulin, leptin, and norepinephrine were performed at baseline and every 1 h for 3 h after the meals. | Meal-induced thermogenesis was higher after CHO-rich, compared to fat-rich meals. CHO-rich meals caused greater cardiac SNS activation in non-obese than in obese women but SNS activation did not appear to influence the thermic effect of the food in either non-obese or obese women. | 26–45 | 100 | 15 | 15 |
| The potential association between endogenous leptin and sympathovagal activities in young obese Japanese women [44]. | To investigate the association between leptin levels and sympathovagal activities in age- and height-matched obese and non-obese healthy young women. | Plasma leptin concentrations were measured by radioimmunoassay and autonomic nervous system activity was assessed during the rest period by a power spectral analysis of heart rate variability, and were identified as very low (VLO), low (LO), and high (HI). | The global SNS index to plasma leptin concentration was markedly reduced in the obese compared to the control group. Dynamics of BMI interaction suggested a reduced sympathetic responsiveness to endogenous leptin production in obese women, and that 30% of total body fat might be the critical point at which leptin resistance is induced. | 18–22 | 100 | 15 | 15 |
| Interactions between leptin, neuropeptide-Y and insulin with chronic diurnal fasting during Ramadan [45]. | To describe changes and interactions between leptin, neuropeptide-Y, and insulin during the pattern of fasting in Ramadan. | Blood variables were measured at 1:00–2:00 p.m. during baseline and on days 14 and 28 of Ramadan fasting—abstinence from all food or drink, including water and chewing gum, from dawn to sunset. | Long-term fasting with interrupted nocturnal eating is associated with significant elevations in serum leptin and insulin and reduction in serum neuropeptide-Y. Changes in leptin appeared to be mediated by insulin. Neuropeptide-Y changes were independent of leptin or insulin. | 20–24 | 100 | 46 | |
| Extra-adipocyte leptin release in human obesity and its relation to sympathoadrenal function [46]. | To measure total and regional norepinephrine spillover, epinephrine secretion rate, and extra-adipocyte leptin release in lean and obese subjects. | Total and regional extra-adipocyte leptin release was measured in obese and nonobese men by arterial and central venous catheterization. Because the plasma clearance of leptin is primarily by renal removal, whole-body leptin release to plasma from renal plasma leptin extraction was estimated. | Whole-body leptin release was 1950 SE 643 ng/min in obese men and 382 SE 124 ng/min in non-obese men. A large contribution of brain leptin release to the plasma leptin pool was found (>40% whole body leptin release), with greater leptin release in obese than in non-obese men. | 35–50 | 0 | 20 | 22 |
| Adiponectin’s relationship with lipid metabolism is independent of body fat mass: Evidence from both cross-sectional and intervention studies [47]. | To investigate the relation between plasma adiponectin and leptin levels, insulin sensitivity, and serum lipids in a cross-sectional study. | Plasma adiponectin and leptin levels, insulin sensitivity, and serum lipids were analyzed in 242 non-obese and obese subjects participating in a 6–12-month weight-loss program. | Plasma adiponectin, but not leptin levels, were correlated with insulin sensitivity, insulin sensitivity index, HDL cholesterol, and triglycerides. Plasma adiponectin increased and plasma leptin decreased after weight loss, and the changes were correlated with subjects’ BMI. | 17–70 | 64 | 95 | 107 |
| A high-protein diet induced sustained reductions in appetite, ad libitum caloric intake, and body weight despite compensatory changes in diurnal plasma leptin and ghrelin concentrations [48]. | To test whether increasing the protein content while maintaining the carbohydrate content in the diet lowers body weight by decreasing appetite and spontaneous caloric intake. | Caloric intake, appetite, anthropometry, and leptin levels were measured in obese subjects undergoing a higher-fat, a lower-fat, and an ad libitum diet over 12 weeks. The AUC of plasma insulin vs. time, leptin, and ghrelin were measured. | Satiety was markedly increased with the isocaloric high-protein diet, despite an unchanged leptin AUC, and spontaneous energy intake decreased with the ad libitum high-protein diet, despite a decreased leptin AUC and increased ghrelin AUC. An increase in dietary protein intake from 15% to 30% of energy at a constant carbohydrate level produces a decrease in ad libitum caloric intake that appears to be mediated by increased leptin sensitivity in the CNS, resulting in weight loss. | 27–62 | 63 | 19 | 0 |
| Determinants of coronary flow velocity reserve in healthy young men [49]. | To identify the risk markers for attenuated coronary flow velocity reserve that exist in healthy young men without evident atherosclerotic risk factors. | Echocardiography and ergospirometry sessions were conducted, and coronary blood flow was measured in non-obese men. Anthropometric measures and the regulation of fat metabolism were assessed by determining the adiponectin and leptin levels. | There was no relationship between coronary flow velocity reserve and serum lipids or body mass index. It was concluded that abdominal fat accumulation and low aerobic fitness are independently associated with coronary flow velocity reserve in men. | 19–36 | 0 | 0 | 37 |
| PPARγ activity in subcutaneous abdominal fat tissue and fat mass gain during short-term overfeeding [50]. | To investigate whether initial subcutaneous PPARγ activity is related to fat mass generation during overfeeding. | Fourteen healthy non-obese subjects were overfed with a diet supplying 50% more energy than the baseline energy requirements for 14 days. Blood analyses of leptin, insulin and GLU, as well as mRNA expression of PPARg1, PPARg2, aP2 and UCP2, in fasting subcutaneous abdominal fat were performed. | Basal levels of Initial PPARg1 and 2, aP2 and UCP2 mRNAs were not related to fat gain. Changes in PPARg1, PPARg2 and aP2 mRNAs were positively related to changes in plasma leptin. | 20–28 | 100 | 0 | 14 |
| The consequences of omitting or adding a meal in males, regarding body composition, food intake, and metabolism [51]. | To investigate the consequence on body composition and related biological and metabolic parameters of omitting or adding a meal. | Twenty-four nonobese subjects were recruited to either omit or add a fourth meal during a 28-day period; blood parameters were tested from lunch to the spontaneously requested dinner in each condition. | Omitting a meal was followed by increases in fat mass and late-evening leptin concentration. Adding a meal had no effect. In both groups, the change in energy content of the fourth eating regime was correlated with the change in adiposity. | 19–25 | 23 | ||
| Ramadan fasting’s effect on plasma leptin, adiponectin concentrations, and body composition in trained young men [52]. | To evaluate the effect of Ramadan fasting on parameters of insulin resistance in trained athletes at rest and after aerobic exercise. | Nonobese subjects performed a progressive cycle-ergometer test one week before, after the first week, and during the fourth week of Ramadan, with biochemical analyses of GLU, cholesterol, HDL cholesterol, triglycerides, creatinine, and serum proteins, as well as insulin, leptin, and adiponectin. | Ramadan fasting was associated with a reduction in body mass and body fat without changes to leptin or adiponectin levels. | 18–22 | 0 | 9 | |
| Improvements in coronary heart disease risk indicators by alternate-day fasting involving adipose tissue modulations [53]. | To examine the effects of alternate-day fasting on adipokine profile, body composition, and coronary heart disease risk indicators in obese adults. | Obese subjects signed for a 10-week trial with the following consecutive diet phases: 2-week baseline control, 4-week alternate-day fasting with controlled feeding, and 4-week alternate-day fasting with self-selected feeding. | Leptin and resistin concentrations were reduced by 21 and 23%, respectively, after 8 weeks of treatment. Lower triacylglycerol concentrations were associated with augmented adiponectin and reduced leptin concentrations post-treatment. | 40–52 | 75 | 12 | 0 |
| Regulation of energy balance during long-term physical inactivity, induced by bed rest, with and without exercise training [54]. | To determine the long-term effects of physical inactivity on energy balance regulation and to test the effect of exercise training on energy balance adjustment to physical inactivity. | Nonobese subjects were divided into two groups: 1—accomplished a strict 60-day bed rest, 2—a combined aerobic/resistance exercise training concomitantly with bed rest. Body composition, spontaneous energy intake, hunger, total energy expenditure, and fasting gut hormones were measured. | Leptin levels did not change in both groups and no group effect was observed. Leptin levels were strongly associated with fat mass. Fasting leptin levels, adjusted for fat mass, negatively correlated with the spontaneous energy intake after the program. | 30–40 | 100 | 0 | 16 |
| Relationships between serum adiponectin, leptin concentrations and bone mineral density, and bone biochemical markers in Chinese women [55]. | To investigate whether serum adipocytokines concentrations are associated with bone mineral density and bone turnover markers. | Serum adiponectin, leptin concentrations, bone turnover biochemical markers, and bone mineral density were analyzed in 265 non-obese premenopausal women. | Adiponectin and leptin were the main circulating peptides secreted by the adipose tissue. There were no significant correlations between leptin concentrations and bone-specific alkaline phosphatase in premenopausal and postmenopausal women. | 26–44 | 100 | 0 | 265 |
| Monitoring the diurnal rhythm of circulating nicotinamide phosphoribosyltransferase (Nampt/visfatin/PBEF): impact of sleep loss and its relationship with glucose metabolism [56]. | To examine the 24-h profile of the serum Nampt in humans under conditions of sleep and sleep deprivation, and to relate the Nampt pattern to morning postprandial GLU metabolism. | Non-obese subjects participated in two 24-h lab sessions starting at 1800 h, including either a regular 8 h per night sleep or continuous wakefulness. Serum Nampt and leptin were measured at 1.5- to 3-h intervals. In the morning, plasma GLU and serum insulin responses to standardized breakfast intake were determined. | Under regular sleep-wake conditions, Nampt levels displayed a pronounced diurnal rhythm, peaking during the early afternoon, which was inverse to leptin profiles, which peaked in the early night. When subjects stayed awake, the Nampt rhythm was preserved but the phase advanced by about 2 h. | 20–24 | 0 | 14 | |
| Changes in serum aldosterone, associated with changes in obesity-related factors in normotensive overweight and obese young adults [57]. | To verify whether reductions in serum aldosterone are associated with favorable changes in obesity-related factors in normotensive overweight/obese young adults. | Obese subjects in a clinical trial, examining the effects of a 1-year diet and physical activity intervention with or without sodium restriction on vascular health. Bodyweight, serum aldosterone, 24-h sodium and potassium excretion and obesity-related factors were measured at baseline, 6, 12 and 24 months. | Decreases in aldosterone were found to be associated with decreases in leptin, insulin, homeostasis assessment of insulin resistance, heart rate, tonic cardiac sympathovagal balance, and increases in adiponectin. | 20–45 | 80 | 285 | 0 |
| The role of leptinemia as a mediator of inflammation in obese adults [58]. | To investigate the role of leptinemia, adjusted by tertiles, on the inflammatory state in obese adults, according to obesity degree. | Obese subjects assessed for inflammation markers such as leptin, adiponectin, and the plasminogen activator inhibitor were grouped according to the adjusted tertiles of the leptin levels. | A negative correlation between leptin concentrations and the adiponectin/leptin ratio, and a positive correlation with the leptin/adiponectin ratio. The ratios were decreased and increased, respectively, according to obesity degree. | 30–58 | 72 | 43 | 0 |
| Metabolic adaptation to caloric restriction and subsequent refeeding: the Minnesota starvation experiment revisited [59]. | To address the variance and kinetics of adaptive thermogenesis, its associations with body composition in the context of endocrine determinants, and its effect on weight regain. | Non-obese subjects accomplished an over-feeding (1 week at + 50% of energy needs) followed by a caloric restriction (3 weeks at 50% of energy needs), and refeeding diet (2 weeks at +50% of energy needs). Adaptive thermogenesis and its determinants were measured, together with body composition, whole-body MRI, isotope dilution, and nitrogen and fluid balances. | Caloric restriction reduced fat mass and fat-free mass and led to reductions in energy expenditure, heart rate, blood pressure, creatinine clearance, the activity of the sympathetic nervous system, plasma leptin, insulin, adiponectin, T3, and testosterone. Sympathetic nervous system activity, plasma leptin, ghrelin, and T3 and their changes were not related to adaptive thermogenesis. | 20–37 | 0 | 32 | |
| Energy homeostasis and appetite-regulating hormones as predictors of weight loss in men and women [60]. | To characterize plasma leptin, ghrelin and adiponectin concentrations in overweight and obese males and females, and to determine whether baseline concentrations of these hormones predict weight loss. | Subjects were assessed for biochemical analyses pre- and post-3- and 6-month participation in a weight-loss program. Baseline concentrations of leptin, adiponectin and ghrelin were determined. | Females had higher baseline concentrations of leptin, adiponectin and ghrelin, as well as ratios of leptin:adiponectin and leptin:ghrelin. Additionally in females, a higher baseline total ghrelin predicted greater weight loss and a higher ratio of leptin:ghrelin predicted weight gain at six months. A higher leptin:ghrelin ratio is a predictor of weight loss failure in females. | 18–60 | 55 | 119 | 0 |
| Randomized and Quasi-Randomized Controlled Trials | Aim | Methods | Main Outcomes | Age Range | Fem (%) | BMI > 25 | BMI < 25 |
|---|---|---|---|---|---|---|---|
| Plasma leptin in moderately obese men: independent effects of weight loss and aerobic exercise [61]. | To describe the independent effects of weight loss and exercise on plasma leptin levels. | Fifty-two sedentary obese men were assigned for 12 weeks to one of the following conditions: (1) control, (2) diet-induced weight loss, (3) exercise-weight stable, and (4) exercise-induced weight loss. Diets consisted of 55–60% carbohydrate, 15–20% protein and 20–25% fat. | Plasma leptin and BMI did not change in the control or the weight-stable groups. Diet and exercise similarly decreased plasma leptin after weight loss. Exercise in the absence of weight loss did not alter leptin levels. Changes in leptin correlated with changes in total and subcutaneous, but not in visceral, adipose tissues. | 42–48 | 0 | 52 | 0 |
| Twenty-four-hour leptin levels respond to cumulative short-term energy imbalance and predict subsequent intake [62]. | To determine whether leptin serves as an indicator of short-term energy balance by measuring the acute effects of small manipulations in energy intake on leptin levels in normal individuals. | Four consecutive dietary treatments of 3 days each were applied to 6 healthy non-obese subjects, and leptin levels were serially measured throughout the study. | During washout after the underfeeding period, leptin levels were 88% of the eucaloric baseline, compared with 135% following overfeeding. Leptin levels did not return to baseline with the washout diet and were restored only after cross-over to the complementary diet treatment. Ad libitum intake changes correlated with changes in leptin levels. | 18–39 | 0 | 6 | 0 |
| A high-glycemic-index carbohydrate diet alters the diurnal rhythm of leptin but not insulin concentrations [63]. | To determine whether post-prandial and diurnal patterns of leptin levels were altered by the type of diet amount of fat. | Nine healthy non-obese adults signed for an 8-day regimen in one of the following isocaloric diets containing 15% protein: A—high glycemic index (GI) CHO, 30% fat; B—low GI CHO, 30%fat; C—high GI CHO, 20% fat; and D—low GI CHO, 20% fat. Serum GLU, insulin, and leptin levels were measured for 24 h on day 8, and on day 9 during an OGTT. | Diets with HGI altered the serum leptin diurnal pattern, causing a rise in leptin beginning at 13:00 h. The area under the curve for leptin between 12:30 and 24:00 h in these diets was 17% greater. During the OGTT, leptin concentrations were similar in all diets. | 20–37 | 44 | 9 | |
| Effect of high-fat meals and fatty acid saturation on postprandial levels of the hormones ghrelin and leptin in healthy men [64]. | To investigate whether high-fat meals, which differed in saturated fatty acid content, acutely modified these hormones. | Eighteen nonobese subjects experienced a high-fat test meal (71% of energy as fat) as breakfast with either a high or low saturated:unsaturated fatty acid ratio on two occasions. Fasting and postprandial measurements of ghrelin, leptin and insulin were performed over 6 h, and at 10 and 24 h following a fat-exclusion lunch, snack, and dinner. | There was no significant effect of fatty meals (either saturated or unsaturated) on ghrelin over 6 h. Leptin decreased in response to both high-fat meals. There was no correlation between ghrelin or leptin and circulating insulin. Increasing dietary saturated fatty acids had no deleterious effects on leptin or total ghrelin. | 18–28 | 0 | 18 | |
| Hormonal responses to a fast-food meal compared with nutritionally comparable meals of different composition [65]. | To compare differences in the acute metabolic response to fast food and “healthy” meals with a similar composition. | Obese subjects were given a standard breakfast followed by one of the three lunches at noon: a fast-food meal; an organic beef meal; and a turkey sandwich made with organic foods and an organic orange juice. Blood parameters were measured every 30 min over a period of 6 h. | Leptin levels, whether expressed as a percent of baseline or absolute values, showed no significant pattern of change over the 6-h postprandial period. | 19–27 | 0 | 6 | 0 |
| Impact of reduced meal frequency, without caloric restriction on glucose regulation in healthy, normal-weight middle-aged men and women [66]. | To evaluate the influence of reduced meal frequency, without a reduction in energy intake, on GLU metabolism in normal-weight, healthy male and female subjects. | Nonobese subjects underwent two 8-week ad libitum diet periods for weight maintenance with either three meals/day (breakfast, lunch, and dinner) or one meal/day (during a 4-h period in the early evening; 4:00 to 8:00 p.m.), with an 11-week washout period. Energy metabolism was evaluated throughout the study by morning OGTT, GLU, insulin, glucagon, leptin, ghrelin, adiponectin, resistin, and BDNF levels. | Diet had no significant effects on morning plasma glucagon, leptin, adiponectin, resistin, and BDNF. | 40–50 | 33 | 0 | 15 |
| Specific insulin sensitivity and leptin responses to a nutritional treatment of obesity, via a combination of energy restriction and fatty fish intake [67]. | To investigate whether the inclusion of three fatty fish servings per week within a hypocaloric diet may have specific health effects on insulin and leptin functions. | Obese subjects were assigned to a control, a fish or a fatty fish-based diet, with a −30% total energy expenditure caloric restriction over an 8-week period. Anthropometry, body composition, lipid profile, leptin and insulin values were measured pre- and post-intervention. | A decrease in leptin was only observed after the fish-enriched diets. There was a positive correlation between insulin and leptin decreases. Sixteen percent of the variability in leptin changes could be explained by the HOMA index change and the type of diet. | 29–45 | 44 | 32 | 0 |
| Sleep curtailment, accompanied by an increased intake of calories from snacks [68]. | To test whether the curtailment of human sleep could promote excessive energy intake. | Obese subjects were confined in a sleep laboratory for 14-day stays with ad libitum access to palatable food and either 5.5-h or 8.5-h bedtimes. Calories consumed during each bedtime condition, total energy expenditure, and 24-h profiles of serum leptin and ghrelin were analyzed. | No significant differences were found in serum leptin and ghrelin between the two sleep conditions. Twenty-four-hour leptin concentrations increased in a similar fashion at the end of the 5.5- and 8.5-h bedtime interventions. | 35–45 | 45 | 11 | 0 |
| High protein intake reduced intrahepatocellular lipid deposition in humans [69]. | To assess the effect of high protein intake on high-fat diet-induced IHCL accumulation and insulin sensitivity in healthy young men. | Non-obese subjects crossover after 4 days on a hypercaloric high-fat diet; a hypercaloric high-fat, high-protein diet; and a control, isocaloric diet. Fasting metabolism analyses were performed at the end of the fourth day of each period, and the expression of key lipogenic genes was assessed in subcutaneous adipose tissue biopsies. | The high-fat diet and hypercaloric high-fat high-protein diet significantly increased plasma leptin concentrations. | 22–26 | 0 | 10 | |
| Carbohydrate restriction (with or without additional dietary cholesterol, provided by eggs) reduces insulin resistance and plasma leptin without modifying appetite hormones in adult men [70]. | To evaluate the effects of additional dietary cholesterol and protein, provided by whole eggs while following carbohydrate-restricted diets, on insulin resistance and appetite hormones. | Obese subjects were allocated to the following 12-week regimes: egg (640 mg/d additional dietary cholesterol) or placebo (0 mg/d additional dietary cholesterol) while following carbohydrate-restricted diets. | There were significant reductions in fasting insulin and fasting leptin concentrations for both groups, which were correlated with the reductions in body weight and body fat. | 40–70 | 0 | 31 | 0 |
| Serum leptin levels in obese males during over- and underfeeding [71]. | To test whether leptin responds to short-term changes in energy balance in obese and lean males. | Obese subjects were signed to four consecutive dietary treatment periods of 3 days each: eucaloric diet, followed by periods of overfeeding (130% of total energy expenditure) or underfeeding (70%), separated by eucaloric (100%) washout period. Total energy expenditure and leptin levels were measured during the third day of each treatment. | Leptin levels were acutely responsive to negative energy balance, but not to positive energy balance unless subjects were previously underfed. During overfeeding, leptin levels increased by 25 ± 11% when subjects were underfed first, but did not increase when subjects were overfed first. | 20–35 | 0 | 8 | 0 |
| Effect of weight gain on cardiac autonomic control during wakefulness and sleep [72]. | To investigate the effect of fat gain on cardiac autonomic control during wakefulness and sleep in humans. | Non-obese subjects were engaged in either a gaining-weight diet (4 kg in 8 weeks) followed by an 8-week weight-loss period or a weight-maintainer control. Analysis of heart rate variability was performed at baseline, after weight gain, and after weight loss, to examine the relationship between changes in heart rate variability and changes in insulin, leptin, and adiponectin levels. | Weight gain was associated with increased insulin and leptin concentrations. Insulin, leptin, and adiponectin increased after fat gain and fell after fat loss. | 26–32 | 38 | 36 | |
| The fall in leptin concentration is a major determinant of the metabolic adaptation induced by caloric restriction, independently of the changes in leptin circadian rhythms [73]. | To explore the changes in 24-h leptin circadian rhythms in response to caloric restriction, and the relationship between these changes and metabolic adaptation. | Obese subjects were assigned to a control group or a caloric restriction group for 6 months. Leptin concentration was assessed every 30 min for 24 h, and leptin circadian variations were fitted by Cosinor analysis. Sedentary energy expenditure and urinary catecholamine excretion were measured for 24 h in a metabolic chamber. | A decrease in the 24-h mesor of circulating leptin of 44 ± 3% was observed. The diurnal amplitude of leptin slightly increased over the diet. A metabolic adaptation of −126 ± 25 kcal/d with a decrease in urinary norepinephrine and T3 concentrations was observed, independently of the changes in mesor leptin. | 26–48 | 57 | 46 | 0 |
| Improvements in vascular health by a low-fat diet, but not a high-fat diet, are mediated by changes in adipocyte biology [74]. | To examine how modulations in flow-mediated dilation by high-fat and low-fat diets relate to changes in adipocyte parameters. | Obese subjects were randomized to a high-fat diet (60% kcal as fat) or a low-fat diet (25% kcal as fat) for 6 weeks. Both groups were restricted by 25% of their energy needs. | Bodyweight decreased in both groups. Fat mass and waist circumference were reduced in the low-fat group only (−4.4 ± 0.3 kg; −3.6 ± 0.8 cm, respectively). Increases in plasma adiponectin and decreases in resistin were shown by the low-fat diet only. Greater decreases in leptin were observed with low-fat (−48 ± 9%) vs. high-fat (−28 ± 12%) diets. | 32–40 | 70 | 17 | 0 |
| Effects of dietary composition on energy expenditure during weight-loss maintenance [75]. | To examine the effects of three diets, differing widely in macronutrient composition and glycemic load, on energy expenditure following weight loss. | Obese subjects, after 10% to 15% weight loss in a diet, consumed an isocaloric low-fat diet (60% carbohydrate, 20% fat, 20% protein; high glycemic load), low–glycemic index diet (40% carbohydrate, 40% fat, and 20% protein; moderate glycemic load), and very low-carbohydrate diet (10% carbohydrate, 60% fat, and 30% protein; low glycemic load) in random order, for 4 weeks. | Leptin was highest with the low-fat diet, intermediate with the low-glycemic index diet, and lowest with the very-low-carbohydrate diet. | 24–38 | 38 | 21 | 0 |
| Whether a low-glycemic-load experimental diet was more satiating than a high-glycemic-load diet [76]. | To investigate the effect of low and high glycemic load diets on satiety and BMI, sex, and serum leptin. | Non-obese and obese subjects were crossed in a feeding study, testing low- vs. high-glycemic-load diets for 28 days, with isocaloric macronutrient distributions differing only in glycemic load and fiber. Fasting leptin levels were measured after each of the diet periods. | Serum leptin concentrations did not differ after the two diet treatments. | 19–42 | 50 | 42 | 40 |
| Alternate day fasting for weight loss in normal weight and overweight subjects: a randomized controlled trial [77]. | To examine the effect of alternate-day fasting on body weight and coronary heart disease risk in non-obese subjects. | Obese subjects were randomized to either an alternate day fasting—ADF; ad libitum “feed day”, alternated with 25% energy intake “fast day”—group or a control group for 12 weeks. | Bodyweight decreased in the alternate-day fasting group. Adiponectin levels increased while leptin decreased in the intervention group vs. controls. LDL cholesterol, HDL cholesterol, homocysteine and resistin concentrations remained unchanged. | 42–50 | 73 | 30 | 0 |
| Effects of experimental sleep restriction on caloric intake and activity energy expenditure [78]. | To determine the effect of 8 days/8 nights of sleep restriction on caloric intake, activity energy expenditure, and circulating levels of leptin and ghrelin. | Obese subjects were randomized into usual sleep vs. a 2/3 sleep restriction for 8 days/8 nights in a hospital-based clinical research unit. Parameters of caloric intake, activity energy expenditure, and circulating levels of leptin and ghrelin were analyzed. | Caloric intake increased in the sleep-restricted group and decreased in the control group. Sleep restriction was not associated with changes in activity energy expenditure. No change was seen in the levels of leptin or ghrelin. | 18–40 | 35 | 17 | 0 |
| Small-sided games training reduces CRP, IL-6 and leptin in sedentary, middle-aged men [79]. | To improve chronic systemic inflammation, which provides protection against the ensuing development of chronic disease. | Subjects were engaged in cycling or small-sided games 3 days/week for 8 weeks, or sedentary control conditions. Pre- and post- evaluations included a dual-energy X-ray absorptiometry scan, sub-maximal aerobic capacity (VO2) and fasting blood analyses of C-reactive protein, interleukin-6, IL-1β, tumor necrosis factor-α, and leptin; IL-10, IL-1 receptor agonist, and adiponectin. | Both treatments increased submaximal VO2 and decreased total body fat mass and C-reactive protein. Only small-sided games increased total body fat-free mass and the concentration of plasma IL-6 and leptin. | 40–58 | 0 | 32 | 0 |
| Differential effects of leptin on adiponectin expression with weight gain, versus obesity [80]. | To investigate the role of weight gain, and the consequent changes in leptin, on altering adiponectin expression in humans. | Non-obese subjects were engaged to either gain 5% of body weight by 8 weeks of overfeeding or to maintain weight. | Modest weight gain (3.8 ± 1.2 kg), but not weight maintenance, resulted in an increased adiponectin level, positively correlated with changes in leptin. In vitro analyses showed that leptin activates cellular signaling pathways and increases adiponectin mRNA in normal-weight, but not in obese, adipose tissue. Obese subjects showed increased caveolin-1 expression, which attenuates leptin-dependent increases in adiponectin. | 22–35 | 14 | 0 | 44 |
| Randomized trial testing the effects of eating frequency on two hormonal biomarkers of metabolism and energy balance [81]. | To verify the possible influence of eating frequency on fasting plasma insulin-like growth factor-I and leptin. | The effects of eating frequency on fasting plasma insulin-like growth factor-I (IGF-1) and leptin were observed in non-obese subjects engaged in two eucaloric interventions lasting 21 days each: low frequency (3 eating occasions/day) and high frequency (8 eating occasions/day). | There were lower serum IGF-1 levels when subjects underwent high- compared to low-eating-frequency conditions. There was no association between eating frequency and plasma leptin levels. | 19–38 | 73 | 11 | |
| Thyroid hormones and changes in body weight and metabolic parameters in response to weight-loss diets: the “Pounds Lost” trial [82]. | To examine the associations between thyroid hormones and changes in body weight and resting metabolic rate (RMR) in a diet-induced weight-loss setting. | Obese subjects from the 2-year “Pounds Lost” trial were assessed for body weight and resting metabolic parameters. Thyroid hormones (free triiodothyronine (T3), free thyroxine (T4), total T3, total T4, and thyroid-stimulating hormone (TSH)), anthropometric measurements, and biochemical parameters were assessed at baseline, 6 months, and 24 months. | Decreases in free T3 and total T3 levels were positively associated with changes in body weight and leptin, GLU, insulin, and triglycerides, both in 6 months and 2 years. No association was observed between baseline leptin levels and weight change. | 30–70 | 83 | 569 | 0 |
| Diurnal distribution of carbohydrates and fat affects substrate oxidation and adipokine secretion in humans [83]. | To investigate the effects of dietary patterns on energy metabolism, and circulating lipids, adipokines, and inflammatory markers. | Non-obese subjects underwent two isocaloric 4-week diets: (1) carbohydrate-rich meals until 13:30 and fat-rich meals between 16:30 and 22:00; or (2) the inverse sequence of meals. During a 12-h clinical investigation day after each intervention period, two meal tolerance tests were performed, at 09:00 and 15:40, respectively. Substrate oxidation and circulating lipids, adipokines, and cytokines were assessed pre- and postprandially. | Diurnal patterns of triglycerides, LDL cholesterol, leptin, visfatin, and LPS-induced cytokine secretion in blood leukocytes were modulated by the diets. Average daily concentrations of leptin and visfatin were lower on Diet 2 than on Diet 1. | 42–48 | 0 | 29 | |
| Effects of alternate-day fasting or daily calorie restriction on body composition, fat distribution, and circulating adipokines: secondary analysis of a randomized controlled trial [84]. | To compare changes in the visceral adipose tissue:saturated adipose tissue ratio, fat-free mass:total mass ratio, and the adipokine profile between alternate-day fasting and daily calorie restriction. | Obese subjects were signed to (1) alternate-day fasting (alternating every 24 h between consuming 25% or 125% of energy needs); (2) calorie restriction (consuming 75% of needs every day); or (3) control (consuming 100% of needs every day) over 24 weeks. | Visceral adipose tissue:subcutaneous adipose tissue ratio did not change. The FFM:total mass ratio similarly increased in both groups compared to the control group. Circulating leptin levels decreased by 18% and 31%, respectively, in the diet groups relative to control. The levels of adiponectin, resistin, IL-6, and TNF-a did not change in either intervention group relative to control. | 42–48 | 84 | 79 | 0 |
| Six weeks of calorie restriction improved body composition and lipid profile in obese and overweight former athletes [85]. | To compare the impact of 20% vs. 30% (of total daily energy expenditure) reduction of daily caloric intake over 6 weeks on body mass reduction and insulin metabolism in former athletes. | Obese subjects signed to 6-week of either a 20% or a 30% restriction diet had body composition variables, lipid profile (total lipids—TL; total cholesterol—TCh; HDL cholesterol—HDL; LDL cholesterol—LDL; triglycerides—TG), GLU, insulin, IGF-1, leptin and adiponectin. | A decrease in TG, TL and leptin, and an increase in adiponectin levels, were observed in both diet groups. | 29–41 | 0 | 63 | 0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
de Assis, G.G.; Murawska-Ciałowicz, E. Leptin—A Potential Bridge between Fat Metabolism and the Brain’s Vulnerability to Neuropsychiatric Disorders: A Systematic Review. J. Clin. Med. 2021, 10, 5714. https://doi.org/10.3390/jcm10235714
de Assis GG, Murawska-Ciałowicz E. Leptin—A Potential Bridge between Fat Metabolism and the Brain’s Vulnerability to Neuropsychiatric Disorders: A Systematic Review. Journal of Clinical Medicine. 2021; 10(23):5714. https://doi.org/10.3390/jcm10235714
Chicago/Turabian Stylede Assis, Gilmara Gomes, and Eugenia Murawska-Ciałowicz. 2021. "Leptin—A Potential Bridge between Fat Metabolism and the Brain’s Vulnerability to Neuropsychiatric Disorders: A Systematic Review" Journal of Clinical Medicine 10, no. 23: 5714. https://doi.org/10.3390/jcm10235714
APA Stylede Assis, G. G., & Murawska-Ciałowicz, E. (2021). Leptin—A Potential Bridge between Fat Metabolism and the Brain’s Vulnerability to Neuropsychiatric Disorders: A Systematic Review. Journal of Clinical Medicine, 10(23), 5714. https://doi.org/10.3390/jcm10235714