
The Active Isoforms of MGP Are Expressed in Healthy and Varicose Veins without Calcification

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Collection and Preparation
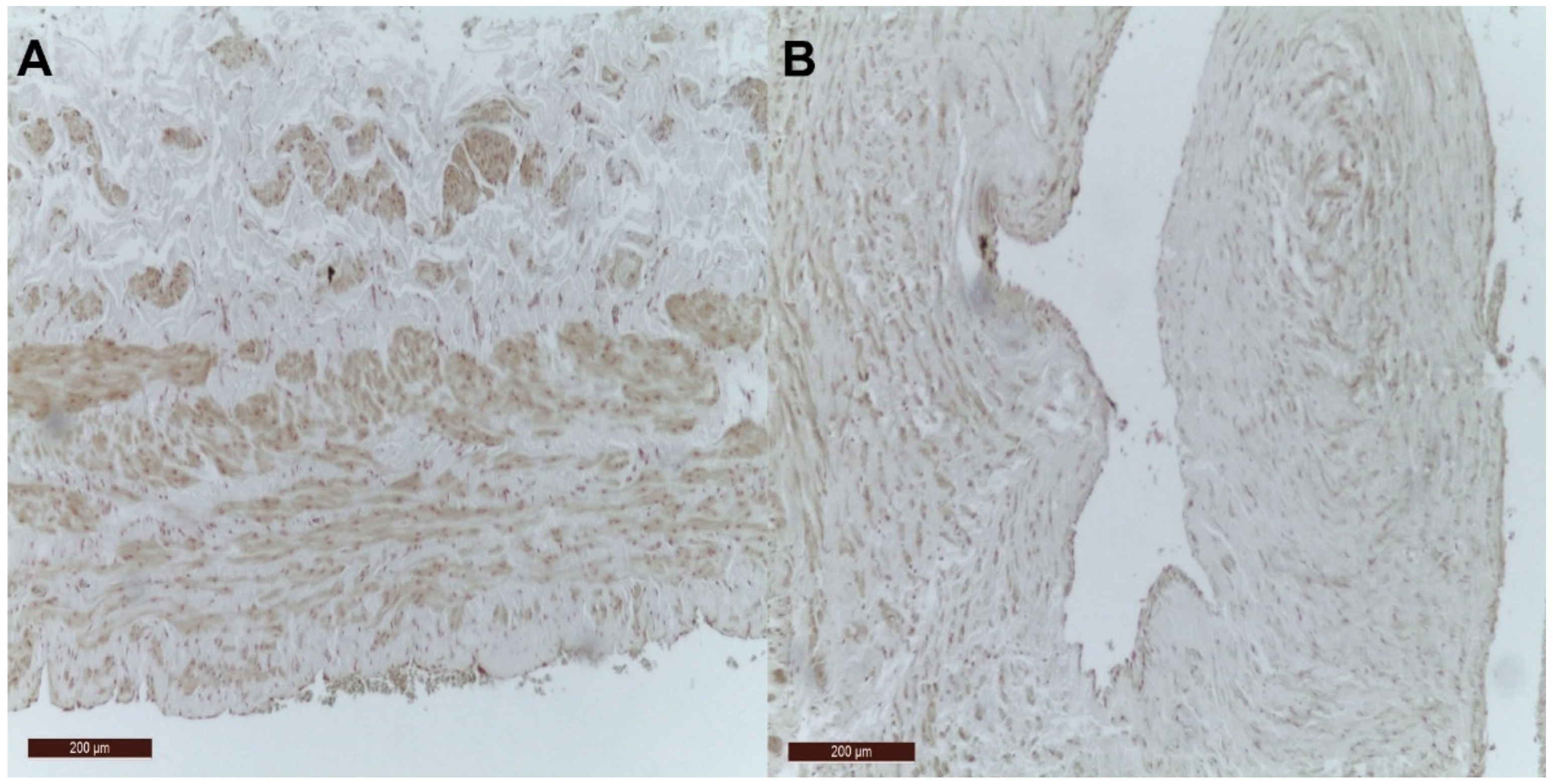
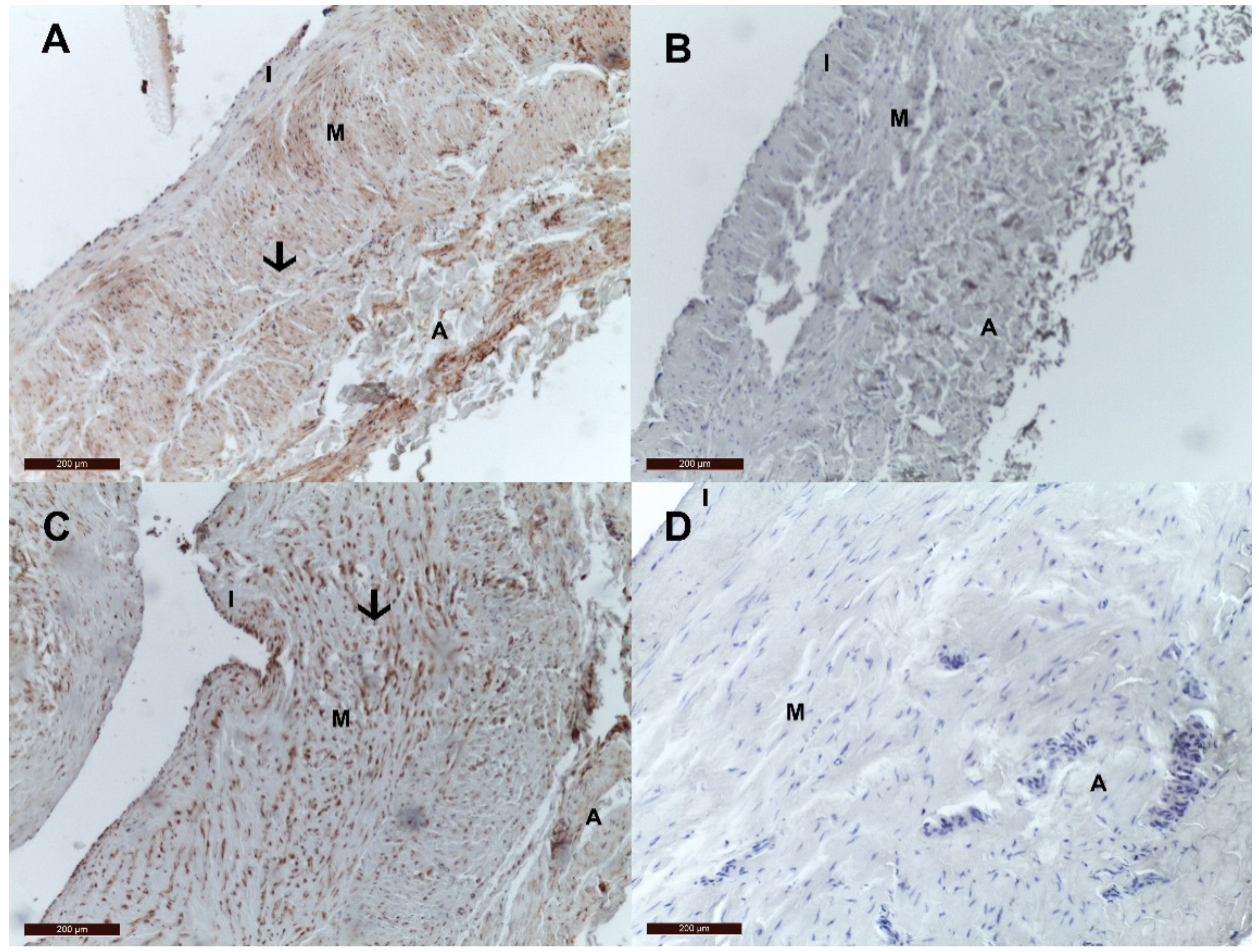
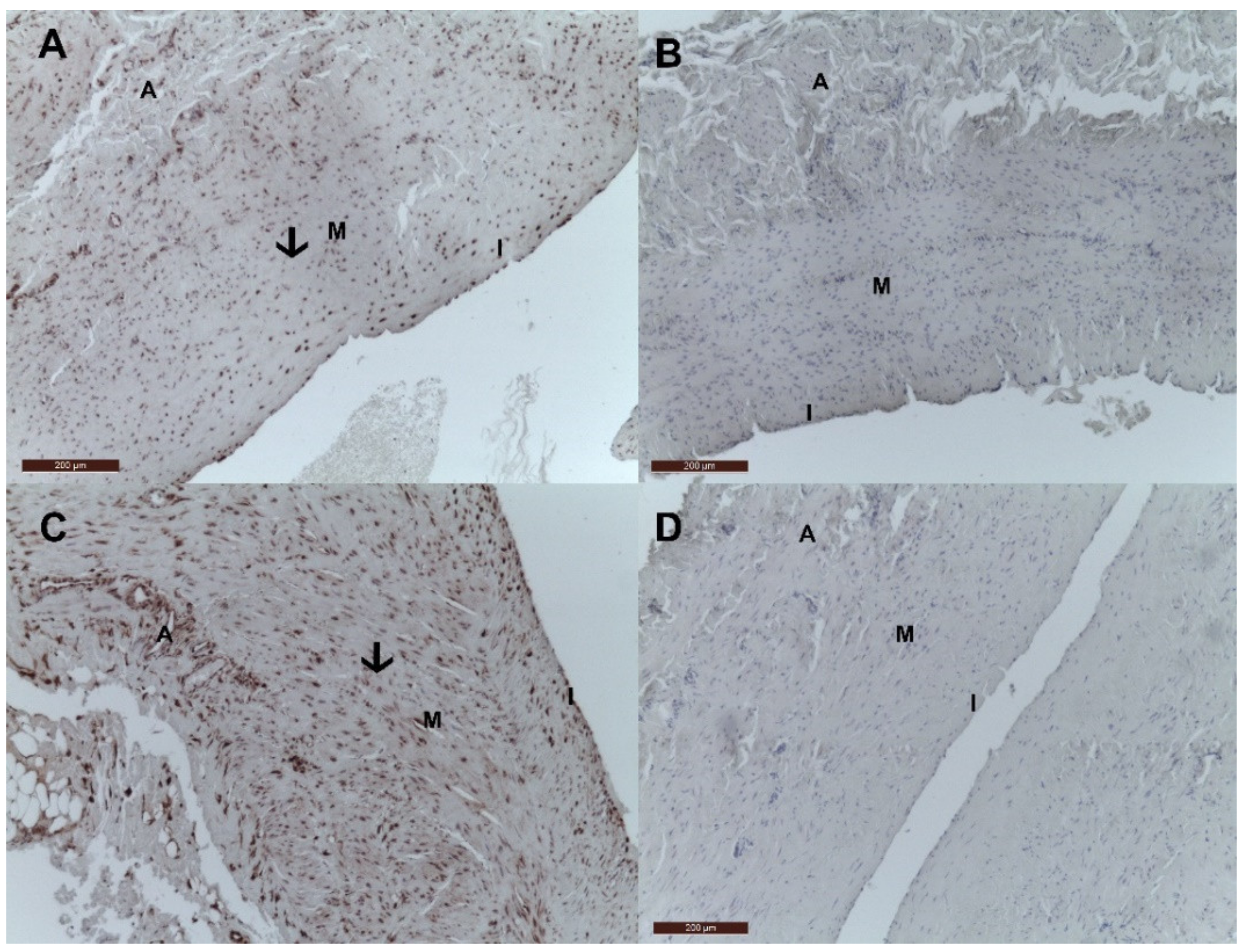
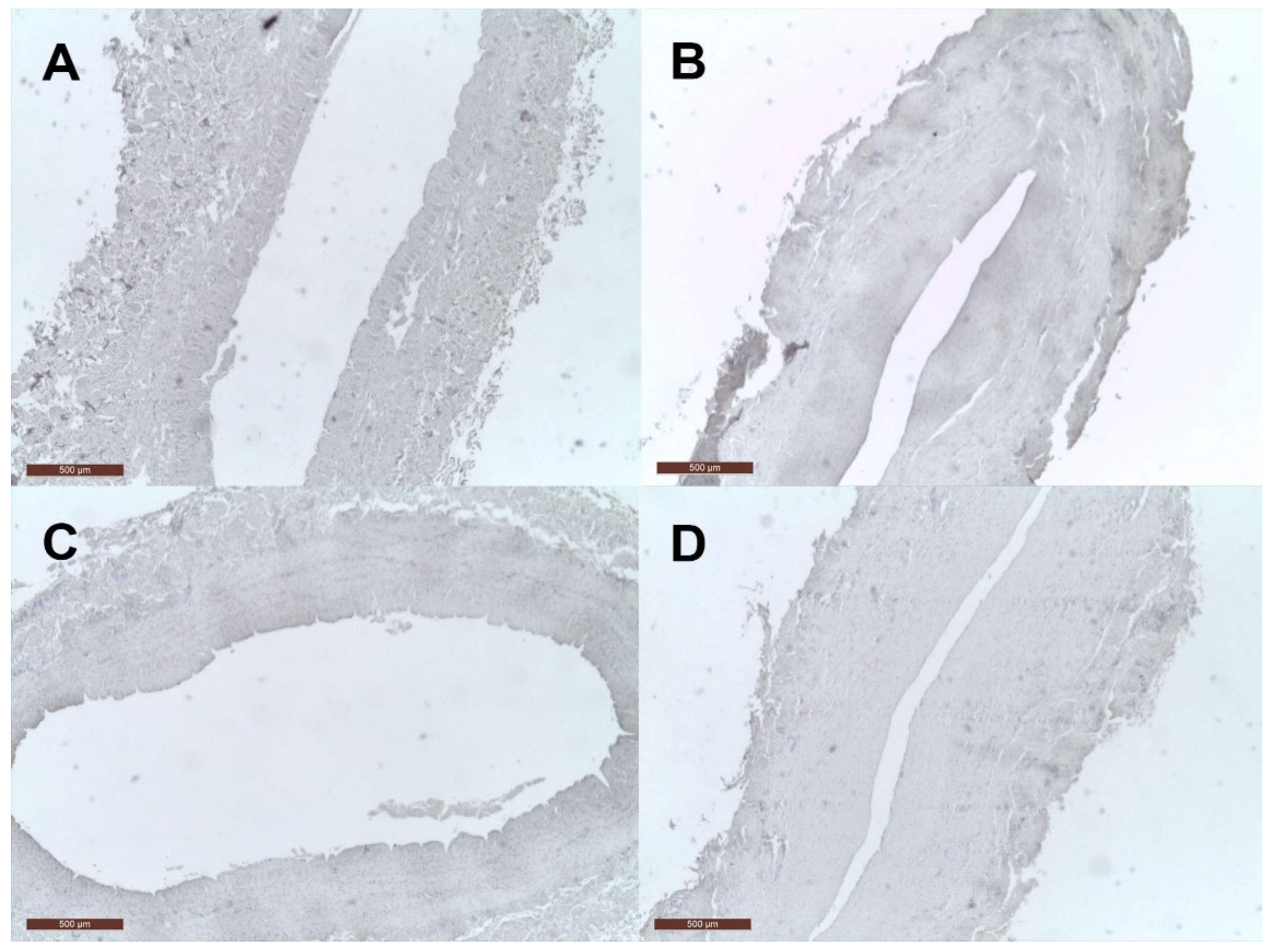
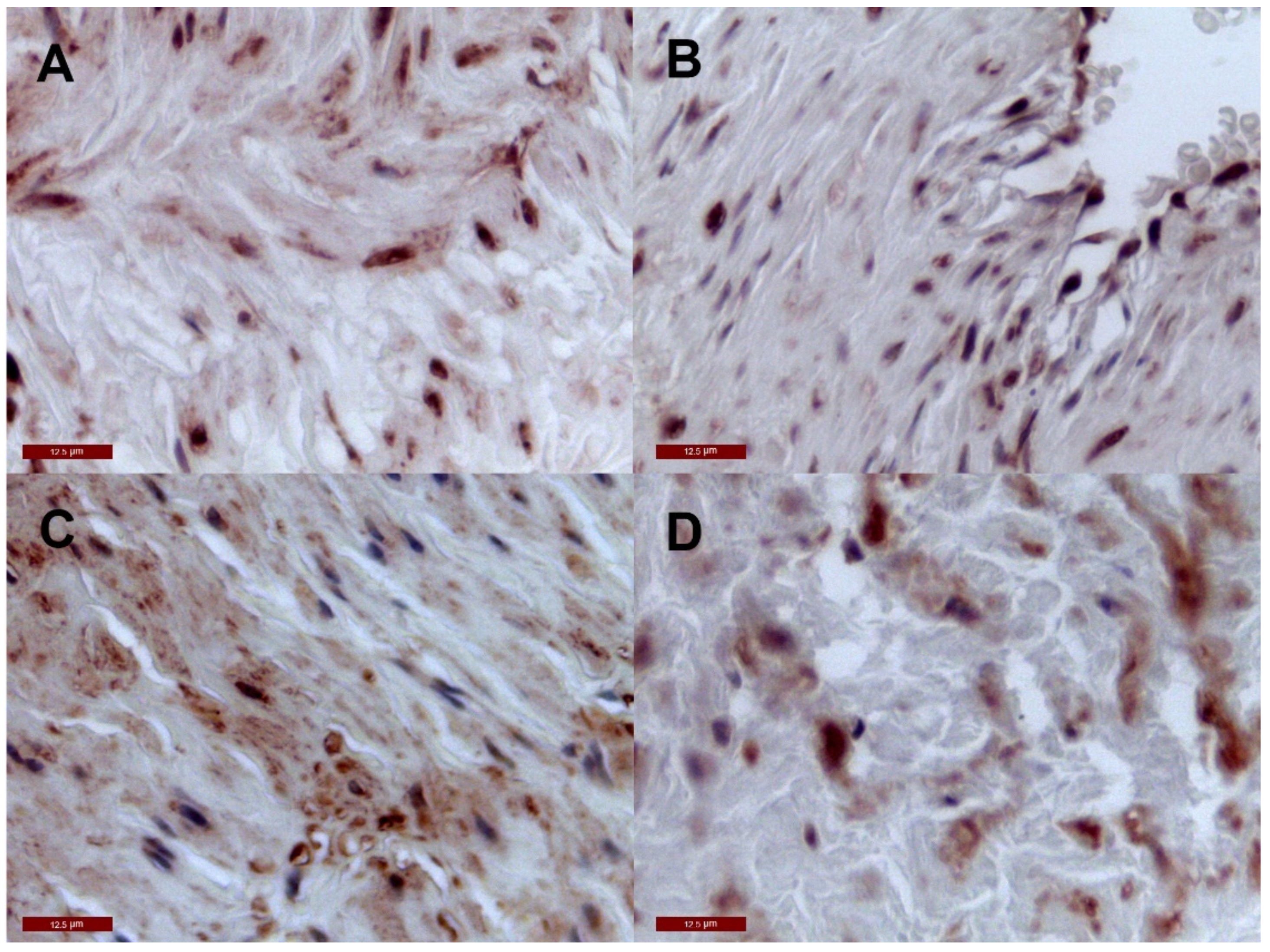
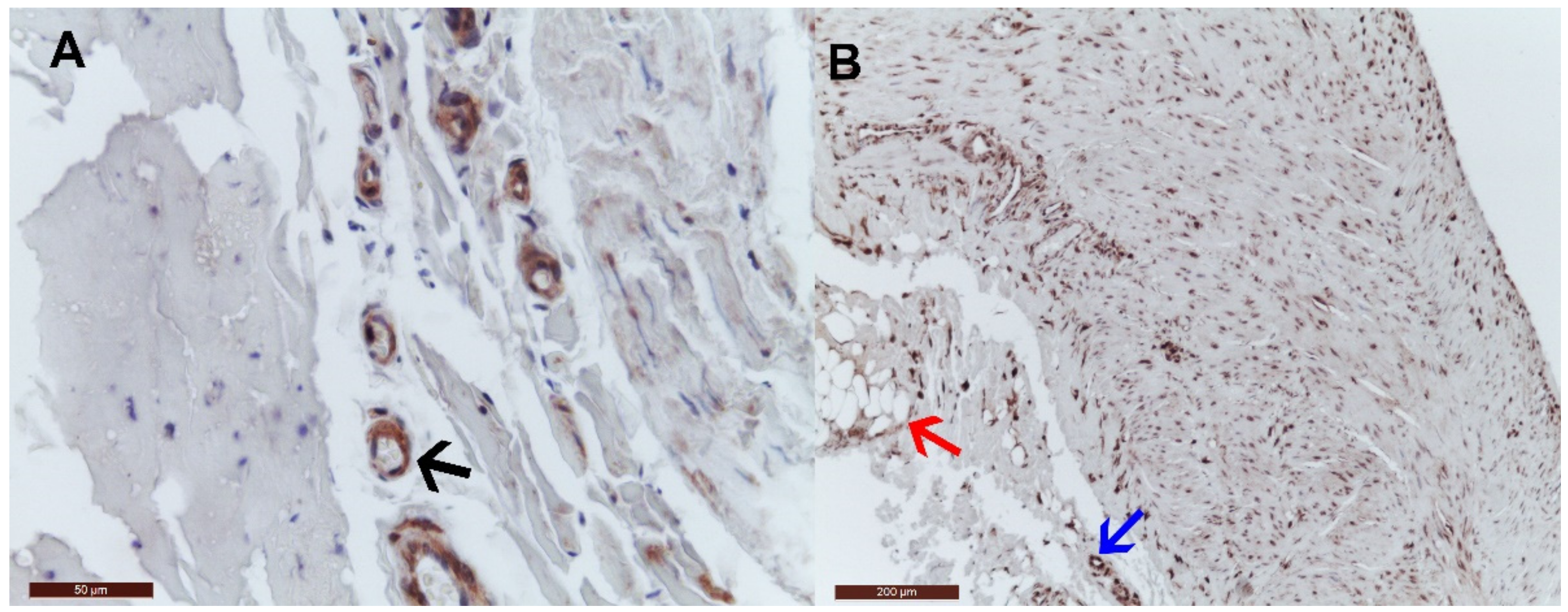
2.2. Immunohistochemistry Tissue Staining
2.3. Plasma tMGP Assessment
2.4. Statistical Analysis
3. Results
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Labropoulos, N.; Tzogias, L.; Malgor, R.D.; Antoniou, G.; Giannoukas, A.D. Phlebosclerosis in lower extremities veins. Phlebology 2014, 29, 186–190. [Google Scholar] [CrossRef] [PubMed]
- Ortega, M.A.; Asúnsolo, Á.; Pekarek, L.; Alvarez-Mon, M.A.; Delforge, A.; Sáez, M.A.; Coca, S.; Sainz, F.; Álvarez-Mon, M.; Buján, J.; et al. Histopathological study of JNK in venous wall of patients with chronic venous insufficiency related to osteogenesis process. Int. J. Med. Sci. 2021, 2021, 1921–1934. [Google Scholar] [CrossRef] [PubMed]
- Cleanthis, M.; Lees, T. Varicose veins and chronic venous insufficiency. In Postgraduate Vascular Surgery. The Candidate’s Guide to the FRCS; Cambridge University Press: Cambridge, UK, 2011; pp. 204–219. [Google Scholar] [CrossRef]
- Oklu, R.; Habito, R.; Mayr, M.; Deipolyi, A.R.; Albadawi, H.; Hesketh, R.; Walker, T.G.; Linskey, K.R.; Long, C.A.; Wicky, S.; et al. Pathogenesis of varicose veins. J. Vasc. Interv. Radiol. 2012, 23, 33–39. [Google Scholar] [CrossRef] [PubMed]
- Cario-Toumaniantz, C.; Boularan, C.; Schurgers, L.J.; Heymann, M.F.; M, L.C.; Léger, J.; Loirand, G.; Pacaud, P. Identification of differentially expressed genes in human varicose veins: Involvement of matrix Gla protein in extracellular matrix remodeling. J. Vasc. Res. 2007, 44, 444–459. [Google Scholar] [CrossRef] [PubMed]
- Schurgers, L.J.; Teunissen, K.J.F.; Knapen, M.H.J.; Kwaijtaal, M.; Van Diest, R.; Appels, A.; Reutelingsperger, C.P.; Cleutjens, J.P.M.; Vermeer, C. Novel Conformation-Specific Antibodies Against Matrix γ-Carboxyglutamic Acid (Gla) Protein Undercarboxylated Matrix Gla Protein as Marker for Vascular Calcification. Arter. Thromb. Vasc. Biol. 2005, 25, 1629–1633. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Silaghi, C.N.; Olteanu, G.; Crăciun, A.M. The behavior of circulating matrix Gla protein, matrix metalloproteinase-9 and nitrotyrosine in patients with varicose veins. Hum. Vet. Med. 2015, 7, 65–69. [Google Scholar]
- He, X.; Wang, Z.; Wei, L.; Cheng, X.; Chen, L.; Gao, F.; Jiang, H. Indoxyl sulfate promotes osteogenic differentiation of vascular smooth muscle cells by miR-155-5p-dependent downregulation of matrix Gla protein via ROS/NF-κB signaling. Exp. Cell Res. 2020, 397, 112301. [Google Scholar] [CrossRef] [PubMed]
- Schurgers, L.J.; Spronk, H.M.H.; Skepper, J.N.; Hackeng, T.M.; Shanahan, C.M.; Vermeer, C.; Weissberg, P.L.; Proudfoot, D. Post-translational modifications regulate matrix Gla protein function: Importance for inhibition of vascular smooth muscle cell calcification. J. Thromb. Haemost. 2007, 5, 2503–2511. [Google Scholar] [CrossRef]
- Roumeliotis, S.; Dounousi, E.; Salmas, M.; Eleftheriadis, T.; Liakopoulos, V. Vascular Calcification in Chronic Kidney Disease: The Role of Vitamin K- Dependent Matrix Gla Protein. Front. Med. 2020, 7, 154. [Google Scholar] [CrossRef]
- Shroff, R.C.; Shanahan, C.M. The vascular biology of calcification. Semin. Dial. 2007, 20, 103–109. [Google Scholar] [CrossRef]
- Zebboudj, A.F.; Imura, M.; Boström, K. Matrix GLA protein, a regulatory protein for bone morphogenetic protein-2. J. Biol. Chem. 2002, 277, 4388–4394. [Google Scholar] [CrossRef] [Green Version]
- Schurgers, L.J. Vitamin K: Key vitamin in controlling vascular calcification in chronic kidney disease. Kidney Int. 2013, 83, 782–784. [Google Scholar] [CrossRef] [Green Version]
- Yao, J.; Guihard, P.J.; Blazquez-Medela, A.M.; Guo, Y.; Liu, T.; Boström, K.I.; Yao, Y. Matrix Gla protein regulates differentiation of endothelial cells derived from mouse embryonic stem cells. Angiogenesis 2016, 19, 1–7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Williams, J.K.; Heistad, D.D. Structure and function of vasa vasorum. Trends Cardiovasc. Med. 1996, 6, 53–57. [Google Scholar] [CrossRef]
- Li, C.; Li, J.; He, F.; Li, K.; Li, X.; Zhang, Y. Matrix Gla protein regulates adipogenesis and is serum marker of visceral adiposity. Adipocyte 2020, 9, 68–76. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pereira, S.S.; Alvarez-Leite, J.I. Adipokines: Biological functions and metabolically healthy obese profile. J. Receptor Ligand Channel Res. 2014, 7, 15–25. [Google Scholar] [CrossRef] [Green Version]
- Silaghi, C.N.; Fodor, D.; Gheorghe, S.R.; Crăciun, A.M. Serum total matrix Gla protein: Reference interval in healthy adults and variations in patients with vascular and osteoarticular diseases. Clin. Chim. Acta 2019, 490, 128–134. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| All (n = 20) | |
|---|---|
| Anthropometric and clinical characteristics | |
| Gender, male/female | 7/13 |
| Age, years | 46.42 ± 8.59 |
| BMI, kg/m2 | 31 ± 5 |
| T2DM, n (%) | 2 (10) |
| RD, n (%) | 2 (10) |
| CVD, n (%) | 4 (20) |
| HT, n (%) | 18 (90) |
| Smokers, n (%) | 8 (40) |
| Biochemical analysis | |
| tMGP, µg/L | |
| before surgery | 59.5 ± 17.2 |
| after surgery | 38.1 ± 11.3 |
| cMGP | ucMGP | pMGP | dpMGP | vK | |
|---|---|---|---|---|---|
| Media | |||||
| All (n = 40) | 38 (95) | 0 (0) | 39 (97.5) | 0 (0) | 0 (0) |
| Control (n = 20) | 20 (100) | 0 (0) | 19 (95) | 0 (0) | 0 (0) |
| VV (n = 20) | 18 (90) | 0 (0) | 20 (100) | 0 (0) | 0 (0) |
| Intima | |||||
| All (n = 40) | 33 (82.5) | 0 (0) | 40 (100) | 0 (0) | 0 (0) |
| Control (n = 20) | 16 (80) | 0 (0) | 20 (100) | 0 (0) | 0 (0) |
| VV (n = 20) | 17 (85) | 0 (0) | 20 (100) | 0 (0) | 0 (0) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gheorghe, S.R.; Vermeer, C.; Olteanu, G.; Silaghi, C.N.; Crăciun, A.M. The Active Isoforms of MGP Are Expressed in Healthy and Varicose Veins without Calcification. J. Clin. Med. 2021, 10, 5896. https://doi.org/10.3390/jcm10245896
Gheorghe SR, Vermeer C, Olteanu G, Silaghi CN, Crăciun AM. The Active Isoforms of MGP Are Expressed in Healthy and Varicose Veins without Calcification. Journal of Clinical Medicine. 2021; 10(24):5896. https://doi.org/10.3390/jcm10245896
Chicago/Turabian StyleGheorghe, Simona R., Cees Vermeer, Gabriel Olteanu, Ciprian N. Silaghi, and Alexandra M. Crăciun. 2021. "The Active Isoforms of MGP Are Expressed in Healthy and Varicose Veins without Calcification" Journal of Clinical Medicine 10, no. 24: 5896. https://doi.org/10.3390/jcm10245896
APA StyleGheorghe, S. R., Vermeer, C., Olteanu, G., Silaghi, C. N., & Crăciun, A. M. (2021). The Active Isoforms of MGP Are Expressed in Healthy and Varicose Veins without Calcification. Journal of Clinical Medicine, 10(24), 5896. https://doi.org/10.3390/jcm10245896