
Targeting Aggressive Pituitary Adenomas at the Molecular Level—A Review

 ,
, 
 and
and 
Abstract
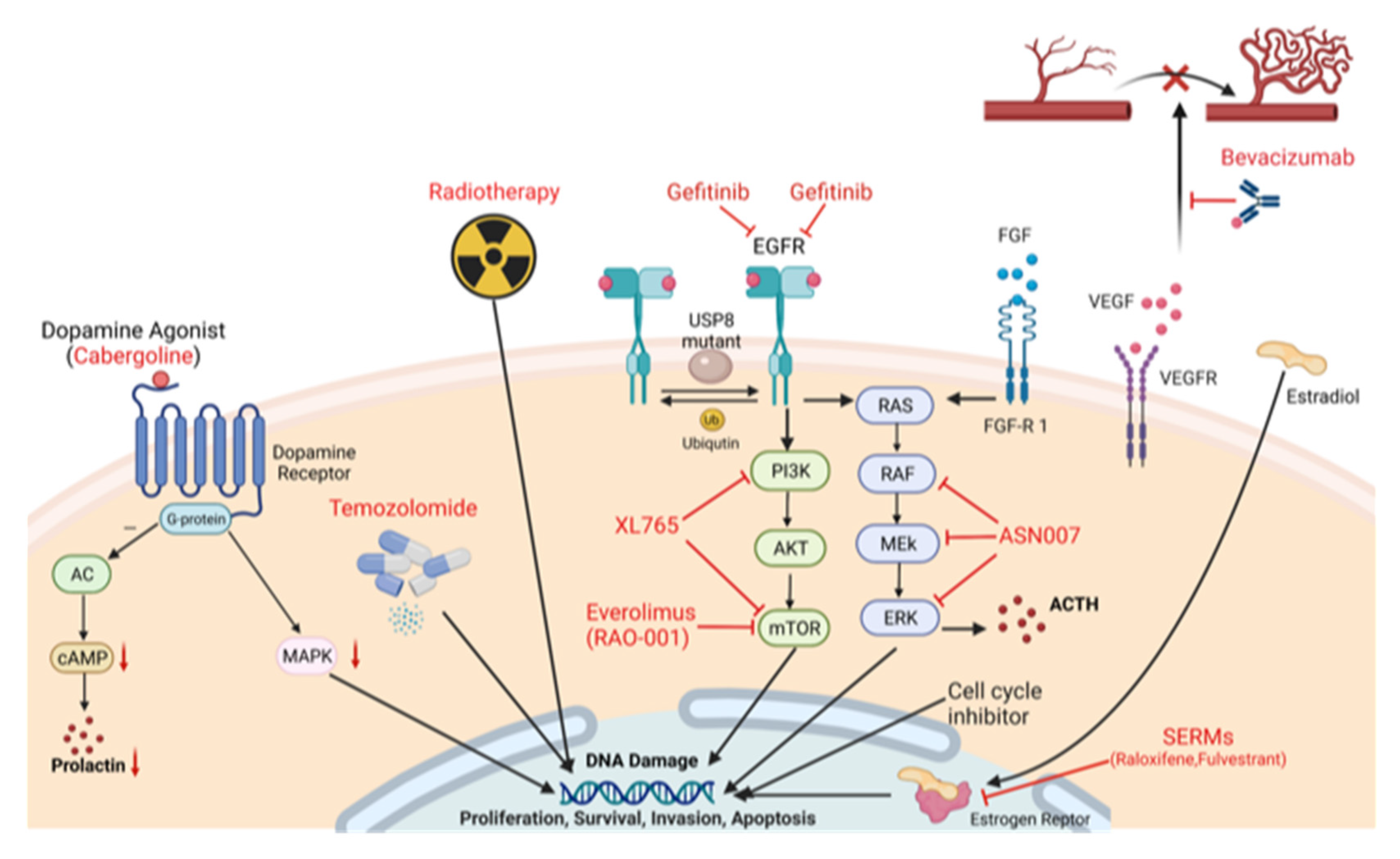
:1. Introduction
2. Emerging Targeted Treatment Strategies at the Molecular Level
2.1. Dopamine Agonist (DA) Treatment, and Treatment of Excess Hormone Secretion
2.2. Epidermal Growth Factor-Receptor (EGFR) Inhibition
2.3. Estrogen Receptor Modulation
2.4. Mammalian Target of Rapamycin (mTOR) Inhibition
2.5. Metalloprotease Inhibition
2.6. Peptide Receptor Radionuclide Therapy (PRRT)
2.7. Vascular Endothelial Growth Factor (VEGF) Inhibition
2.8. Fibroblast Growth Factor (FGF)
2.9. Raf/MEK/ERK Pathway
3. Non-Targeted Treatment
3.1. TMZ and 5-Fluorouracil (5-FU) Treatment
3.2. TMZ and Radiotherapy
4. Checkpoint Inhibition
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Fahlbusch, R.; Buchfelder, M. Pituitary surgery. In The Pituitary; Academic Press: London, UK, 2011; pp. 703–719. [Google Scholar]
- Scheithauer, B.W.; Kovacs, K.T.; Laws, E.R., Jr.; Randall, R.V. Pathology of invasive pituitary tumors with special reference to functional classification. J. Neurosurg. 1986, 65, 733–744. [Google Scholar] [CrossRef] [Green Version]
- Hardy, J.; Vezina, J.L. Transsphenoidal neurosurgery of intracranial neoplasm. Adv. Neurol. 1976, 15, 261–273. [Google Scholar]
- Knosp, E.; Steiner, E.; Kitz, K.; Matula, C. Pituitary adenomas with invasion of the cavernous sinus space: A magnetic resonance imaging classification compared with surgical findings. Neurosurgery 1993, 33, 610–617; discussion 617–618. [Google Scholar] [CrossRef]
- Mete, O.; Lopes, M.B. Overview of the 2017 WHO classification of pituitary tumors. Endocr. Pathol. 2017, 28, 228–243. [Google Scholar] [CrossRef]
- Raverot, G.; Burman, P.; McCormack, A.; Heaney, A.; Petersenn, S.; Popovic, V.; Trouillas, J.; Dekkers, O.M. European Society of Endocrinology Clinical Practice Guidelines for the management of aggressive pituitary tumours and carcinomas. Eur. J. Endocrinol. 2018, 178, G1–G24. [Google Scholar] [CrossRef] [PubMed]
- Trouillas, J.; Roy, P.; Sturm, N.; Dantony, E.; Cortet-Rudelli, C.; Viennet, G.; Bonneville, J.F.; Assaker, R.; Auger, C.; Brue, T.; et al. A new prognostic clinicopathological classification of pituitary adenomas: A multicentric case-control study of 410 patients with 8 years post-operative follow-up. Acta Neuropathol. 2013, 126, 123–135. [Google Scholar] [CrossRef] [PubMed]
- Miermeister, C.P.; Petersenn, S.; Buchfelder, M.; Fahlbusch, R.; Lüdecke, D.K.; Hölsken, A.; Bergmann, M.; Knappe, H.U.; Hans, V.H.; Flitsch, J.; et al. Histological criteria for atypical pituitary adenomas—Data from the German pituitary adenoma registry suggests modifications. Acta Neuropathol. Commun. 2015, 3, 50. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, A.; Liu, W.; Cao, P.; Zheng, Y.; Bu, Z.; Zhou, T. Endoscopic Versus Microscopic Transsphenoidal Surgery in the Treatment of Pituitary Adenoma: A Systematic Review and Meta-Analysis. World Neurosurg. 2017, 101, 236–246. [Google Scholar] [CrossRef]
- Nimsky, C.; von Keller, B.; Ganslandt, O.; Fahlbusch, R. Intraoperative high-field magnetic resonance imaging in transsphenoidal surgery of hormonally inactive pituitary macroadenomas. Neurosurgery 2006, 59, 105–114; discussion 105–114. [Google Scholar] [CrossRef]
- Carl, B.; Bopp, M.; Voellger, B.; Saß, B.; Nimsky, C. Augmented Reality in Transsphenoidal Surgery. World Neurosurg. 2019, 125, e873–e883. [Google Scholar] [CrossRef]
- Trainer, P.J.; Drake, W.M.; Katznelson, L.; Freda, P.U.; Herman-Bonert, V.; van der Lely, A.J.; Dimaraki, E.V.; Stewart, P.M.; Friend, K.E.; Vance, M.L.; et al. Treatment of Acromegaly with the Growth Hormone-Receptor Antagonist Pegvisomant. N. Engl. J. Med. 2000, 342, 1171–1177. [Google Scholar] [CrossRef]
- Feelders, R.A.; Newell-Price, J.; Pivonello, R.; Nieman, L.K.; Hofland, L.J.; Lacroix, A. Advances in the medical treatment of Cushing’s syndrome. Lancet Diabetes Endocrinol. 2019, 7, 300–312. [Google Scholar] [CrossRef]
- Tritos, N.A.; Biller, B.M.K. Medical Management of Cushing Disease. Neurosurg. Clin. N. Am. 2019, 30, 499–508. [Google Scholar] [CrossRef]
- Webb, S.M.; Ware, J.E.; Forsythe, A.; Yang, M.; Badia, X.; Nelson, L.M.; Signorovitch, J.E.; McLeod, L.; Maldonado, M.; Zgliczynski, W.; et al. Treatment effectiveness of pasireotide on health-related quality of life in patients with Cushing’s disease. Eur. J. Endocrinol. 2014, 171, 89–98. [Google Scholar] [CrossRef] [Green Version]
- Lacroix, A.; Gu, F.; Gallardo, W.; Pivonello, R.; Yu, Y.; Witek, P.; Boscaro, M.; Salvatori, R.; Yamada, M.; Tauchmanova, L.; et al. Efficacy and safety of once-monthly pasireotide in Cushing’s disease: A 12 month clinical trial. Lancet Diabetes Endocrinol. 2018, 6, 17–26. [Google Scholar] [CrossRef]
- Pivonello, R.; Ferone, D.; de Herder, W.W.; Kros, J.M.; Del Basso De Caro, M.L.; Arvigo, M.; Annunziato, L.; Lombardi, G.; Colao, A.; Hofland, L.J.; et al. Dopamine Receptor Expression and Function in Corticotroph Pituitary Tumors. J. Clin. Endocrinol. Metab. 2004, 89, 2452–2462. [Google Scholar] [CrossRef] [Green Version]
- Petrossians, P.; Ronci, N.; Valdés Socin, H.; Kalife, A.; Stevenaert, A.; Bloch, B.; Tabarin, A.; Beckers, A. ACTH silent adenoma shrinking under cabergoline. Eur. J. Endocrinol. 2001, 144, 51–57. [Google Scholar] [CrossRef] [Green Version]
- Kunasegaran, S.; Croxson, M.S.; Holdaway, I.; Murphy, R. An unusual case of Cushing’s syndrome due to bihormonal ACTH-prolactin secreting pituitary macroadenoma with rapid response to cabergoline. BMJ Case Rep. 2017, 2017, bcr-2017-219921. [Google Scholar] [CrossRef]
- Manavela, M.P.; Danilowicz, K.; Bruno, O.D. Macrocorticotropinoma shrinkage and control of hypercortisolism under long-term cabergoline therapy: Case report. Pituitary 2012, 15 (Suppl. 1), 33–36. [Google Scholar] [CrossRef]
- Nakhleh, A.; Saiegh, L.; Reut, M.; Ahmad, M.S.; Pearl, I.W.; Shechner, C. Cabergoline treatment for recurrent Cushing’s disease during pregnancy. Hormones 2016, 15, 453–458. [Google Scholar] [CrossRef] [Green Version]
- Sek, K.S.; Deepak, D.S.; Lee, K.O. Use of cabergoline for the management of persistent Cushing’s disease in pregnancy. BMJ Case Rep. 2017, 2017, bcr-2016-217855. [Google Scholar] [CrossRef]
- Miyoshi, T.; Otsuka, F.; Takeda, M.; Inagaki, K.; Suzuki, J.; Ogura, T.; Date, I.; Hashimoto, K.; Makino, H. Effect of cabergoline treatment on Cushing’s disease caused by aberrant adrenocorticotropin-secreting macroadenoma. J. Endocrinol. Investig. 2004, 27, 1055–1059. [Google Scholar] [CrossRef]
- Burman, P.; Edén-Engström, B.; Ekman, B.; Karlsson, F.A.; Schwarcz, E.; Wahlberg, J. Limited value of cabergoline in Cushing’s disease: A prospective study of a 6-week treatment in 20 patients. Eur. J. Endocrinol. 2016, 174, 17–24. [Google Scholar] [CrossRef] [Green Version]
- Godbout, A.; Manavela, M.; Danilowicz, K.; Beauregard, H.; Bruno, O.D.; Lacroix, A. Cabergoline monotherapy in the long-term treatment of Cushing’s disease. Eur. J. Endocrinol. 2010, 163, 709–716. [Google Scholar] [CrossRef]
- Clinicaltrials.gov. Available online: https://clinicaltrials.gov (accessed on 6 November 2021).
- Pivonello, R.; Matrone, C.; Filippella, M.; Cavallo, L.M.; Di Somma, C.; Cappabianca, P.; Colao, A.; Annunziato, L.; Lombardi, G. Dopamine receptor expression and function in clinically nonfunctioning pituitary tumors: Comparison with the effectiveness of cabergoline treatment. J. Clin. Endocrinol. Metab. 2004, 89, 1674–1683. [Google Scholar] [CrossRef]
- Gabalec, F.; Beranek, M.; Netuka, D.; Masopust, V.; Nahlovsky, J.; Cesak, T.; Marek, J.; Cap, J. Dopamine 2 receptor expression in various pathological types of clinically non-functioning pituitary adenomas. Pituitary 2012, 15, 222–226. [Google Scholar] [CrossRef]
- Gabalec, F.; Drastikova, M.; Cesak, T.; Netuka, D.; Masopust, V.; Machac, J.; Marek, J.; Cap, J.; Beranek, M. Dopamine 2 and somatostatin 1-5 receptors coexpression in clinically non-functioning pituitary adenomas. Physiol. Res. 2015, 64, 369–377. [Google Scholar] [CrossRef]
- Vieira Neto, L.; Wildemberg, L.E.; Moraes, A.B.; Colli, L.M.; Kasuki, L.; Marques, N.V.; Gasparetto, E.L.; de Castro, M.; Takiya, C.M.; Gadelha, M.R. Dopamine receptor subtype 2 expression profile in nonfunctioning pituitary adenomas and in vivo response to cabergoline therapy. Clin. Endocrinol. 2015, 82, 739–746. [Google Scholar] [CrossRef]
- Garcia, E.C.; Naves, L.A.; Silva, A.O.; de Castro, L.F.; Casulari, L.A.; Azevedo, M.F. Short-term treatment with cabergoline can lead to tumor shrinkage in patients with nonfunctioning pituitary adenomas. Pituitary 2013, 16, 189–194. [Google Scholar] [CrossRef]
- Greenman, Y.; Cooper, O.; Yaish, I.; Robenshtok, E.; Sagiv, N.; Jonas-Kimchi, T.; Yuan, X.; Gertych, A.; Shimon, I.; Ram, Z.; et al. Treatment of clinically nonfunctioning pituitary adenomas with dopamine agonists. Eur. J. Endocrinol. 2016, 175, 63–72. [Google Scholar] [CrossRef]
- Batista, R.L.; Musolino, N.R.C.; Cescato, V.A.S.; da Silva, G.O.; Medeiros, R.S.S.; Herkenhoff, C.G.B.; Trarbach, E.B.; Cunha-Neto, M.B. Cabergoline in the Management of Residual Nonfunctioning Pituitary Adenoma: A Single-Center, Open-Label, 2-Year Randomized Clinical Trial. Am. J. Clin. Oncol. 2019, 42, 221–227. [Google Scholar] [CrossRef]
- LeRiche, V.K.; Asa, S.L.; Ezzat, S. Epidermal growth factor and its receptor (EGF-R) in human pituitary adenomas: EGF-R correlates with tumor aggressiveness. J. Clin. Endocrinol. Metab. 1996, 81, 656–662. [Google Scholar] [CrossRef] [Green Version]
- Ben-Jonathan, N.; Chen, S.; Dunckley, J.A.; LaPensee, C.; Kansra, S. Estrogen Receptor-α Mediates the Epidermal Growth Factor-Stimulated Prolactin Expression and Release in Lactotrophs. Endocrinology 2009, 150, 795–802. [Google Scholar] [CrossRef] [Green Version]
- Fukuoka, H.; Cooper, O.; Mizutani, J.; Tong, Y.; Ren, S.-G.; Bannykh, S.; Melmed, S. HER2/ErbB2 Receptor Signaling in Rat and Human Prolactinoma Cells: Strategy for Targeted Prolactinoma Therapy. Mol. Endocrinol. 2011, 25, 92–103. [Google Scholar] [CrossRef] [Green Version]
- Fukuoka, H.; Cooper, O.; Ben-Shlomo, A.; Mamelak, A.; Ren, S.G.; Bruyette, D.; Melmed, S. EGFR as a therapeutic target for human, canine, and mouse ACTH-secreting pituitary adenomas. J. Clin. Investig. 2011, 121, 4712–4721. [Google Scholar] [CrossRef] [Green Version]
- Chen, S.; Bangaru, M.L.; Sneade, L.; Dunckley, J.A.; Ben-Jonathan, N.; Kansra, S. Epidermal growth factor receptor cross-talks with ligand-occupied estrogen receptor-alpha to modulate both lactotroph proliferation and prolactin gene expression. Am. J. Physiol. Enocrinol. Metab. 2009, 297, E331–E339. [Google Scholar] [CrossRef] [Green Version]
- Cooper, O.; Mamelak, A.; Bannykh, S.; Carmichael, J.; Bonert, V.; Lim, S.; Cook-Wiens, G.; Ben-Shlomo, A. Prolactinoma ErbB receptor expression and targeted therapy for aggressive tumors. Endocrine 2014, 46, 318–327. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.; Kano, M.; Araki, T.; Cooper, O.; Fukuoka, H.; Tone, Y.; Tone, M.; Melmed, S. ErbB receptor-driven prolactinomas respond to targeted lapatinib treatment in female transgenic mice. Endocrinology 2015, 156, 71–79. [Google Scholar] [CrossRef] [Green Version]
- Liu, Q.; Yu, S.; Zhao, W.; Qin, S.; Chu, Q.; Wu, K. EGFR-TKIs resistance via EGFR-independent signaling pathways. Mol. Cancer 2018, 17, 53. [Google Scholar] [CrossRef] [Green Version]
- Wu, S.G.; Shih, J.Y. Management of acquired resistance to EGFR TKI-targeted therapy in advanced non-small cell lung cancer. Mol. Cancer 2018, 17, 38. [Google Scholar] [CrossRef]
- Medina, P.J.; Goodin, S. Lapatinib: A dual inhibitor of human epidermal growth factor receptor tyrosine kinases. Clin. Ther. 2008, 30, 1426–1447. [Google Scholar] [CrossRef]
- Cooper, O.; Bonert, V.S.; Rudnick, J.; Pressman, B.D.; Lo, J.; Salvatori, R.; Yuen, K.C.J.; Fleseriu, M.; Melmed, S. EGFR/ErbB2-Targeting Lapatinib Therapy for Aggressive Prolactinomas. J. Clin. Endocrinol. Metab. 2021, 106, e917–e925. [Google Scholar] [CrossRef]
- Reincke, M.; Sbiera, S.; Hayakawa, A.; Theodoropoulou, M.; Osswald, A.; Beuschlein, F.; Meitinger, T.; Mizuno-Yamasaki, E.; Kawaguchi, K.; Saeki, Y.; et al. Mutations in the deubiquitinase gene USP8 cause Cushing’s disease. Nat. Genet. 2015, 47, 31–38. [Google Scholar] [CrossRef]
- Ma, Z.Y.; Song, Z.J.; Chen, J.H.; Wang, Y.F.; Li, S.Q.; Zhou, L.F.; Mao, Y.; Li, Y.M.; Hu, R.G.; Zhang, Z.Y.; et al. Recurrent gain-of-function USP8 mutations in Cushing’s disease. Cell Res. 2015, 25, 306–317. [Google Scholar] [CrossRef]
- Dahlman-Wright, K.; Cavailles, V.; Fuqua, S.A.; Jordan, V.C.; Katzenellenbogen, J.A.; Korach, K.S.; Maggi, A.; Muramatsu, M.; Parker, M.G.; Gustafsson, J.A. International Union of Pharmacology. LXIV. Estrogen receptors. Pharmacol. Rev. 2006, 58, 773–781. [Google Scholar] [CrossRef] [Green Version]
- Björnström, L.; Sjöberg, M. Mechanisms of estrogen receptor signaling: Convergence of genomic and nongenomic actions on target genes. Mol. Endocrinol. 2005, 19, 833–842. [Google Scholar] [CrossRef] [Green Version]
- Patel, H.K.; Bihani, T. Selective estrogen receptor modulators (SERMs) and selective estrogen receptor degraders (SERDs) in cancer treatment. Pharmacol. Ther. 2018, 186, 1–24. [Google Scholar] [CrossRef]
- Wright, D.J.; Earnhardt, J.N.; Perry, R.; Bailey, S.; Komm, B.; Minck, D.R.; Cukierski, M.A. Carcinogenicity and hormone studies with the tissue-selective estrogen receptor modulator bazadoxifene. J. Cell. Physiol. 2013, 228, 724–733. [Google Scholar] [CrossRef]
- Hannen, R.; Steffani, M.; Voellger, B.; Carl, B.; Wang, J.; Bartsch, J.W.; Nimsky, C. Effects of anti-estrogens on cell invasion and survival in pituitary adenoma cells: A systematic study. J. Steroid Biochem. Mol. Biol. 2019, 187, 88–96. [Google Scholar] [CrossRef]
- Walker, A.B.; Eldridge, P.R.; MacFarlane, I.A. Clomiphene-induced pituitary apoplexy in a patient with acromegaly. Postgrad. Med. J. 1996, 72, 172–173. [Google Scholar] [CrossRef] [Green Version]
- Dimaraki, E.V.; Symons, K.V.; Barkan, A.L. Raloxifene decreases serum IGF-I in male patients with active acromegaly. Eur. J. Endocrinol. 2004, 150, 481–487. [Google Scholar] [CrossRef] [Green Version]
- Choudhary, C.; Hamrahian, A.H.; Bena, J.F.; Recinos, P.; Kennedy, L.; Dobri, G. The Effect of Raloxifene on Serum Prolactin Level in Patients with Prolactinoma. Endocr. Pract. 2019, 25, 684–688. [Google Scholar] [CrossRef] [PubMed]
- Lamberts, S.W.; de Quijada, M.; Klijn, J.G. The effect of tamoxifen on GH and PRL secretion by human pituitary tumors. J. Endocrinol. Investig. 1980, 3, 343–347. [Google Scholar] [CrossRef] [PubMed]
- Lamberts, S.W.; Verleun, T.; Oosterom, R. Effect of tamoxifen administration on prolactin release by invasive prolactin-secreting pituitary adenomas. Neuroendocrinology 1982, 34, 339–342. [Google Scholar] [CrossRef] [PubMed]
- Christian, Z.; Hatanpaa, K.; Auchus, R.; Hammes, S.; Patel, A.; Mickey, B. Dopamine Agonist and Tamoxifen Combination Therapy for a Prolactin-Secreting Pituitary Tumor Resistant to Dopamine Agonist Monotherapy: Case Report and Review. Interdiscip. Neurosurg. 2020, 21, 100777. [Google Scholar] [CrossRef]
- Voellger, B.; Kirches, E.; Wilisch-Neumann, A.; Weise, A.; Tapia-Perez, J.H.; Rupa, R.; Mawrin, C.; Firsching, R. Resveratrol decreases B-cell lymphoma-2 expression and viability in GH3 pituitary adenoma cells of the rat. OncoTargets Ther. 2013, 9, 1269–1276. [Google Scholar] [CrossRef] [Green Version]
- Voellger, B.; Waldt, N.; Rupa, R.; Kirches, E.; Melhem, O.; Ochel, H.J.; Mawrin, C.; Firsching, R. Combined effects of resveratrol and radiation in GH3 and TtT/GF pituitary adenoma cells. J. Neurooncol. 2018, 139, 573–582. [Google Scholar] [CrossRef]
- Zhang, Z.; Bartsch, J.W.; Benzel, J.; Lei, T.; Nimsky, C.; Voellger, B. Selective estrogen receptor modulators decrease invasiveness in pituitary adenoma cell lines AtT-20 and TtT/GF by affecting expression of MMP-14 and ADAM12. FEBS Open Bio 2020, 10, 2489–2498. [Google Scholar] [CrossRef]
- Takayasu, S.; Usutani, M.; Makita, K.; Daimon, M. The activation of G protein-coupled receptor 30 increases pro-opiomelanocortin gene expression through cAMP/PKA/NR4A pathway in mouse pituitary corticotroph AtT-20 cells. Neurosci. Lett. 2020, 739, 135468. [Google Scholar] [CrossRef]
- Dang, V.H.; Choi, K.C.; Jeung, E.B. Membrane-impermeable estrogen is involved in regulation of calbindin-D9k expression via non-genomic pathways in a rat pituitary cell line, GH3 cells. Toxicol. In Vitro 2010, 24, 1229–1236. [Google Scholar] [CrossRef]
- Viñas, R.; Watson, C.S. Bisphenol S disrupts estradiol-induced nongenomic signaling in a rat pituitary cell line: Effects on cell functions. Environ. Health Perspect. 2013, 121, 352–358. [Google Scholar] [CrossRef] [PubMed]
- Watson, C.S.; Jeng, Y.J.; Hu, G.; Wozniak, A.; Bulayeva, N.; Guptarak, J. Estrogen- and xenoestrogen-induced ERK signaling in pituitary tumor cells involves estrogen receptor-α interactions with G protein-αi and caveolin I. Steroids 2012, 77, 424–432. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Engelman, J.A.; Luo, J.; Cantley, L.C. The evolution of phosphatidylinositol 3-kinases as regulators of growth and metabolism. Nat. Rev. Genet. 2006, 7, 606–619. [Google Scholar] [CrossRef]
- Monsalves, E.; Juraschka, K.; Tateno, T.; Agnihotri, S.; Asa, S.L.; Ezzat, S.; Zadeh, G.J.E.-R.C. The PI3K/AKT/mTOR pathway in the pathophysiology and treatment of pituitary adenomas. Endocr. Relat. Cancer 2014, 21, R331–R344. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Musat, M.; Korbonits, M.; Kola, B.; Borboli, N.; Hanson, M.; Nanzer, A.; Grigson, J.; Jordan, S.; Morris, D.; Gueorguiev, M.; et al. Enhanced protein kinase B/Akt signalling in pituitary tumours. Endocr. Relat. Cancer 2005, 12, 423–433. [Google Scholar] [CrossRef] [Green Version]
- Sajjad, E.A.; Zieliński, G.; Maksymowicz, M.; Hutnik, Ł.; Bednarczuk, T.; Włodarski, P. mTOR is frequently active in GH-secreting pituitary adenomas without influencing their morphopathological features. Endocr. Pathol. 2013, 24, 11–19. [Google Scholar] [CrossRef]
- Jia, W.; Sanders, A.J.; Jia, G.; Liu, X.; Lu, R.; Jiang, W.G. Expression of the mTOR Pathway Regulators in Human Pituitary Adenomas Indicates the Clinical Course. Anticancer Res. 2013, 33, 3123. [Google Scholar]
- Benjamin, D.; Colombi, M.; Moroni, C.; Hall, M.N. Rapamycin passes the torch: A new generation of mTOR inhibitors. J. Nat. Rev. Drug Discov. 2011, 10, 868–880. [Google Scholar] [CrossRef]
- Vilar, E.; Perez-Garcia, J.; Tabernero, J. Pushing the Envelope in the mTOR Pathway: The Second Generation of Inhibitors. Mol. Cancer Ther. 2011, 10, 395. [Google Scholar] [CrossRef] [Green Version]
- Gorshtein, A.; Rubinfeld, H.; Kendler, E.; Theodoropoulou, M.; Cerovac, V.; Stalla, G.K.; Cohen, Z.R.; Hadani, M.; Shimon, I. Mammalian target of rapamycin inhibitors rapamycin and RAD001 (everolimus) induce anti-proliferative effects in GH-secreting pituitary tumor cells in vitro. Endocr.-Relat. Cancer 2009, 16, 1017–1027. [Google Scholar] [CrossRef] [Green Version]
- Di Pasquale, C.; Gentilin, E.; Falletta, S.; Bellio, M.; Buratto, M.; degli Uberti, E.; Chiara Zatelli, M. PI3K/Akt/mTOR pathway involvement in regulating growth hormone secretion in a rat pituitary adenoma cell line. Endocrine 2018, 60, 308–316. [Google Scholar] [CrossRef]
- Zatelli, M.C.; Minoia, M.; Filieri, C.; Tagliati, F.; Buratto, M.; Ambrosio, M.R.; Lapparelli, M.; Scanarini, M.; degli Uberti, E.C. Effect of everolimus on cell viability in nonfunctioning pituitary adenomas. J. Clin. Endocrinol. Metab. 2010, 95, 968–976. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Donovan, L.E.; Arnal, A.V.; Wang, S.H.; Odia, Y. Widely metastatic atypical pituitary adenoma with mTOR pathway STK11(F298L) mutation treated with everolimus therapy. CNS Oncol. 2016, 5, 203–209. [Google Scholar] [CrossRef]
- Zhang, D.; Way, J.S.; Zhang, X.; Sergey, M.; Bergsneider, M.; Wang, M.B.; Yong, W.H.; Heaney, A.P. Effect of Everolimus in Treatment of Aggressive Prolactin-Secreting Pituitary Adenomas. J. Clin. Endocrinol. Metab. 2019, 104, 1929–1936. [Google Scholar] [CrossRef]
- Visse, R.; Nagase, H. Matrix metalloproteinases and tissue inhibitors of metalloproteinases: Structure, function, and biochemistry. Circ. Res. 2003, 92, 827–839. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tallant, C.; Marrero, A.; Gomis-Rüth, F.X. Matrix metalloproteinases: Fold and function of their catalytic domains. Biochim. Biophys. Acta 2010, 1803, 20–28. [Google Scholar] [CrossRef]
- Mochizuki, S.; Okada, Y. ADAMs in cancer cell proliferation and progression. Cancer Sci. 2007, 98, 621–628. [Google Scholar] [CrossRef] [PubMed]
- Gialeli, C.; Theocharis, A.D.; Karamanos, N.K. Roles of matrix metalloproteinases in cancer progression and their pharmacological targeting. FEBS J. 2011, 278, 16–27. [Google Scholar] [CrossRef] [PubMed]
- Roy, R.; Yang, J.; Moses, M.A. Matrix metalloproteinases as novel biomarkers and potential therapeutic targets in human cancer. J. Clin. Oncol. 2009, 27, 5287–5297. [Google Scholar] [CrossRef] [Green Version]
- Liu, W.; Matsumoto, Y.; Okada, M.; Miyake, K.; Kunishio, K.; Kawai, N.; Tamiya, T.; Nagao, S. Matrix metalloproteinase 2 and 9 expression correlated with cavernous sinus invasion of pituitary adenomas. J. Med. Investig. 2005, 52, 151–158. [Google Scholar] [CrossRef] [Green Version]
- Malik, M.T.; Kakar, S.S. Regulation of angiogenesis and invasion by human Pituitary tumor transforming gene (PTTG) through increased expression and secretion of matrix metalloproteinase-2 (MMP-2). Mol. Cancer 2006, 5, 61. [Google Scholar] [CrossRef] [Green Version]
- Beaulieu, E.; Kachra, Z.; Mousseau, N.; Delbecchi, L.; Hardy, J.; Béliveau, R. Matrix metalloproteinases and their inhibitors in human pituitary tumors. Neurosurgery 1999, 45, 1432–1441. [Google Scholar] [CrossRef]
- Turner, H.E.; Nagy, Z.; Esiri, M.M.; Harris, A.L.; Wass, J.A.H. Role of matrix metalloproteinase 9 in pituitary tumor behavior. J. Clin. Endocrinol. Metab. 2000, 85, 2931–2935. [Google Scholar] [CrossRef] [PubMed]
- Gültekin, G.D.; Çabuk, B.; Vural, Ç.; Ceylan, S. Matrix metalloproteinase-9 and tissue inhibitor of matrix metalloproteinase-2: Prognostic biological markers in invasive prolactinomas. J. Clin. Neurosci. 2015, 22, 1282–1287. [Google Scholar] [CrossRef] [PubMed]
- Hussaini, I.M.; Trotter, C.; Zhao, Y.; Abdel-Fattah, R.; Amos, S.; Xiao, A.; Agi, C.U.; Redpath, G.T.; Fang, Z.; Leung, G.K.K.; et al. Matrix metalloproteinase-9 is differentially expressed in nonfunctioning invasive and noninvasive pituitary adenomas and increases invasion in human pituitary adenoma cell line. Am. J. Pathol. 2007, 170, 356–365. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, X.; Feng, M.; Zhang, Y.; Dai, C.; Sun, B.; Bao, X.; Deng, K.; Yao, Y.; Wang, R. Expression of Matrix Metalloproteinase-9, Pituitary Tumor Transforming Gene, High Mobility Group A 2, and Ki-67 in Adrenocorticotropic Hormone-Secreting Pituitary Tumors and Their Association with Tumor Recurrence. World Neurosurg. 2018, 113, e213–e221. [Google Scholar] [CrossRef] [PubMed]
- Knappe, U.J.; Hagel, C.; Lisboa, B.W.; Wilczak, W.; Lüdecke, D.K.; Saeger, W. Expression of serine proteases and metalloproteinases in human pituitary adenomas and anterior pituitary lobe tissue. Acta Neuropathol. 2003, 106, 471–478. [Google Scholar] [CrossRef]
- Pan, Y.; Han, C.; Wang, C.; Hu, G.; Luo, C.; Gan, X.; Zhang, F.; Lu, Y.; Ding, X. ADAM10 promotes pituitary adenoma cell migration by regulating cleavage of CD44 and L1. J. Mol. Endocrinol. 2012, 49, 21–33. [Google Scholar] [CrossRef]
- Wang, J.; Voellger, B.; Benzel, J.; Schlomann, U.; Nimsky, C.; Bartsch, J.W.; Carl, B. Metalloproteinases ADAM12 and MMP-14 are associated with cavernous sinus invasion in pituitary adenomas. Int. J. Cancer 2016, 139, 1327–1339. [Google Scholar] [CrossRef] [Green Version]
- Coussens, L.M.; Fingleton, B.; Matrisian, L.M. Matrix Metalloproteinase Inhibitors and Cancer—Trials and Tribulations. Science 2002, 295, 2387–2392. [Google Scholar] [CrossRef]
- Strosberg, J.; El-Haddad, G.; Wolin, E.; Hendifar, A.; Yao, J.; Chasen, B.; Mittra, E.; Kunz, P.L.; Kulke, M.H.; Jacene, H.; et al. Phase 3 Trial of (177)Lu-Dotatate for Midgut Neuroendocrine Tumors. N. Engl. J. Med. 2017, 376, 125–135. [Google Scholar] [CrossRef]
- Hope, T.A.; Calais, J.; Zhang, L.; Dieckmann, W.; Millo, C. (111)In-Pentetreotide Scintigraphy Versus (68)Ga-DOTATATE PET: Impact on Krenning Scores and Effect of Tumor Burden. J. Nucl. Med. 2019, 60, 1266–1269. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chinezu, L.; Vasiljevic, A.; Jouanneau, E.; François, P.; Borda, A.; Trouillas, J.; Raverot, G. Expression of somatostatin receptors, SSTR2A and SSTR5, in 108 endocrine pituitary tumors using immunohistochemical detection with new specific monoclonal antibodies. Hum. Pathol. 2014, 45, 71–77. [Google Scholar] [CrossRef] [PubMed]
- Komor, J.; Reubi, J.C.; Christ, E.R. Peptide receptor radionuclide therapy in a patient with disabling non-functioning pituitary adenoma. Pituitary 2014, 17, 227–231. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giuffrida, G.; Ferraù, F.; Laudicella, R.; Cotta, O.R.; Messina, E.; Granata, F.; Angileri, F.F.; Vento, A.; Alibrandi, A.; Baldari, S.; et al. Peptide receptor radionuclide therapy for aggressive pituitary tumors: A monocentric experience. Endocr. Connect. 2019, 8, 528–535. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Napoleone, F. Vascular endothelial growth factor: Basic science and clinical progress. J. Endocr. Rev. 2004, 25, 581–611. [Google Scholar] [CrossRef]
- Turner, H.E.; Nagy, Z.; Gatter, K.C.; Esiri, M.M.; Harris, A.L.; Wass, J.A.H. Angiogenesis in pituitary adenomas and the normal pituitary gland. J. Clin. Endocrinol. Metab. 2000, 85, 1159–1162. [Google Scholar] [CrossRef]
- Fukui, S.; Nawashiro, H.; Otani, N.; Ooigawa, H.; Yano, A.; Nomura, N.; Tokumaru, A.M.; Miyazawa, T.; Ohnuki, A.; Tsuzuki, N.; et al. Vascular endothelial growth factor expression in pituitary adenomas. Acta Neurochir. Suppl. 2003, 86, 519–521. [Google Scholar] [CrossRef]
- Ortiz, L.D.; Syro, L.V.; Scheithauer, B.W.; Ersen, A.; Uribe, H.; Fadul, C.E.; Rotondo, F.; Horvath, E.; Kovacs, K. Anti-VEGF Therapy in Pituitary Carcinoma. Pituitary 2012, 15, 445–449. [Google Scholar] [CrossRef]
- Hamerlik, P.; Lathia, J.D.; Rasmussen, R.; Wu, Q.; Bartkova, J.; Lee, M.; Moudry, P.; Bartek, J., Jr.; Fischer, W.; Lukas, J.; et al. Autocrine VEGF-VEGFR2-Neuropilin-1 signaling promotes glioma stem-like cell viability and tumor growth. J. Exp. Med. 2012, 209, 507–520. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; He, Q.; Meng, X.; Zhou, S.; Zhu, Y.; Xu, J.; Tao, R. Apatinib (YN968D1) and Temozolomide in Recurrent Invasive Pituitary Adenoma: Case Report and Literature Review. World Neurosurg. 2019, 124, 319–322. [Google Scholar] [CrossRef]
- Marques, P.; Barry, S.; Carlsen, E.; Collier, D.; Ronaldson, A.; Awad, S.; Dorward, N.; Grieve, J.; Mendoza, N.; Muquit, S.; et al. Pituitary tumour fibroblast-derived cytokines influence tumour aggressiveness. Endocr. Relat. Cancer 2019, 26, 853–865. [Google Scholar] [CrossRef] [PubMed]
- Cheng, S.Q.; Fan, H.Y.; Xu, X.; Gao, W.W.; Lv, S.G.; Ye, M.H.; Wu, M.J.; Shen, X.L.; Cheng, Z.J.; Zhu, X.G.; et al. Over-expression of LRIG1 suppresses biological function of pituitary adenoma via attenuation of PI3K/AKT and Ras/Raf/ERK pathways in vivo and in vitro. J. Huazhong Univ. Sci. Technol. Med. Sci. 2016, 36, 558–563. [Google Scholar] [CrossRef] [PubMed]
- McCormack, A.; Dekkers, O.M.; Petersenn, S.; Popovic, V.; Trouillas, J.; Raverot, G.; Burman, P.; Hubalewska-Dydejezky, A.; Assie, G.; Bach, L.; et al. Treatment of aggressive pituitary tumours and carcinomas: Results of a European Society of Endocrinology (ESE) survey 2016. Eur. J. Endocrinol. 2018, 178, 265–276. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thearle, M.S.; Freda, P.U.; Bruce, J.N.; Isaacson, S.R.; Lee, Y.; Fine, R.L. Temozolomide (Temodar®) and capecitabine (Xeloda®) treatment of an aggressive corticotroph pituitary tumor. Pituitary 2011, 14, 418–424. [Google Scholar] [CrossRef] [Green Version]
- Zacharia, B.E.; Gulati, A.P.; Bruce, J.N.; Carminucci, A.S.; Wardlaw, S.L.; Siegelin, M.; Remotti, H.; Lignelli, A.; Fine, R.L. High response rates and prolonged survival in patients with corticotroph pituitary tumors and refractory Cushing disease from capecitabine and temozolomide (CAPTEM): A case series. Neurosurgery 2014, 74, E447–E455; discussion E455. [Google Scholar] [CrossRef]
- Chakravarti, A.; Erkkinen, M.G.; Nestler, U.; Stupp, R.; Mehta, M.; Aldape, K.; Gilbert, M.R.; Black, P.M.; Loeffler, J.S. Temozolomide-Mediated Radiation Enhancement in Glioblastoma: A Report on Underlying Mechanisms. Clin. Cancer Res. 2006, 12, 4738. [Google Scholar] [CrossRef] [Green Version]
- Stupp, R.; Mason, W.P.; van den Bent, M.J.; Weller, M.; Fisher, B.; Taphoorn, M.J.; Belanger, K.; Brandes, A.A.; Marosi, C.; Bogdahn, U.; et al. Radiotherapy plus concomitant and adjuvant temozolomide for glioblastoma. N. Engl. J. Med. 2005, 352, 987–996. [Google Scholar] [CrossRef]
- Yamamoto, H.; Imai, K. Microsatellite instability: An update. Arch. Toxicol. 2015, 89, 899–921. [Google Scholar] [CrossRef]
- Basu, A.; Ramamoorthi, G.; Jia, Y.; Faughn, J.; Wiener, D.; Awshah, S.; Kodumudi, K.; Czerniecki, B.J. Chapter Six—Immunotherapy in breast cancer: Current status and future directions. In Advances in Cancer Research; Wang, X.-Y., Fisher, P.B., Eds.; Academic Press: Cambridge, MA, USA, 2019; Volume 143, pp. 295–349. [Google Scholar] [CrossRef]
- Heshmati, H.M.; Kujas, M.; Casanova, S.; Wollan, P.C.; Racadot, J.; van Effenterre, R.; Derome, P.J.; Turpin, G. Prevalence of lymphocytic infiltrate in 1400 pituitary adenomas. Endocr. J. 1998, 45, 357–361. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lupi, I.; Manetti, L.; Caturegli, P.; Menicagli, M.; Cosottini, M.; Iannelli, A.; Acerbi, G.; Bevilacqua, G.; Bogazzi, F.; Martino, E. Tumor infiltrating lymphocytes but not serum pituitary antibodies are associated with poor clinical outcome after surgery in patients with pituitary adenoma. J. Clin. Endocrinol. Metab. 2010, 95, 289–296. [Google Scholar] [CrossRef]
- Iacovazzo, D.; Chiloiro, S.; Carlsen, E.; Bianchi, A.; Giampietro, A.; Tartaglione, T.; Bima, C.; Bracaccia, M.E.; Lugli, F.; Lauretti, L.; et al. Tumour-infiltrating cytotoxic T lymphocytes in somatotroph pituitary neuroendocrine tumours. Endocrine 2020, 67, 651–658. [Google Scholar] [CrossRef] [Green Version]
- Wang, P.-F.; Wang, T.-J.; Yang, Y.-K.; Yao, K.; Li, Z.; Li, Y.M.; Yan, C.-X. The expression profile of PD-L1 and CD8+ lymphocyte in pituitary adenomas indicating for immunotherapy. J. Neurooncol. 2018, 139, 89–95. [Google Scholar] [CrossRef]
- Butte, M.J.; Keir, M.E.; Phamduy, T.B.; Sharpe, A.H.; Freeman, G.J. Programmed Death-1 Ligand 1 Interacts Specifically with the B7-1 Costimulatory Molecule to Inhibit T Cell Responses. Immunity 2007, 27, 111–122. [Google Scholar] [CrossRef] [Green Version]
- Suteau, V.; Collin, A.; Menei, P.; Rodien, P.; Rousselet, M.-C.; Briet, C. Expression of programmed death-ligand 1 (PD-L1) in human pituitary neuroendocrine tumor. Cancer Immunol. Immunother. 2020, 69, 2053–2061. [Google Scholar] [CrossRef] [PubMed]
- Mei, Y.; Bi, W.L.; Greenwald, N.F.; Du, Z.; Agar, N.Y.R.; Kaiser, U.B.; Woodmansee, W.W.; Reardon, D.A.; Freeman, G.J.; Fecci, P.E.; et al. Increased expression of programmed death ligand 1 (PD-L1) in human pituitary tumors. Oncotarget 2016, 7, 76565–76576. [Google Scholar] [CrossRef] [Green Version]
- Salomon, M.P.; Wang, X.; Marzese, D.M.; Hsu, S.C.; Nelson, N.; Zhang, X.; Matsuba, C.; Takasumi, Y.; Ballesteros-Merino, C.; Fox, B.A.; et al. The Epigenomic Landscape of Pituitary Adenomas Reveals Specific Alterations and Differentiates Among Acromegaly, Cushing’s Disease and Endocrine-Inactive Subtypes. Clin. Cancer Res. 2018, 24, 4126–4136. [Google Scholar] [CrossRef] [Green Version]
- Kemeny, H.R.; Elsamadicy, A.A.; Farber, S.H.; Champion, C.D.; Lorrey, S.J.; Chongsathidkiet, P.; Woroniecka, K.I.; Cui, X.; Shen, S.H.; Rhodin, K.E.; et al. Targeting PD-L1 Initiates Effective Antitumor Immunity in a Murine Model of Cushing Disease. Clin. Cancer Res. 2020, 26, 1141–1151. [Google Scholar] [CrossRef] [PubMed]
- Reck, M.; Rodríguez-Abreu, D.; Robinson, A.G.; Hui, R.; Csőszi, T.; Fülöp, A.; Gottfried, M.; Peled, N.; Tafreshi, A.; Cuffe, S. Pembrolizumab versus chemotherapy for PD-L1–positive non–small-cell lung cancer. N. Engl. J. Med. 2016, 375, 1823–1833. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weber, J.S.; D’Angelo, S.P.; Minor, D.; Hodi, F.S.; Gutzmer, R.; Neyns, B.; Hoeller, C.; Khushalani, N.I.; Miller, W.H., Jr.; Lao, C.D.; et al. Nivolumab versus chemotherapy in patients with advanced melanoma who progressed after anti-CTLA-4 treatment (CheckMate 037): A randomised, controlled, open-label, phase 3 trial. Lancet Oncol. 2015, 16, 375–384. [Google Scholar] [CrossRef]
- Albarel, F.; Gaudy, C.; Castinetti, F.; Carré, T.; Morange, I.; Conte-Devolx, B.; Grob, J.J.; Brue, T. Long-term follow-up of ipilimumab-induced hypophysitis, a common adverse event of the anti-CTLA-4 antibody in melanoma. Eur. J. Endocrinol. 2015, 172, 195–204. [Google Scholar] [CrossRef] [Green Version]
- Wolchok, J.D.; Chiarion-Sileni, V.; Gonzalez, R.; Rutkowski, P.; Grob, J.J.; Cowey, C.L.; Lao, C.D.; Wagstaff, J.; Schadendorf, D.; Ferrucci, P.F.; et al. Overall Survival with Combined Nivolumab and Ipilimumab in Advanced Melanoma. N. Engl. J. Med. 2017, 377, 1345–1356. [Google Scholar] [CrossRef]
- Majd, N.; Waguespack, S.G.; Janku, F.; Fu, S.; Penas-Prado, M.; Xu, M.; Alshawa, A.; Kamiya-Matsuoka, C.; Raza, S.M.; McCutcheon, I.E.; et al. Efficacy of pembrolizumab in patients with pituitary carcinoma: Report of four cases from a phase II study. J. Immunother. Cancer 2020, 8, e001532. [Google Scholar] [CrossRef] [PubMed]
- Darvin, P.; Toor, S.M.; Sasidharan Nair, V.; Elkord, E. Immune checkpoint inhibitors: Recent progress and potential biomarkers. Exp. Mol. Med. 2018, 50, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Rizvi, N.A.; Hellmann, M.D.; Snyder, A.; Kvistborg, P.; Makarov, V.; Havel, J.J.; Lee, W.; Yuan, J.; Wong, P.; Ho, T.S.; et al. Cancer immunology. Mutational landscape determines sensitivity to PD-1 blockade in non-small cell lung cancer. Science 2015, 348, 124–128. [Google Scholar] [CrossRef] [Green Version]
- Snyder, A.; Makarov, V.; Merghoub, T.; Yuan, J.; Zaretsky, J.M.; Desrichard, A.; Walsh, L.A.; Postow, M.A.; Wong, P.; Ho, T.S.; et al. Genetic basis for clinical response to CTLA-4 blockade in melanoma. N. Engl. J. Med. 2014, 371, 2189–2199. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilky, B.A. Immune checkpoint inhibitors: The linchpins of modern immunotherapy. Immunol. Rev. 2019, 290, 6–23. [Google Scholar] [CrossRef]
- Lin, A.L.; Jonsson, P.; Tabar, V.; Yang, T.J.; Cuaron, J.; Beal, K.; Cohen, M.; Postow, M.; Rosenblum, M.; Shia, J.; et al. Marked Response of a Hypermutated ACTH-Secreting Pituitary Carcinoma to Ipilimumab and Nivolumab. J. Clin. Endocrinol. Metab. 2018, 103, 3925–3930. [Google Scholar] [CrossRef] [Green Version]
- Johnson, B.E.; Mazor, T.; Hong, C.; Barnes, M.; Aihara, K.; McLean, C.Y.; Fouse, S.D.; Yamamoto, S.; Ueda, H.; Tatsuno, K.; et al. Mutational analysis reveals the origin and therapy-driven evolution of recurrent glioma. Science 2014, 343, 189–193. [Google Scholar] [CrossRef] [Green Version]
- Hunter, C.; Smith, R.; Cahill, D.P.; Stephens, P.; Stevens, C.; Teague, J.; Greenman, C.; Edkins, S.; Bignell, G.; Davies, H.; et al. A hypermutation phenotype and somatic MSH6 mutations in recurrent human malignant gliomas after alkylator chemotherapy. Cancer Res. 2006, 66, 3987–3991. [Google Scholar] [CrossRef] [PubMed] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Mechanism of Action | Intervention(s) | Trial Identifier | Study Title | Recruitment Status | Condition | Phase | Number of Enrolled Patients | Study Type | Primary Purpose |
|---|---|---|---|---|---|---|---|---|---|
| checkpoint inhibition | Ipilimumab, Nivolumab | NCT04042753 | Nivolumab and Ipilimumab in People With Aggressive Pituitary Tumors | recruiting | pituitary tumor | II | 21 * | clinical trial | treatment |
| checkpoint inhibition | Ipilimumab, Nivolumab | NCT02834013 | Nivolumab and Ipilimumab in Treating Patients With Rare Tumors | recruiting | pituitary carcinoma | II | 818 * | clinical trial | treatment |
| checkpoint inhibition | Pembrolizumab | NCT02721732 | Pembrolizumab in Treating Patients With Rare Tumors That Cannot Be Removed by Surgery or Are Metastatic | active, not recruiting | pituitary tumor | II | 202 | clinical trial | treatment |
| dopamine agonist treatment | Cabergoline | NCT03271918 | Cabergoline in Nonfunctioning Pituitary Adenomas (NFPA) | completed | NFPA | III | 140 | clinical trial | treatment |
| dopamine agonist treatment | Cabergoline | NCT02288962 | Dopamine Agonist Treatment of Non-functioning Pituitary Adenomas | recruiting | NFPA | III | 60 * | clinical trial | treatment |
| dopamine agonist treatment | Cabergoline | NCT00889525 | Study of Cabergoline in Treatment of Corticotroph Pituitary Adenoma | completed | Cushing’s disease | III | unknown | clinical trial | treatment |
| epidermal growth factor receptor (EGFR) inhibition | Lapatinib | NCT00939523 | Targeted Therapy With Lapatinib in Patients With Recurrent Pituitary Tumors Resistant to Standard Therapy | completed | pituitary tumor | II | 9 | clinical trial | treatment |
| epidermal growth factor receptor (EGFR) inhibition | Gefitinib | NCT02484755 | Targeted Therapy With Gefitinib in Patients With USP8 **-mutated Cushing’s Disease | unknown | Cushing’s disease | II | 6 * | clinical trial | treatment |
| interference with deoxyribonucleid acid (DNA) replication | Temozolomide, Radiotherapy | NCT04244708 | The Effect of Chemoradiotherapy in Patients With Refractory Pituitary Adenomas | not yet recruiting | pituitary tumor | II | 150 * | clinical trial | treatment |
| interference with deoxyribonucleid acid (DNA) replication | Temozolomide, Fluorouracil | NCT03930771 | Capecitabine and Temozolomide for Treatment of Recurrent Pituitary Adenomas | terminated | pituitary tumor | II | 1 | clinical trial | treatment |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Voellger, B.; Zhang, Z.; Benzel, J.; Wang, J.; Lei, T.; Nimsky, C.; Bartsch, J.-W. Targeting Aggressive Pituitary Adenomas at the Molecular Level—A Review. J. Clin. Med. 2022, 11, 124. https://doi.org/10.3390/jcm11010124
Voellger B, Zhang Z, Benzel J, Wang J, Lei T, Nimsky C, Bartsch J-W. Targeting Aggressive Pituitary Adenomas at the Molecular Level—A Review. Journal of Clinical Medicine. 2022; 11(1):124. https://doi.org/10.3390/jcm11010124
Chicago/Turabian StyleVoellger, Benjamin, Zhuo Zhang, Julia Benzel, Junwen Wang, Ting Lei, Christopher Nimsky, and Jörg-Walter Bartsch. 2022. "Targeting Aggressive Pituitary Adenomas at the Molecular Level—A Review" Journal of Clinical Medicine 11, no. 1: 124. https://doi.org/10.3390/jcm11010124
APA StyleVoellger, B., Zhang, Z., Benzel, J., Wang, J., Lei, T., Nimsky, C., & Bartsch, J. -W. (2022). Targeting Aggressive Pituitary Adenomas at the Molecular Level—A Review. Journal of Clinical Medicine, 11(1), 124. https://doi.org/10.3390/jcm11010124