
Recovering or Persisting: The Immunopathological Features of SARS-CoV-2 Infection in Children

 ,
,  , , , , ,
, , , , ,  ,
,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Population
- − Children with acute CoV2 infection (a microbiologically confirmed diagnosis through RT-PCR on nasopharyngeal swab by).
- − Children with PCC after microbiologically confirmed (with PCR on nasopharyngeal swab) acute COVID-19 were identified using an internationally developed survey (https://isaric.org/research/covid-19-clinical-research-resources/paediatric-follow-up (accessed on 4 July 2022)). Since there is not yet a consensus definition of post-acute sequelae of SARS-CoV-2 infection (PASC) in children, we defined as “PCC children” those having at least one persistent symptom for more than eight weeks after the diagnosis of acute COVID-19, according to recent studies in children and the NICE guidelines (https://www.nice.org.uk/guidance/ng188/resources/covid19-rapid-guideline-managing-the-longterm-effects-of-covid19-pdf-51035515742 (accessed on 19 July 2022)). Specifically, we chose the NICE guidelines because the persistence of symptoms for 8 weeks in children, who usually suffer from mild disease during acute infection, places an important burden on a child’s and a family’s quality of life, and patients seek medical attention before the 12 weeks cut-off.
- − Recovered children: those that reported no persistent symptoms after acute CoV2 infection and that were assessed at least 28 days from the onset of COVID-19 symptoms.
Inclusion and Exclusion Criteria
- Children aged 0–18 years.
- The child sought/needed primary or secondary medical care for COVID-19.
- Laboratory (RT-PCR) diagnosis of acute COVID-19.
- >28 days from the onset of COVID-19 symptoms.
- Parent’s/carer’s/guardian’s consent to participate.
2.2. Immunological Studies
2.2.1. Cytokines Analyses
2.2.2. Flow Cytometry Analysis
2.3. Ethic Committee Approval
2.4. Quantification and Statistical Analysis
3. Results
3.1. Study Population
3.2. Perturbation of Circulating B Cells and T Regulatory Subsets by CoV2 Infection in the Childhood
3.3. Heterogeneous Expression of Inflammatory Cytokines in CoV2+ Children
3.4. Age-Related Distribution of B Cells and Treg Subsets in CoV2+ Children
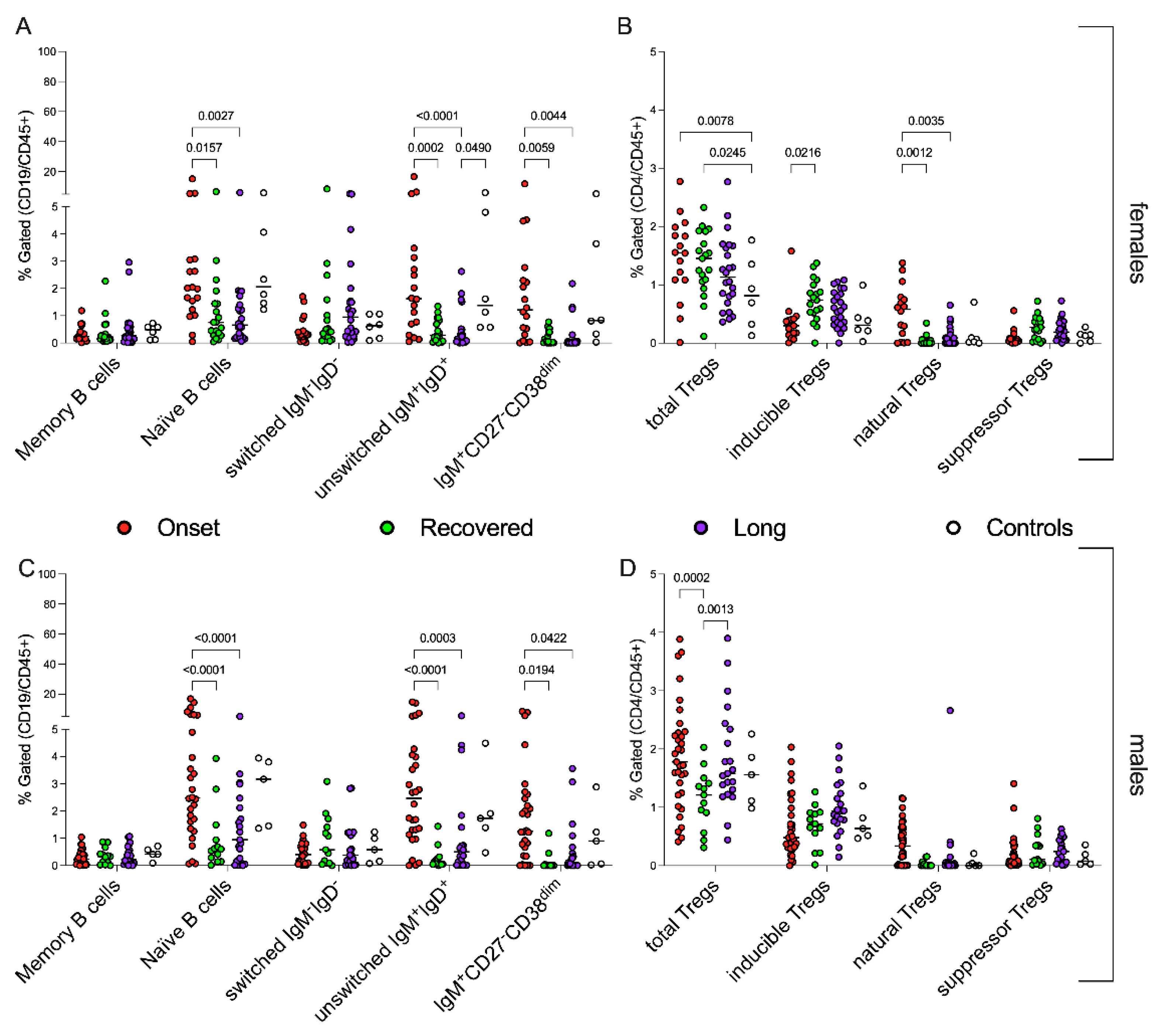
3.5. Sex-Related Modulation of Immune System in CoV2+ Children
3.6. Peripheral Blood Distribution of B Cells and Treg Subsets during Follow Up
4. Discussion
5. Limitations of the Study
6. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Parri, N.; Lenge, M.; Cantoni, B.; Arrighini, A.; Romanengo, M.; Urbino, A.; Da Dalt, L.; Verdoni, L.; Giacchero, R.; Lanari, M.; et al. COVID-19 in 17 Italian Pediatric Emergency Departments. Pediatrics 2020, 146, e20201235. [Google Scholar] [CrossRef] [PubMed]
- Götzinger, F.; Santiago-García, B.; Noguera-Julián, A.; Lanaspa, M.; Lancella, L.; Carducci, F.I.C.; Gabrovska, N.; Velizarova, S.; Prunk, P.; Osterman, V.; et al. COVID-19 in Children and Adolescents in Europe: A Multinational, Multicentre Cohort Study. Lancet Child Adolesc. Health 2020, 4, 653–661. [Google Scholar] [CrossRef]
- CDC COVID-19 Response Team. Severe Outcomes among Patients with Coronavirus Disease 2019 (COVID-19)-United States, 12 February–16 March 2020. MMWR Morb. Mortal. Wkly. Rep. 2020, 69, 343–346. [Google Scholar] [CrossRef] [PubMed]
- Dong, Y.; Mo, X.; Hu, Y.; Qi, X.; Jiang, F.; Jiang, Z.; Tong, S. Epidemiology of COVID-19 Among Children in China. Pediatrics 2020, 145, e20200702. [Google Scholar] [CrossRef] [Green Version]
- Feldstein, L.R.; Rose, E.B.; Horwitz, S.M.; Collins, J.P.; Newhams, M.M.; Son, M.B.F.; Newburger, J.W.; Kleinman, L.C.; Heidemann, S.M.; Martin, A.A.; et al. Multisystem Inflammatory Syndrome in U.S. Children and Adolescents. N. Engl. J. Med. 2020, 383, 334–346. [Google Scholar] [CrossRef]
- Onder, G.; Rezza, G.; Brusaferro, S. Case-Fatality Rate and Characteristics of Patients Dying in Relation to COVID-19 in Italy. JAMA 2020, 323, 1775–1776. [Google Scholar] [CrossRef]
- Wu, Z.; McGoogan, J.M. Characteristics of and Important Lessons from the Coronavirus Disease 2019 (COVID-19) Outbreak in China: Summary of a Report of 72,314 Cases from the Chinese Center for Disease Control and Prevention. JAMA 2020, 323, 1239–1242. [Google Scholar] [CrossRef]
- Bai, K.; Liu, W.; Liu, C.; Fu, Y.; Hu, J.; Qin, Y.; Zhang, Q.; Chen, H.; Xu, F.; Li, C. Clinical Analysis of 25 COVID-19 Infections in Children. Pediatric Infect. Dis. J. 2020, 39, e100–e103. [Google Scholar] [CrossRef]
- Lu, X.; Zhang, L.; Du, H.; Zhang, J.; Li, Y.Y.; Qu, J.; Zhang, W.; Wang, Y.; Bao, S.; Li, Y.; et al. SARS-CoV-2 Infection in Children. N. Engl. J. Med. 2020, 382, 1663–1665. [Google Scholar] [CrossRef] [Green Version]
- Valentini, P.; Sodero, G.; Buonsenso, D. The Relationship between COVID-19 and Innate Immunity in Children: A Review. Children 2021, 8, 266. [Google Scholar] [CrossRef]
- CMMID COVID-19 Working Group; Davies, N.G.; Klepac, P.; Liu, Y.; Prem, K.; Jit, M.; Eggo, R.M. Age-Dependent Effects in the Transmission and Control of COVID-19 Epidemics. Nat. Med. 2020, 26, 1205–1211. [Google Scholar] [CrossRef]
- Butt, A.A.; Dargham, S.R.; Loka, S.; Shaik, R.M.; Chemaitelly, H.; Tang, P.; Hasan, M.R.; Coyle, P.V.; Yassine, H.M.; Al-Khatib, H.A.; et al. Coronavirus Disease 2019 Disease Severity in Children Infected With the Omicron Variant. Clin. Infect. Dis. 2022, ciac275. [Google Scholar] [CrossRef]
- Carfì, A.; Bernabei, R.; Landi, F. Gemelli Against COVID-19 Post-Acute Care Study Group Persistent Symptoms in Patients after Acute COVID-19. JAMA 2020, 324, 603–605. [Google Scholar] [CrossRef]
- Stephenson, T.; Allin, B.; Nugawela, M.D.; Rojas, N.; Dalrymple, E.; Pereira, S.P.; Soni, M.; Knight, M.; Cheung, E.Y.; Heyman, I.; et al. Long COVID (Post-COVID-19 Condition) in Children: A Modified Delphi Process. Arch. Dis. Child. 2022, 107, 674–680. [Google Scholar] [CrossRef]
- Buonsenso, D.; Gennaro, L.D.; Rose, C.D.; Morello, R.; D’Ilario, F.; Zampino, G.; Piazza, M.; Boner, A.L.; Iraci, C.; O’Connell, S.; et al. Long-Term Outcomes of Pediatric Infections: From Traditional Infectious Diseases to Long Covid. Future Microbiol. 2022, 17, 551–571. [Google Scholar] [CrossRef]
- Buonsenso, D.; Munblit, D.; De Rose, C.; Sinatti, D.; Ricchiuto, A.; Carfi, A.; Valentini, P. Preliminary Evidence on Long COVID in Children. Acta Paediatr. 2021, 110, 2208–2211. [Google Scholar] [CrossRef]
- Ludvigsson, J.F. Case Report and Systematic Review Suggest That Children May Experience Similar Long-term Effects to Adults after Clinical COVID-19. Acta Paediatr. 2021, 110, 914–921. [Google Scholar] [CrossRef]
- Osmanov, I.M.; Spiridonova, E.; Bobkova, P.; Gamirova, A.; Shikhaleva, A.; Andreeva, M.; Blyuss, O.; El-Taravi, Y.; DunnGalvin, A.; Comberiati, P.; et al. Risk Factors for Post-COVID-19 Condition in Previously Hospitalised Children Using the ISARIC Global Follow-up Protocol: A Prospective Cohort Study. Eur. Respir. J. 2022, 59, 2101341. [Google Scholar] [CrossRef]
- Buonsenso, D.; Pujol, F.E.; Munblit, D.; Pata, D.; McFarland, S.; Simpson, F.K. Clinical characteristics, activity levels and mental health problems in children with long coronavirus disease: A survey of 510 children. Future Microbiol. 2022, 17, 577–588. [Google Scholar] [CrossRef]
- Buonsenso, D.; Di Giuda, D.; Sigfrid, L.; Pizzuto, D.A.; Di Sante, G.; De Rose, C.; Lazzareschi, I.; Sali, M.; Baldi, F.; Chieffo, D.P.R.; et al. Evidence of Lung Perfusion Defects and Ongoing Inflammation in an Adolescent with Post-Acute Sequelae of SARS-CoV-2 Infection. Lancet Child Adolesc. Health 2021, 5, 677–680. [Google Scholar] [CrossRef]
- Di Sante, G.; Buonsenso, D.; De Rose, C.; Tredicine, M.; Palucci, I.; De Maio, F.; Camponeschi, C.; Bonadia, N.; Biasucci, D.; Pata, D.; et al. Immunopathology of SARS-CoV-2 Infection: A Focus on T Regulatory and B Cell Responses in Children Compared with Adults. Children 2022, 9, 681. [Google Scholar] [CrossRef]
- Buonsenso, D.; Di Sante, G.; Sali, M. CURE COVID-19 Study Group Cytokine Profile in an Adolescent With Pediatric Multisystem Inflammatory Syndrome Temporally Related to COVID-19. Pediatr. Infect. Dis. J. 2020, 39, e213–e215. [Google Scholar] [CrossRef]
- Nalbandian, A.; Sehgal, K.; Gupta, A.; Madhavan, M.V.; McGroder, C.; Stevens, J.S.; Cook, J.R.; Nordvig, A.S.; Shalev, D.; Sehrawat, T.S.; et al. Post-Acute COVID-19 Syndrome. Nat. Med. 2021, 27, 601–615. [Google Scholar] [CrossRef]
- Couzin-Frankel, J. Clues to Long COVID. Science 2022, 376, 1261–1265. [Google Scholar] [CrossRef]
- Diorio, C.; McNerney, K.O.; Lambert, M.; Paessler, M.; Anderson, E.M.; Henrickson, S.E.; Chase, J.; Liebling, E.J.; Burudpakdee, C.; Lee, J.H.; et al. Evidence of Thrombotic Microangiopathy in Children with SARS-CoV-2 across the Spectrum of Clinical Presentations. Blood Adv. 2020, 4, 6051–6063. [Google Scholar] [CrossRef]
- Sinha, P.; Matthay, M.A.; Calfee, C.S. Is a “Cytokine Storm” Relevant to COVID-19? JAMA Intern. Med. 2020, 180, 1152–1154. [Google Scholar] [CrossRef]
- Mallapaty, S. Kids and COVID: Why Young Immune Systems Are Still on Top. Nature 2021, 597, 166–168. [Google Scholar] [CrossRef]
- Moderbacher, C.R.; Ramirez, S.I.; Dan, J.M.; Grifoni, A.; Hastie, K.M.; Weiskopf, D.; Belanger, S.; Abbott, R.K.; Kim, C.; Choi, J.; et al. Antigen-Specific Adaptive Immunity to SARS-CoV-2 in Acute COVID-19 and Associations with Age and Disease Severity. Cell 2020, 183, 996–1012. [Google Scholar] [CrossRef]
- Meckiff, B.J.; Ramírez-Suástegui, C.; Fajardo, V.; Chee, S.J.; Kusnadi, A.; Simon, H.; Eschweiler, S.; Grifoni, A.; Pelosi, E.; Weiskopf, D.; et al. Imbalance of Regulatory and Cytotoxic SARS-CoV-2-Reactive CD4+ T Cells in COVID-19. Cell 2020, 183, 1340–1353.e16. [Google Scholar] [CrossRef]
- Grifoni, A.; Tian, Y.; Sette, A.; Weiskopf, D. Transcriptomic Immune Profiles of Human Flavivirus-specific T-cell Responses. Immunology 2020, 160, 3–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Consiglio, C.R.; Cotugno, N.; Sardh, F.; Pou, C.; Amodio, D.; Rodriguez, L.; Tan, Z.; Zicari, S.; Ruggiero, A.; Pascucci, G.R.; et al. The Immunology of Multisystem Inflammatory Syndrome in Children with COVID-19. Cell 2020, 183, 968–981.e7. [Google Scholar] [CrossRef] [PubMed]
- Alivernini, S.; Tolusso, B.; Petricca, L.; Bui, L.; Di Sante, G.; Peluso, G.; Benvenuto, R.; Fedele, A.L.; Federico, F.; Ferraccioli, G.; et al. Synovial Features of Patients with Rheumatoid Arthritis and Psoriatic Arthritis in Clinical and Ultrasound Remission Differ under Anti-TNF Therapy: A Clue to Interpret Different Chances of Relapse after Clinical Remission? Ann. Rheum. Dis. 2017, 76, 1228–1236. [Google Scholar] [CrossRef] [PubMed]
- Ding, Y.; Zhou, L.; Xia, Y.; Wang, W.; Wang, Y.; Li, L.; Qi, Z.; Zhong, L.; Sun, J.; Tang, W.; et al. Reference Values for Peripheral Blood Lymphocyte Subsets of Healthy Children in China. J. Allergy Clin. Immunol. 2018, 142, 970–973.e8. [Google Scholar] [CrossRef] [PubMed]
- Schatorjé, E.J.H.; Gemen, E.F.A.; Driessen, G.J.A.; Leuvenink, J.; van Hout, R.W.N.M.; de Vries, E. Paediatric Reference Values for the Peripheral T Cell Compartment: T Lymphocyte Subsets in Children. Scand. J. Immunol. 2012, 75, 436–444. [Google Scholar] [CrossRef]
- Weisberg, S.P.; Connors, T.J.; Zhu, Y.; Baldwin, M.R.; Lin, W.-H.; Wontakal, S.; Szabo, P.A.; Wells, S.B.; Dogra, P.; Gray, J.; et al. Distinct Antibody Responses to SARS-CoV-2 in Children and Adults across the COVID-19 Clinical Spectrum. Nat. Immunol. 2021, 22, 25–31. [Google Scholar] [CrossRef]
- Tosif, S.; Neeland, M.R.; Sutton, P.; Licciardi, P.V.; Sarkar, S.; Selva, K.J.; Do, L.A.H.; Donato, C.; Toh, Z.Q.; Higgins, R.; et al. Immune Responses to SARS-CoV-2 in Three Children of Parents with Symptomatic COVID-19. Nat. Commun. 2020, 11, 5703. [Google Scholar] [CrossRef]
- Cohen, C.A.; Li, A.P.Y.; Hachim, A.; Hui, D.S.C.; Kwan, M.Y.W.; Tsang, O.T.Y.; Chiu, S.S.; Chan, W.H.; Yau, Y.S.; Kavian, N.; et al. SARS-CoV-2 Specific T Cell Responses Are Lower in Children and Increase with Age and Time after Infection. Nat. Commun. 2021, 12, 4678. [Google Scholar] [CrossRef]
- Bartsch, Y.C.; Wang, C.; Zohar, T.; Fischinger, S.; Atyeo, C.; Burke, J.S.; Kang, J.; Edlow, A.G.; Fasano, A.; Baden, L.R.; et al. Humoral Signatures of Protective and Pathological SARS-CoV-2 Infection in Children. Nat. Med. 2021, 27, 454–462. [Google Scholar] [CrossRef]
- Zuo, J.; Dowell, A.C.; Pearce, H.; Verma, K.; Long, H.M.; Begum, J.; Aiano, F.; Amin-Chowdhury, Z.; Hoschler, K.; Brooks, T.; et al. Robust SARS-CoV-2-Specific T Cell Immunity Is Maintained at 6 Months Following Primary Infection. Nat. Immunol. 2021, 22, 620–626. [Google Scholar] [CrossRef]
- Merad, M.; Blish, C.A.; Sallusto, F.; Iwasaki, A. The Immunology and Immunopathology of COVID-19. Science 2022, 375, 1122–1127. [Google Scholar] [CrossRef]
- Phetsouphanh, C.; Darley, D.R.; Wilson, D.B.; Howe, A.; Munier, C.M.L.; Patel, S.K.; Juno, J.A.; Burrell, L.M.; Kent, S.J.; Dore, G.J.; et al. Immunological Dysfunction Persists for 8 Months Following Initial Mild-to-Moderate SARS-CoV-2 Infection. Nat. Immunol. 2022, 23, 210–216. [Google Scholar] [CrossRef]
- Buonsenso, D.; Munblit, D.; Pazukhina, E.; Ricchiuto, A.; Sinatti, D.; Zona, M.; De Matteis, A.; D’Ilario, F.; Gentili, C.; Lanni, R.; et al. Post-COVID Condition in Adults and Children Living in the Same Household in Italy: A Prospective Cohort Study Using the ISARIC Global Follow-Up Protocol. Front. Pediatr. 2022, 10, 834875. [Google Scholar] [CrossRef]
- Townsend, L.; Dyer, A.H.; Naughton, A.; Kiersey, R.; Holden, D.; Gardiner, M.; Dowds, J.; O’Brien, K.; Bannan, C.; Nadarajan, P.; et al. Longitudinal Analysis of COVID-19 Patients Shows Age-Associated T Cell Changes Independent of Ongoing Ill-Health. Front. Immunol. 2021, 12, 676932. [Google Scholar] [CrossRef]
- Ryan, F.J.; Hope, C.M.; Masavuli, M.G.; Lynn, M.A.; Mekonnen, Z.A.; Yeow, A.E.L.; Garcia-Valtanen, P.; Al-Delfi, Z.; Gummow, J.; Ferguson, C.; et al. Long-Term Perturbation of the Peripheral Immune System Months after SARS-CoV-2 Infection. BMC Med. 2022, 20, 26. [Google Scholar] [CrossRef]
- Kusnadi, A.; Ramírez-Suástegui, C.; Fajardo, V.; Chee, S.J.; Meckiff, B.J.; Simon, H.; Pelosi, E.; Seumois, G.; Ay, F.; Vijayanand, P.; et al. Severely Ill Patients with COVID-19 Display Impaired Exhaustion Features in SARS-CoV-2–Reactive CD8+ T Cells. Sci. Immunol. 2021, 6, eabe4782. [Google Scholar] [CrossRef]
- Files, J.K.; Boppana, S.; Perez, M.D.; Sarkar, S.; Lowman, K.E.; Qin, K.; Sterrett, S.; Carlin, E.; Bansal, A.; Sabbaj, S.; et al. Sustained Cellular Immune Dysregulation in Individuals Recovering from SARS-CoV-2 Infection. J. Clin. Investig. 2021, 131, e140491. [Google Scholar] [CrossRef]
- Pušnik, J.; Richter, E.; Schulte, B.; Dolscheid-Pommerich, R.; Bode, C.; Putensen, C.; Hartmann, G.; Alter, G.; Streeck, H. Memory B Cells Targeting SARS-CoV-2 Spike Protein and Their Dependence on CD4+ T Cell Help. Cell Rep. 2021, 35, 109320. [Google Scholar] [CrossRef]
- Shuwa, H.A.; Shaw, T.N.; Knight, S.B.; Wemyss, K.; McClure, F.A.; Pearmain, L.; Prise, I.; Jagger, C.; Morgan, D.J.; Khan, S.; et al. Alterations in T and B Cell Function Persist in Convalescent COVID-19 Patients. Med 2021, 2, 720–735.e4. [Google Scholar] [CrossRef]
- Boothby, I.C.; Cohen, J.N.; Rosenblum, M.D. Regulatory T Cells in Skin Injury: At the Crossroads of Tolerance and Tissue Repair. Sci. Immunol. 2020, 5, eaaz9631. [Google Scholar] [CrossRef]
- Linterman, M.A.; Pierson, W.; Lee, S.K.; Kallies, A.; Kawamoto, S.; Rayner, T.F.; Srivastava, M.; Divekar, D.P.; Beaton, L.; Hogan, J.J.; et al. Foxp3+ Follicular Regulatory T Cells Control the Germinal Center Response. Nat. Med. 2011, 17, 975–982. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lund, J.M.; Hsing, L.; Pham, T.T.; Rudensky, A.Y. Coordination of Early Protective Immunity to Viral Infection by Regulatory T Cells. Science 2008, 320, 1220–1224. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Conway, S.R.; Lazarski, C.A.; Field, N.E.; Jensen-Wachspress, M.; Lang, H.; Kankate, V.; Durkee-Shock, J.; Kinoshita, H.; Suslovic, W.; Webber, K.; et al. SARS-CoV-2-Specific T Cell Responses Are Stronger in Children With Multisystem Inflammatory Syndrome Compared to Children With Uncomplicated SARS-CoV-2 Infection. Front. Immunol. 2022, 12, 793197. [Google Scholar] [CrossRef]
- Moss, P. The T Cell Immune Response against SARS-CoV-2. Nat. Immunol. 2022, 23, 186–193. [Google Scholar] [CrossRef] [PubMed]
- Kudlay, D.; Kofiadi, I.; Khaitov, M. Peculiarities of the T Cell Immune Response in COVID-19. Vaccines 2022, 10, 242. [Google Scholar] [CrossRef]
- Pandolfi, F.; Cianci, R.; Casciano, F.; Pagliari, D.; De Pasquale, T.; Landolfi, R.; Di Sante, G.; Kurnick, J.T.; Ria, F. Skewed T-Cell Receptor Repertoire: More than a Marker of Malignancy, a Tool to Dissect the Immunopathology of Inflammatory Diseases. J. Biol. Regul. Homeost. Agents 2011, 25, 153–161. [Google Scholar]
- Yonker, L.M.; Gilboa, T.; Ogata, A.F.; Senussi, Y.; Lazarovits, R.; Boribong, B.P.; Bartsch, Y.C.; Loiselle, M.; Rivas, M.N.; Porritt, R.A.; et al. Multisystem Inflammatory Syndrome in Children Is Driven by Zonulin-Dependent Loss of Gut Mucosal Barrier. J. Clin. Investig. 2021, 131, 149633. [Google Scholar] [CrossRef]
- Stephenson, T.; Pereira, S.M.P.; Shafran, R.; de Stavola, B.L.; Rojas, N.; McOwat, K.; Simmons, R.; Zavala, M.; O’Mahoney, L.; Chalder, T.; et al. Physical and Mental Health 3 Months after SARS-CoV-2 Infection (Long COVID) among Adolescents in England (CLoCk): A National Matched Cohort Study. Lancet Child Adolesc. Health 2022, 6, 230–239. [Google Scholar] [CrossRef]
- Liu, J.; Yang, X.; Wang, H.; Li, Z.; Deng, H.; Liu, J.; Xiong, S.; He, J.; Feng, X.; Guo, C.; et al. Analysis of the Long-Term Impact on Cellular Immunity in COVID-19-Recovered Individuals Reveals a Profound NKT Cell Impairment. mBio 2021, 12, e00085-21. [Google Scholar] [CrossRef]
- Siska, P.J.; Decking, S.-M.; Babl, N.; Matos, C.; Bruss, C.; Singer, K.; Klitzke, J.; Schön, M.; Simeth, J.; Köstler, J.; et al. Metabolic Imbalance of T Cells in COVID-19 Is Hallmarked by Basigin and Mitigated by Dexamethasone. J. Clin. Investig. 2021, 131, e148225. [Google Scholar] [CrossRef]
- Su, Y.; Yuan, D.; Chen, D.G.; Ng, R.H.; Wang, K.; Choi, J.; Li, S.; Hong, S.; Zhang, R.; Xie, J.; et al. Multiple Early Factors Anticipate Post-Acute COVID-19 Sequelae. Cell 2022, 185, 881–895.e20. [Google Scholar] [CrossRef]
- Gold, J.E.; Okyay, R.A.; Licht, W.E.; Hurley, D.J. Investigation of Long COVID Prevalence and Its Relationship to Epstein-Barr Virus Reactivation. Pathogens 2021, 10, 763. [Google Scholar] [CrossRef]
- Pieniawska-Śmiech, K.; Kuraszewicz, A.; Sado, J.; Śmiech, K.; Lewandowicz-Uszyńska, A. Assessment of COVID-19 Incidence and the Ability to Synthesise Anti-SARS-CoV-2 Antibodies of Paediatric Patients with Primary Immunodeficiency. J. Clin. Med. 2021, 10, 5111. [Google Scholar] [CrossRef]
- Morrocchi, E.; Pighi, C.; Pascucci, G.R.; Cotugno, N.; Medri, C.; Amodio, D.; Colagrossi, L.; Ruggiero, A.; Manno, E.C.; Tumeo, C.C.; et al. Perinatally HIV-Infected Adolescents and Young Adults Demonstrate Distinct BNT162b2 MRNA COVID-19 Vaccine Immunogenicity. Clin. Infect. Dis. 2022, ciac408. [Google Scholar] [CrossRef]
- Glynne, P.; Tahmasebi, N.; Gant, V.; Gupta, R. Long COVID Following Mild SARS-CoV-2 Infection: Characteristic T Cell Alterations and Response to Antihistamines. J. Investig. Med. 2022, 70, 61–67. [Google Scholar] [CrossRef]
- Zimmermann, P.; Pittet, L.F.; Curtis, N. How Common Is Long COVID in Children and Adolescents? Pediatric Infect. Dis. J. 2021, 40, e482–e487. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Long CoV2+ | CoV2+ Onset | CoV2+ Recovered | CoV2− Controls | ||
|---|---|---|---|---|---|
| n = 51 | n = 49 | n = 32 | n = 9 | ||
| Age | 10.6 ± 4.62 | 7.1 ± 5.6 | 7.75 ± 5.5 | 7.3 ± 5.9 | |
| Females | 26 (51) | 17 (34.7) | 20 (62.5) | 3 (33.33) | |
| Distance from acute infection (months) | 4.2 ± 2.16 | 0 | 4.2 ± 2.04 | 0 | |
| Severity of Acute Disease | Asymptomatic | 4 (7.8) | 6 (12.2%) | 4 (12.5) | / |
| Mild | 42 (82.4) | 27 (55.1) | 26 (81.25) | ||
| Moderate | 5 (9.8) | 10 (20.5) | 0 | ||
| Severe | 0 | 6 (12.2) | 2 (6.25) | ||
| Persistent symptoms in Long COVID children | Headache | 19 (37.2) | / | / | / |
| Dyspnea after efforts | 15 (29.4) | ||||
| Muscle-skeletal pain | 12 (23.5) | ||||
| Gastrointestinal issues | 11 (21.5) | ||||
| Fatigue | 10 (19.6) | ||||
| Chest pain | 10 (19.6) | ||||
| Tachycardia | 6 (11.7) | ||||
| Anosmia | 5 (9.8) | ||||
| Dysgeusia | 5 (9.8) | ||||
| Skin rashes | 5 (9.8) | ||||
| Low grade fever | 4 (7.8) | ||||
| Sleeping problems | 4 (7.8) | ||||
| Lymphocyte Populations | Onset | Recovered | Long | p-Values | ||
|---|---|---|---|---|---|---|
| a | b | c | ||||
| IgD−CD27+ memory B cells | 0.3 ± 0.3 | 0.4 ± 0.4 | 0.4 ± 0.6 | 1.0 | 1.0 | 1.0 |
| IgD+CD27− naïve B cells | 3.9 ± 3.9 | 1.1 ± 1.4 | 1.2 ± 1.3 | <0.0001 | <0.0001 | 1.0 |
| IgD−IgM− B cells | 0.5 ± 0.4 | 1.0 ± 1.6 | 1.0 ± 1.3 | 0.6 | 0.5 | 1.0 |
| IgD+IgM+ B cells | 3.4 ± 6.4 | 0.4 ± 0.4 | 0.7 ± 1.2 | <0.0001 | <0.0001 | 1.0 |
| IgM+CD27−CD38dim B cells | 2.2 ± 2.5 | 0.2 ± 0.3 | 0.4 ± 0.8 | <0.0001 | <0.0001 | 0.8 |
| CD25highFOXP3+ Treg | 1.9 ± 0.8 | 1.3 ± 0.5 | 1.5 ± 0.8 | <0.0001 | 0.0004 | 0.14 |
| Inducible Tregs | 0.5 ± 0.5 | 0.7 ± 0.4 | 0.8 ± 0.5 | 0.5 | 0.1 | 0.9 |
| Natural Tregs | 0.4 ± 0.4 | 0.04 ± 0.07 | 0.1 ± 0.4 | 0.0005 | 0.0026 | 0.8 |
| Suppressor T regs | 0.2 ± 0.3 | 0.3 ± 0.2 | 0.2 ± 0.2 | 0.7 | 0.8 | 0.9 |
| Lymphocyte Populations | Onset | Recovered | Long | p-Values | ||
|---|---|---|---|---|---|---|
| a | b | c | ||||
| IgD−CD27+ memory B cells | 0.3 ± 0.3 | 0.3 ± 0.3 | 0.5 ± 0.6 | 1.0 | 1.0 | 1.0 |
| IgD+CD27− naïve B cells | 2.6 ± 1.9 | 0.9 ± 1.2 | 1.2 ± 1.2 | 0.0002 | <0.0001 | 0.9 |
| IgD−IgM− B cells | 0.4 ± 0.4 | 0.9 ± 0.9 | 0.9 ± 0.9 | 0.6 | 0.4 | 1.0 |
| IgD+IgM+ B cells | 2.2 ± 1.7 | 0.3 ± 0.4 | 1.0 ± 1.5 | <0.0001 | 0.001 | 0.2 |
| IgM+CD27−CD38dim B cells | 1.6 ± 1.3 | 0.2 ± 0.3 | 0.6 ± 0.9 | 0.002 | 0.02 | 0.7 |
| CD25highFOXP3+ total Treg | 1.5 ± 0.8 | 1.1 ± 0.6 | 1.4 ± 0.7 | 0.1 | 1.0 | 0.3 |
| Inducible Tregs | 0.5 ± 0.5 | 0.7 ± 0.4 | 0.8 ± 0.4 | 0.7 | 0.2 | 0.9 |
| Natural Tregs | 0.3 ± 0.4 | 0.03 ± 0.05 | 0.1 ± 0.2 | 0.2 | 0.3 | 1.0 |
| Suppressor T regs | 0.2 ± 0.3 | 0.3 ± 0.2 | 0.3 ± 0.2 | 0.9 | 0.9 | 1.0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Buonsenso, D.; Valentini, P.; De Rose, C.; Tredicine, M.; Pereyra Boza, M.d.C.; Camponeschi, C.; Morello, R.; Zampino, G.; Brooks, A.E.S.; Rende, M.; et al. Recovering or Persisting: The Immunopathological Features of SARS-CoV-2 Infection in Children. J. Clin. Med. 2022, 11, 4363. https://doi.org/10.3390/jcm11154363
Buonsenso D, Valentini P, De Rose C, Tredicine M, Pereyra Boza MdC, Camponeschi C, Morello R, Zampino G, Brooks AES, Rende M, et al. Recovering or Persisting: The Immunopathological Features of SARS-CoV-2 Infection in Children. Journal of Clinical Medicine. 2022; 11(15):4363. https://doi.org/10.3390/jcm11154363
Chicago/Turabian StyleBuonsenso, Danilo, Piero Valentini, Cristina De Rose, Maria Tredicine, Maria del Carmen Pereyra Boza, Chiara Camponeschi, Rosa Morello, Giuseppe Zampino, Anna E. S. Brooks, Mario Rende, and et al. 2022. "Recovering or Persisting: The Immunopathological Features of SARS-CoV-2 Infection in Children" Journal of Clinical Medicine 11, no. 15: 4363. https://doi.org/10.3390/jcm11154363
APA StyleBuonsenso, D., Valentini, P., De Rose, C., Tredicine, M., Pereyra Boza, M. d. C., Camponeschi, C., Morello, R., Zampino, G., Brooks, A. E. S., Rende, M., Ria, F., Sanguinetti, M., Delogu, G., Sali, M., Di Sante, G., & on behalf of the Gemelli-Pediatric COVID-19 Team. (2022). Recovering or Persisting: The Immunopathological Features of SARS-CoV-2 Infection in Children. Journal of Clinical Medicine, 11(15), 4363. https://doi.org/10.3390/jcm11154363