
Regulatory Roles of Estrogens in Psoriasis

Abstract
:1. Introduction
2. Physiology of Estrogens
2.1. Physiological Levels of Estrogens
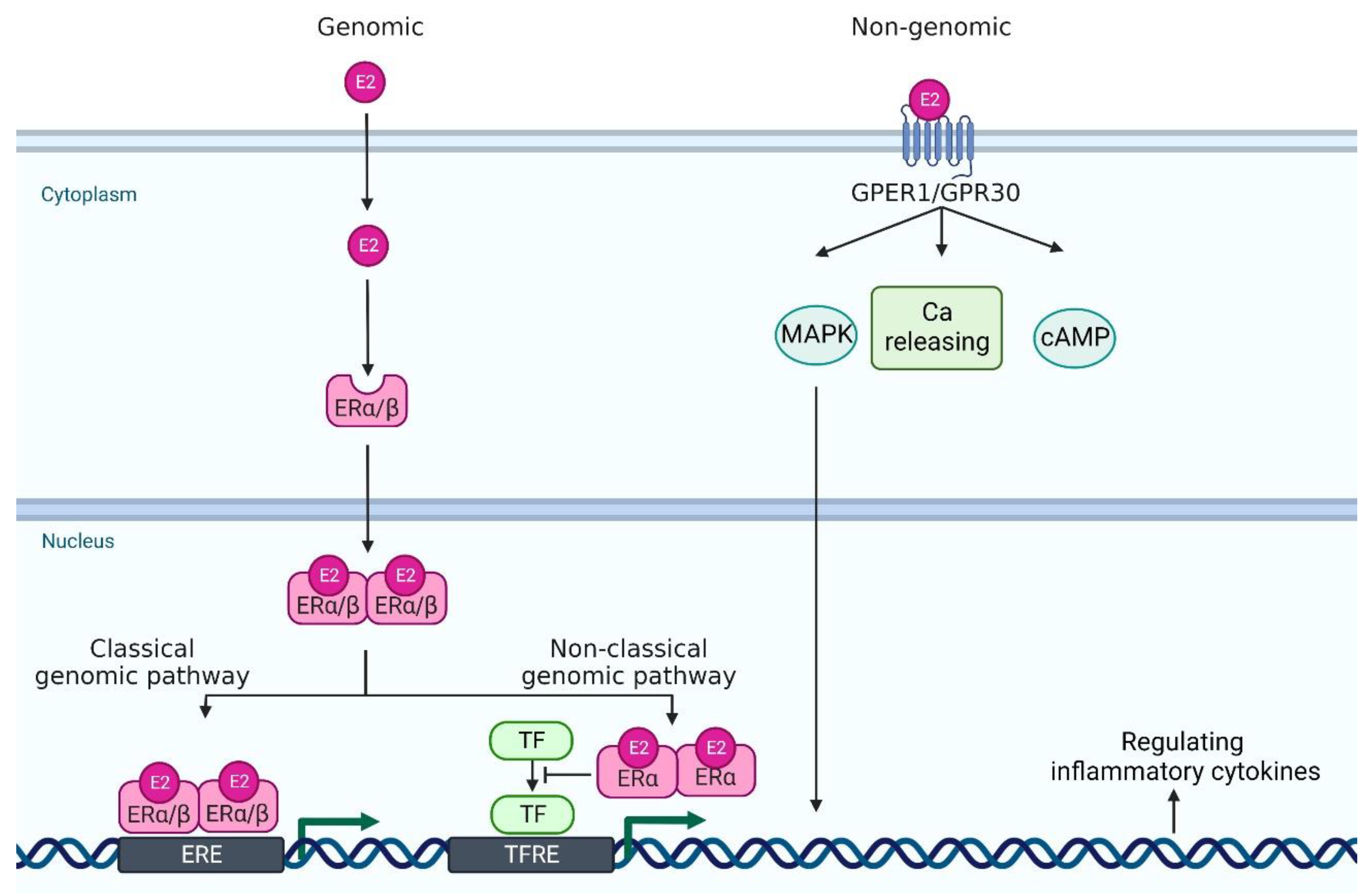
2.2. Estrogen Receptors and Their Signaling
3. Epidemiological and Case Series Studies about the Possible Involvement of Estrogens in Psoriasis
4. In Vitro Studies Regarding the Immuno-Regulatory Action of E2
4.1. Keratinocytes
4.2. Neutrophils, Monocytes, and Macrophages
4.3. DCs
4.4. T cells
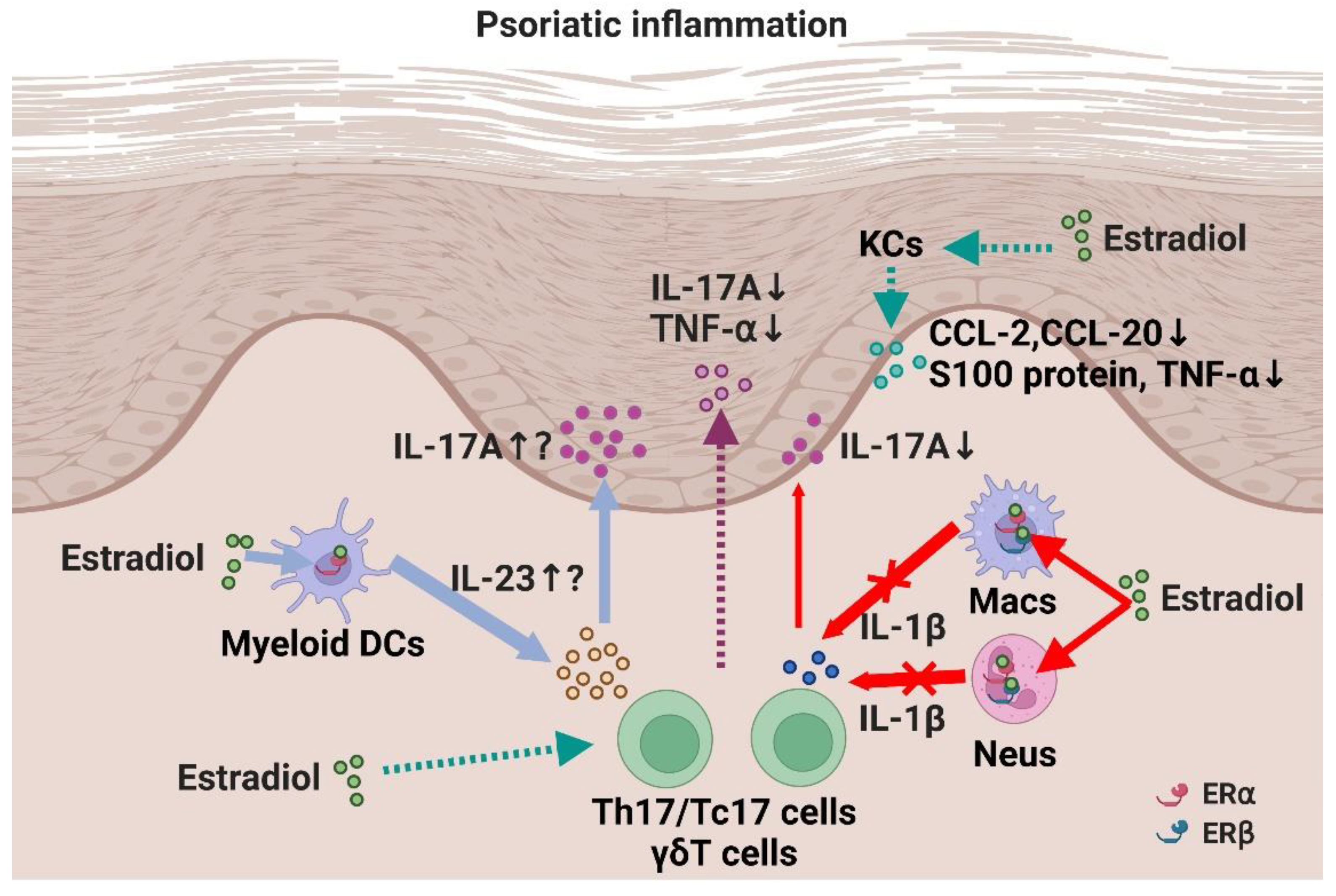
5. In Vivo Studies Regarding the Role of E2 on Psoriatic Inflammation
6. Concluding Remarks
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Nestle, F.O.; Kaplan, D.H.; Barker, J. Psoriasis. N. Engl. J. Med. 2009, 361, 496–509. [Google Scholar] [CrossRef] [PubMed]
- Parisi, R.; Symmons, D.P.M.; Griffiths, C.E.M.; Ashcroft, D.M. Global epidemiology of psoriasis: A systematic review of incidence and prevalence. J. Investig. Dermatol. 2013, 133, 377–385. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McGeachy, M.J.; Cua, D.J.; Gaffen, S.L. The IL-17 Family of Cytokines in Health and Disease. Immunity 2019, 50, 892–906. [Google Scholar] [CrossRef]
- Cai, Y.; Shen, X.; Ding, C.; Qi, C.; Li, K.; Li, X.; Jala, V.R.; Zhang, H.-g.; Wang, T.; Zheng, J.; et al. Pivotal Role of Dermal IL-17-Producing γδ T Cells in Skin Inflammation. Immunity 2011, 35, 596–610. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cai, Y.; Xue, F.; Quan, C.; Qu, M.; Liu, N.; Zhang, Y.; Fleming, C.; Hu, X.; Zhang, H.; Weichselbaum, R.; et al. A Critical Role of the IL-1β–IL-1R Signaling Pathway in Skin Inflammation and Psoriasis Pathogenesis. J. Investig. Dermatol. 2019, 139, 146–156. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Greb, J.E.; Goldminz, A.M.; Elder, J.T.; Lebwohl, M.G.; Gladman, D.D.; Wu, J.J.; Mehta, N.N.; Finlay, A.Y.; Gottlieb, A.B. Psoriasis. Nat. Rev. Dis. Prim. 2016, 2, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Griffiths, C.E.M.; Armstrong, A.W.; Gudjonsson, J.E.; Barker, J.N.W.N. Psoriasis. Lancet 2021, 397, 1301–1315. [Google Scholar] [CrossRef]
- Kanda, N.; Watanabe, S. Regulatory roles of sex hormones in cutaneous biology and immunology. J. Dermatol. Sci. 2005, 38, 1–7. [Google Scholar] [CrossRef]
- Klein, S.L.; Flanagan, K.L. Sex differences in immune responses. Nat. Rev. Immunol. 2016, 16, 626–638. [Google Scholar] [CrossRef]
- Hall, O.J.; Klein, S.L. Progesterone-based compounds affect immune responses and susceptibility to infections at diverse mucosal sites. Mucosal Immunol. 2017, 10, 1097–1107. [Google Scholar] [CrossRef] [Green Version]
- Straub, R.H. The complex role of estrogens in inflammation. Endocr. Rev. 2007, 28, 521–574. [Google Scholar] [CrossRef] [Green Version]
- Stanczyk, F.Z.; Clarke, N.J. Measurement of estradiol-challenges ahead. J. Clin. Endocrinol. Metab. 2014, 99, 56–58. [Google Scholar] [CrossRef] [PubMed]
- Kovats, S. Estrogen receptors regulate innate immune cells and signaling pathways. Cell. Immunol. 2015, 294, 63–69. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ghisletti, S.; Meda, C.; Maggi, A.; Vegeto, E. 17β-Estradiol Inhibits Inflammatory Gene Expression by Controlling NF-κB Intracellular Localization. Mol. Cell. Biol. 2005, 25, 2957–2968. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Björnström, L.; Sjöberg, M. Mechanisms of estrogen receptor signaling: Convergence of genomic and nongenomic actions on target genes. Mol. Endocrinol. 2005, 19, 833–842. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chadjichristos, C.; Ghayor, C.; Kypriotou, M.; Martin, G.; Renard, E.; Ala-Kokko, L.; Suske, G.; De Crombrugghe, B.; Pujol, J.P.; Galéra, P. Sp1 and Sp3 transcription factors mediate interleukin-1β down-regulation of human type II collagen gene expression in articular chondrocytes. J. Biol. Chem. 2003, 278, 39762–39772. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Novoszel, P.; Holcmann, M.; Stulnig, G.; De Sa Fernandes, C.; Zyulina, V.; Borek, I.; Linder, M.; Bogusch, A.; Drobits, B.; Bauer, T.; et al. Psoriatic skin inflammation is promoted by c-Jun/AP-1-dependent CCL2 and IL-23 expression in dendritic cells. EMBO Mol. Med. 2021, 13, 1–18. [Google Scholar] [CrossRef]
- Wang, S.; Wang, W.; Wesley, R.A.; Danner, R.L. A Sp1 binding site of the tumor necrosis factor α promoter functions as a nitric oxide response element. J. Biol. Chem. 1999, 274, 33190–33193. [Google Scholar] [CrossRef] [Green Version]
- Nerlich, A.; Ruangkiattikul, N.; Laarmann, K.; Janze, N.; Dittrich-Breiholz, O.; Kracht, M.; Goethe, R. C/EBPβ is a transcriptional key regulator of IL-36α in murine macrophages. Biochim. Biophys. Acta—Gene Regul. Mech. 2015, 1849, 966–978. [Google Scholar] [CrossRef]
- Liu, T.; Zhang, L.; Joo, D.; Sun, S.C. NF-κB signaling in inflammation. Signal Transduct. Target. Ther. 2017, 2, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Notas, G.; Kampa, M.; Castanas, E. G Protein-Coupled Estrogen Receptor in Immune Cells and Its Role in Immune-Related Diseases. Front. Endocrinol. 2020, 11, 579420. [Google Scholar] [CrossRef] [PubMed]
- Revankar, C.M.; Cimino, D.F.; Sklar, L.A.; Arterburn, J.B.; Prossnitz, E.R. A transmembrane intracellular estrogen receptor mediates rapid cell signaling. Science 2005, 307, 1625–1630. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hägg, D.; Sundström, A.; Eriksson, M.; Schmitt-Egenolf, M. Severity of Psoriasis Differs Between Men and Women: A Study of the Clinical Outcome Measure Psoriasis Area and Severity Index (PASI) in 5438 Swedish Register Patients. Am. J. Clin. Dermatol. 2017, 18, 583–590. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Furue, M.; Yamazaki, S.; Jimbow, K.; Tsuchida, T.; Amagai, M.; Tanaka, T.; Matsunaga, K.; Muto, M.; Morita, E.; Akiyama, M.; et al. Prevalence of dermatological disorders in Japan: A nationwide, cross-sectional, seasonal, multicenter, hospital-based study. J. Dermatol. 2011, 38, 310–320. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, H.; Takahashi, I.; Tsuji, H.; Ibe, M.; Kinouchi, M.; Hashimoto, Y.; Ishida-Yamamoto, A.; Matsuo, S.; Ohkuma, N.; Ohkawara, A.; et al. Analysis of psoriatic patients registered in Asahikawa Medical College Hospital from 1983 to 2007. J. Dermatol. 2009, 36, 632–637. [Google Scholar] [CrossRef]
- Kawada, A.; Tezuka, T.; Nakamizo, Y.; Kimura, H.; Nakagawa, H.; Ohkido, M.; Ozawa, A.; Ohkawara, A.; Kobayashi, H.; Harada, S.; et al. A survey of psoriasis patients in Japan from 1982 to 2001. J. Dermatol. Sci. 2003, 31, 59–64. [Google Scholar] [CrossRef]
- El-Komy, M.H.M.; Mashaly, H.; Sayed, K.S.; Hafez, V.; El-Mesidy, M.S.; Said, E.R.; Amer, M.A.; AlOrbani, A.M.; Saadi, D.G.; El-Kalioby, M.; et al. Clinical and epidemiologic features of psoriasis patients in an Egyptian medical center. JAAD Int. 2020, 1, 81–90. [Google Scholar] [CrossRef]
- Lee, J.Y.; Kang, S.; Park, J.S.; Jo, S.J. Prevalence of psoriasis in Korea: A population-based epidemiological study using the Korean national health insurance database. Ann. Dermatol. 2017, 29, 761–767. [Google Scholar] [CrossRef] [Green Version]
- Na, S.J.; Jo, S.J.; Youn, J. Il Clinical study on psoriasis patients for past 30 years (1982-2012) in Seoul National University Hospital Psoriasis Clinic. J. Dermatol. 2013, 40, 731–735. [Google Scholar] [CrossRef]
- Tsai, T.F.; Wang, T.S.; Hung, S.T.; Tsai, P.I.C.; Schenkel, B.; Zhang, M.; Tang, C.H. Epidemiology and comorbidities of psoriasis patients in a national database in Taiwan. J. Dermatol. Sci. 2011, 63, 40–46. [Google Scholar] [CrossRef]
- Bayaraa, B.; Imafuku, S. Relationship between environmental factors, age of onset and familial history in Japanese patients with psoriasis. J. Dermatol. 2018, 45, 715–718. [Google Scholar] [CrossRef] [PubMed]
- Farber, E.M.; Nall, M.L. The natural history of psoriasis in 5600 patients. Dermatologica 1974, 148, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Armstrong, A.W.; Mehta, M.D.; Schupp, C.W.; Gondo, G.C.; Bell, S.J.; Griffiths, C.E.M. Psoriasis Prevalence in Adults in the United States. JAMA Dermatol. 2021, 157, 940–946. [Google Scholar] [CrossRef] [PubMed]
- Henseler, T.; Christophers, E. Psoriasis of early and late onset: Characterization of two types of psoriasis vulgaris. J. Am. Acad. Dermatol. 1985, 13, 450–456. [Google Scholar] [CrossRef]
- Guillet, C.; Seeli, C.; Nina, M.; Maul, L.V.; Maul, J.-T. The impact of gender and sex in psoriasis: What to be aware of when treating women with psoriasis. Int. J. Women’s Dermatol. 2022, 8, e010. [Google Scholar] [CrossRef] [PubMed]
- Parisi, R.; Iskandar, I.Y.K.; Kontopantelis, E.; Augustin, M.; Griffiths, C.E.M.; Ashcroft, D.M. National, regional, and worldwide epidemiology of psoriasis: Systematic analysis and modelling study. BMJ 2020, 369, 369. [Google Scholar] [CrossRef] [PubMed]
- Murphy, F.R.; Stolman, L.P. Generalized Pustular Psoriasis. Arch. Dermatol. 1979, 115, 1215–1216. [Google Scholar] [CrossRef] [Green Version]
- Murase, J.E.; Chan, K.K.; Garite, T.J.; Cooper, D.M.; Weinstein, G.D. Hormonal effect on psoriasis in pregnancy and post partum. Arch. Dermatol. 2005, 141, 601–606. [Google Scholar] [CrossRef]
- Raychaudhuri, S.P.; Navare, T.; Gross, J.; Raychaudhuri, S.K. Clinical course of psoriasis during pregnancy. Int. J. Dermatol. 2003, 42, 518–520. [Google Scholar] [CrossRef] [PubMed]
- Mowad, C.M.; Margolis, D.J.; Halpern, A.C.; Suri, B.; Synnestvedt, M.; Guzzo, C.A. Hormonal Influences on Women with Psoriasis. Cutis 1998, 61, 257–260. [Google Scholar]
- Park, B.S.; Youn, J. Il Factors influencing psoriasis: An analysis based upon the extent of involvement and clinical type. J. Dermatol. 1998, 25, 97–102. [Google Scholar] [CrossRef] [PubMed]
- Boyd, A.S.; Morris, L.F.; Phillips, C.M.; Menter, M.A. Psoriasis and pregnancy: Hormone and immune system interaction. Int. J. Dermatol. 1996, 35, 169–172. [Google Scholar] [CrossRef] [PubMed]
- McHugh, N.J.; Laurent, M.R. The effect of pregnancy on the onset of psoriatic arthritis. Br. J. Rheumatol. 1989, 28, 50–52. [Google Scholar] [CrossRef] [PubMed]
- Cemil, B.C.; Cengiz, F.P.; Atas, H.; Ozturk, G.; Canpolat, F. Sex hormones in male psoriasis patients and their correlation with the Psoriasis Area and Severity Index. J. Dermatol. 2015, 42, 500–503. [Google Scholar] [CrossRef]
- Braverman, I.M.; Cohen, I.; O’keefe, E. Metabolic and Ultrastructural Studies in a Patient With Pustular Psoriasis (von Zumbusch). Arch. Dermatol. 1972, 105, 189–196. [Google Scholar] [CrossRef]
- Spangler, A.S.; Antoniades, H.N.; Sotman, S.L.; Inderbitizin, T.M. Enhancement of the anti-inflammatory action of hydrocortisone by estrogen. J. Clin. Endocrinol. Metab. 1969, 29, 650–655. [Google Scholar] [CrossRef]
- Boyd, A.S.; King, J. Tamoxifen-induced remission of psoriasis. J. Am. Acad. Dermatol. 1999, 41, 887–889. [Google Scholar] [CrossRef]
- Smolinska, E.; Moskot, M.; Jakóbkiewicz-Banecka, J.; Wegrzyn, G.; Banecki, B.; Szczerkowska-Dobosz, A.; Purzycka-Bohdan, D.; Gabig-Ciminska, M. Molecular action of isoflavone genistein in the human epithelial cell line HaCaT. PLoS ONE 2018, 13, e0192297. [Google Scholar] [CrossRef]
- Bocheńska, K.; Moskot, M.; Smolińska-Fijołek, E.; Jakóbkiewicz-Banecka, J.; Szczerkowska-Dobosz, A.; Słomiński, B.; Gabig-Cimińska, M. Impact of isoflavone genistein on psoriasis in in vivo and in vitro investigations. Sci. Rep. 2021, 11, 1–16. [Google Scholar] [CrossRef]
- Dainichi, T.; Kitoh, A.; Otsuka, A.; Nakajima, S.; Nomura, T.; Kaplan, D.H.; Kabashima, K. The epithelial immune microenvironment (EIME) in atopic dermatitis and psoriasis. Nat. Immunol. 2018, 19, 1286–1298. [Google Scholar] [CrossRef]
- Kanda, N.; Watanabe, S. 17β-Estradiol Inhibits MCP-1 Production in Human Keratinocytes. J. Investig. Dermatol. 2003, 120, 1058–1066. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kanda, N.; Watanabe, S. 17β-estradiol inhibits the production of RANTES in human keratinocytes. J. Investig. Dermatol. 2003, 120, 420–427. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, H.; Yao, Q.; Mariscal, A.G.; Wu, X.; Hülse, J.; Pedersen, E.; Helin, K.; Waisman, A.; Vinkel, C.; Thomsen, S.F.; et al. Epigenetic control of IL-23 expression in keratinocytes is important for chronic skin inflammation. Nat. Commun. 2018, 9, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Wang, A.; Wei, J.; Lu, C.; Chen, H.; Zhong, X.; Lu, Y.; Li, L.; Huang, H.; Dai, Z.; Han, L. Genistein suppresses psoriasis-related inflammation through a STAT3–NF-κB-dependent mechanism in keratinocytes. Int. Immunopharmacol. 2019, 69, 270–278. [Google Scholar] [CrossRef]
- Toichi, E.; Tachibana, T.; Furukawa, F. Rapid improvement of psoriasis vulgaris during drug-induced agranulocytosis. J. Am. Acad. Dermatol. 2000, 43, 391–395. [Google Scholar] [CrossRef]
- Ikeda, S.; Takahashi, H.; Suga, Y.; Eto, H.; Etoh, T.; Okuma, K.; Takahashi, K.; Kanbara, T.; Seishima, M.; Morita, A.; et al. Therapeutic depletion of myeloid lineage leukocytes in patients with generalized pustular psoriasis indicates a major role for neutrophils in the immunopathogenesis of psoriasis. J. Am. Acad. Dermatol. 2013, 68, 609–617. [Google Scholar] [CrossRef]
- Sumida, H.; Yanagida, K.; Kita, Y.; Abe, J.; Matsushima, K.; Nakamura, M.; Ishii, S.; Sato, S.; Shimizu, T. Interplay between CXCR2 and BLT1 Facilitates Neutrophil Infiltration and Resultant Keratinocyte Activation in a Murine Model of Imiquimod-Induced Psoriasis. J. Immunol. 2014, 192, 4361–4369. [Google Scholar] [CrossRef]
- Han, G.; Havnaer, A.; Lee, H.H.; Carmichael, D.J.; Martinez, L.R. Biological depletion of neutrophils attenuates pro-inflammatory markers and the development of the psoriatic phenotype in a murine model of psoriasis. Clin. Immunol. 2020, 210, 108294. [Google Scholar] [CrossRef]
- Morimura, S.; Oka, T.; Sugaya, M.; Sato, S. CX3CR1 deficiency attenuates imiquimod-induced psoriasis-like skin inflammation with decreased M1 macrophages. J. Dermatol. Sci. 2016, 82, 175–188. [Google Scholar] [CrossRef]
- Matsumoto, R.; Sugimoto, Y.; Kabashima, K.; Matsumoto, R.; Dainichi, T.; Tsuchiya, S.; Nomura, T.; Kitoh, A. Epithelial TRAF6 drives IL-17–mediated psoriatic inflammation. JCI Insight 2018, 3, 121175. [Google Scholar] [CrossRef]
- Ito, A.; Bebo, B.F.; Matejuk, A.; Zamora, A.; Silverman, M.; Fyfe-Johnson, A.; Offner, H. Estrogen Treatment Down-Regulates TNF-α Production and Reduces the Severity of Experimental Autoimmune Encephalomyelitis in Cytokine Knockout Mice. J. Immunol. 2001, 167, 542–552. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carruba, G.; D’Agostino, P.; Miele, M.; Calabrò, M.; Barbera, C.; Di Bella, G.; Milano, S.; Ferlazzo, V.; Caruso, R.; La Rosa, M.; et al. Estrogen regulates cytokine production and apoptosis in PMA-differentiated, macrophage-like U937 cells. J. Cell. Biochem. 2003, 90, 187–196. [Google Scholar] [CrossRef] [PubMed]
- Bouman, A.; Schipper, M.; Heineman, M.J.; Faas, M. 17β-Estradiol and progesterone do not influence the production of cytokines from lipopolysaccharide-stimulated monocytes in humans. Fertil. Steril. 2004, 82, 1212–1219. [Google Scholar] [CrossRef] [PubMed]
- Rogers, A.; Eastell, R. The effect of 17β-estradiol on production of cytokines in cultures of peripheral blood. Bone 2001, 29, 30–34. [Google Scholar] [CrossRef]
- Buyon, J.P.; Korchak, H.M.; Rutherford, L.E.; Ganguly, M.; Weissmann, G. Female hormones reduce neutrophil responsiveness in vitro. Arthritis Rheum. 1984, 27, 623–630. [Google Scholar] [CrossRef] [PubMed]
- Molloy, E.J.; O’Neill, A.J.; Grantham, J.J.; Sheridan-Pereira, M.; Fitzpatrick, J.M.; Webb, D.W.; Watson, R.W.G. Sex-specific alterations in neutrophil apoptosis: The role of estradiol and progesterone. Blood 2003, 102, 2653–2659. [Google Scholar] [CrossRef] [Green Version]
- Relloso, M.; Aragoneses-Fenoll, L.; Lasarte, S.; Bourgeois, C.; Romera, G.; Kuchler, K.; Corbí, A.L.; Muñoz-Fernández, M.A.; Nombela, C.; Rodríguez-Fernández, J.L.; et al. Estradiol impairs the Th17 immune response against Candida albicans. J. Leukoc. Biol. 2012, 91, 159–165. [Google Scholar] [CrossRef]
- Anipindi, V.C.; Bagri, P.; Roth, K.; Dizzell, S.E.; Nguyen, P.V.; Shaler, C.R.; Chu, D.K.; Jiménez-Saiz, R.; Liang, H.; Swift, S.; et al. Estradiol Enhances CD4+ T-Cell Anti-Viral Immunity by Priming Vaginal DCs to Induce Th17 Responses via an IL-1-Dependent Pathway. PLoS Pathog. 2016, 12, 1–27. [Google Scholar] [CrossRef]
- Bengtsson, Å.K.; Ryan, E.J.; Giordano, D.; Magaletti, D.M.; Clark, E.A. 17β-estradiol (E2) modulates cytokine and chemokine expression in human monocyte-derived dendritic cells. Blood 2004, 104, 1404–1410. [Google Scholar] [CrossRef]
- Chen, R.-Y.; Fan, Y.-M.; Zhang, Q.; Liu, S.; Li, Q.; Ke, G.-L.; Li, C.; You, Z. Estradiol Inhibits Th17 Cell Differentiation through Inhibition of RORγT Transcription by Recruiting the ERα/REA Complex to Estrogen Response Elements of the RORγT Promoter. J. Immunol. 2015, 194, 4019–4028. [Google Scholar] [CrossRef] [Green Version]
- Bouman, A.; Jan Heineman, M.; Faas, M.M. Sex hormones and the immune response in humans. Hum. Reprod. Update 2005, 11, 411–423. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lélu, K.; Laffont, S.; Delpy, L.; Paulet, P.-E.; Périnat, T.; Tschanz, S.A.; Pelletier, L.; Engelhardt, B.; Guéry, J.-C. Estrogen Receptor α Signaling in T Lymphocytes Is Required for Estradiol-Mediated Inhibition of Th1 and Th17 Cell Differentiation and Protection against Experimental Autoimmune Encephalomyelitis. J. Immunol. 2011, 187, 2386–2393. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ralston, S.H.; Russell, R.G.G.; Gowen, M. Estrogen inhibits release of tumor necrosis factor from peripheral blood mononuclear cells in postmenopausal women. J. Bone Miner. Res. 1990, 5, 983–988. [Google Scholar] [CrossRef] [PubMed]
- Gilmore, W.; Weiner, L.P.; Correale, J. Effect of estradiol on cytokine secretion by proteolipid protein-specific T cell clones isolated from multiple sclerosis patients and normal control subjects. J. Immunol. 1997, 158, 446–451. [Google Scholar] [PubMed]
- Khan, D.; Ansar Ahmed, S. The immune system is a natural target for estrogen action: Opposing effects of estrogen in two prototypical autoimmune diseases. Front. Immunol. 2016, 6, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Haghmorad, D.; Salehipour, Z.; Nosratabadi, R.; Rastin, M.; Kokhaei, P.; Mahmoudi, M.B.; Amini, A.A.; Mahmoudi, M. Medium-dose estrogen ameliorates experimental autoimmune encephalomyelitis in ovariectomized mice. J. Immunotoxicol. 2016, 13, 885–896. [Google Scholar] [CrossRef] [Green Version]
- Yamanaka, K.; Yamamoto, O.; Honda, T. Pathophysiology of psoriasis: A review. J. Dermatol. 2021, 48, 722–731. [Google Scholar] [CrossRef]
- Danesh, M.; Murase, J.E. The immunologic effects of estrogen on psoriasis: A comprehensive review. Int. J. Women’s Dermatol. 2015, 1, 104–107. [Google Scholar] [CrossRef] [Green Version]
- Iwano, R.; Iwashita, N.; Takagi, Y.; Fukuyama, T. Estrogen receptor α activation aggravates imiquimod-induced psoriasis-like dermatitis in mice by enhancing dendritic cell interleukin-23 secretion. J. Appl. Toxicol. 2020, 40, 1353–1361. [Google Scholar] [CrossRef]
- Adachi, A.; Honda, T.; Egawa, G.; Kanameishi, S.; Takimoto, R.; Miyake, T.; Hossain, M.R.; Komine, M.; Ohtsuki, M.; Gunzer, M.; et al. Estradiol suppresses psoriatic inflammation in mice by regulating neutrophil and macrophage functions. J. Allergy Clin. Immunol. 2022. [Google Scholar] [CrossRef]
- Bouman, A.; Moes, H.; Heineman, M.J.; De Leij, L.F.M.H.; Faas, M.M. The immune response during the luteal phase of the ovarian cycle: Increasing sensitivity of human monocytes to endotoxin. Fertil. Steril. 2001, 76, 555–559. [Google Scholar] [CrossRef]
- Cannon, J.G.; Dinarello, C.A. Increased plasma interleukin-1 activity in women after ovulation. Science 1984, 227, 1247–1249. [Google Scholar] [CrossRef] [PubMed]
- Brännström, M.; Fridén, B.E.; Jasper, M.; Norman, R.J. Variations in peripheral blood levels of immunoreactive tumor necrosis factor α (TNFα) throughout the menstrual cycle and secretion of TNFα from the human corpus luteum. Eur. J. Obstet. Gynecol. Reprod. Biol. 1999, 83, 213–217. [Google Scholar] [CrossRef]
- Shelley, W.B. Generalized Pustular Psoriasis Induced by Potassium Iodide. Jama 1967, 201, 1009. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Prevalence Ratio of Psoriasis | ||
|---|---|---|
| Men | Women | |
| Farber 1974 [32] | 46% | 54% |
| Henseler 1985 [34] | 50.8% | 49.2% |
| Kawada 2003 [26] | 65.80% | 34.20% |
| Takahashi 2009 [25] | 66.40% | 33.60% |
| Tsai 2011 [30] | 61.60% | 38.40% |
| Furue 2011 [24] | 72% | 28% |
| Na 2013 [29] | 54.60% | 45.40% |
| Lee 2017 [28] | 57.30% | 42.70% |
| Hӓgg 2017 [23] | 59.80% | 40.20% |
| Bayaraa 2018 [31] | 67.10% | 32.9% |
| El-komy 2020 [27] | 56.30% | 43.70% |
| Armstrong 2021 [33] | 48.60% | 51.40% |
| Estrogen | |
|---|---|
| Keratinocytes | RANTES↓(physiological to high) [40] |
| CCL-2↓(physiological to high) [39] | |
| CCL-20↓(isoflavone) [42] | |
| S100A7↓(isoflavone) [42] | |
| S100A9↓(isoflavone) [42] | |
| Neutrophils | superoxide anion (O2−)↓(not mentioned) [48] |
| degranulation↓(high) [48] | |
| apoptosis(physiological to high) [49] | |
| migration↓(physiological to high) [49] | |
| Monocytes/Macrophages | IL-1β→~↓(high) [50,51,52] |
| TNF-α→~↓(high) [51,52,53] | |
| Dendritic cells | IL-23↓(high) [54] |
| IL-1β↑(physiological) [55] | |
| IL-8↑(high) [56] | |
| CCL-2↑(high) [56] | |
| T cells | IL-17↓(physiological) [57] |
| TNF-α↓(high), TNF-α↑(low) [58,59] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Adachi, A.; Honda, T. Regulatory Roles of Estrogens in Psoriasis. J. Clin. Med. 2022, 11, 4890. https://doi.org/10.3390/jcm11164890
Adachi A, Honda T. Regulatory Roles of Estrogens in Psoriasis. Journal of Clinical Medicine. 2022; 11(16):4890. https://doi.org/10.3390/jcm11164890
Chicago/Turabian StyleAdachi, Akimasa, and Tetsuya Honda. 2022. "Regulatory Roles of Estrogens in Psoriasis" Journal of Clinical Medicine 11, no. 16: 4890. https://doi.org/10.3390/jcm11164890
APA StyleAdachi, A., & Honda, T. (2022). Regulatory Roles of Estrogens in Psoriasis. Journal of Clinical Medicine, 11(16), 4890. https://doi.org/10.3390/jcm11164890