
Natural Bioactives: Back to the Future in the Fight against Human Papillomavirus? A Narrative Review



Abstract
:1. Introduction
2. Methodology
2.1. Criteria for Reference Selection
2.2. Research Question
2.3. Study Selection
2.4. Charting the Data
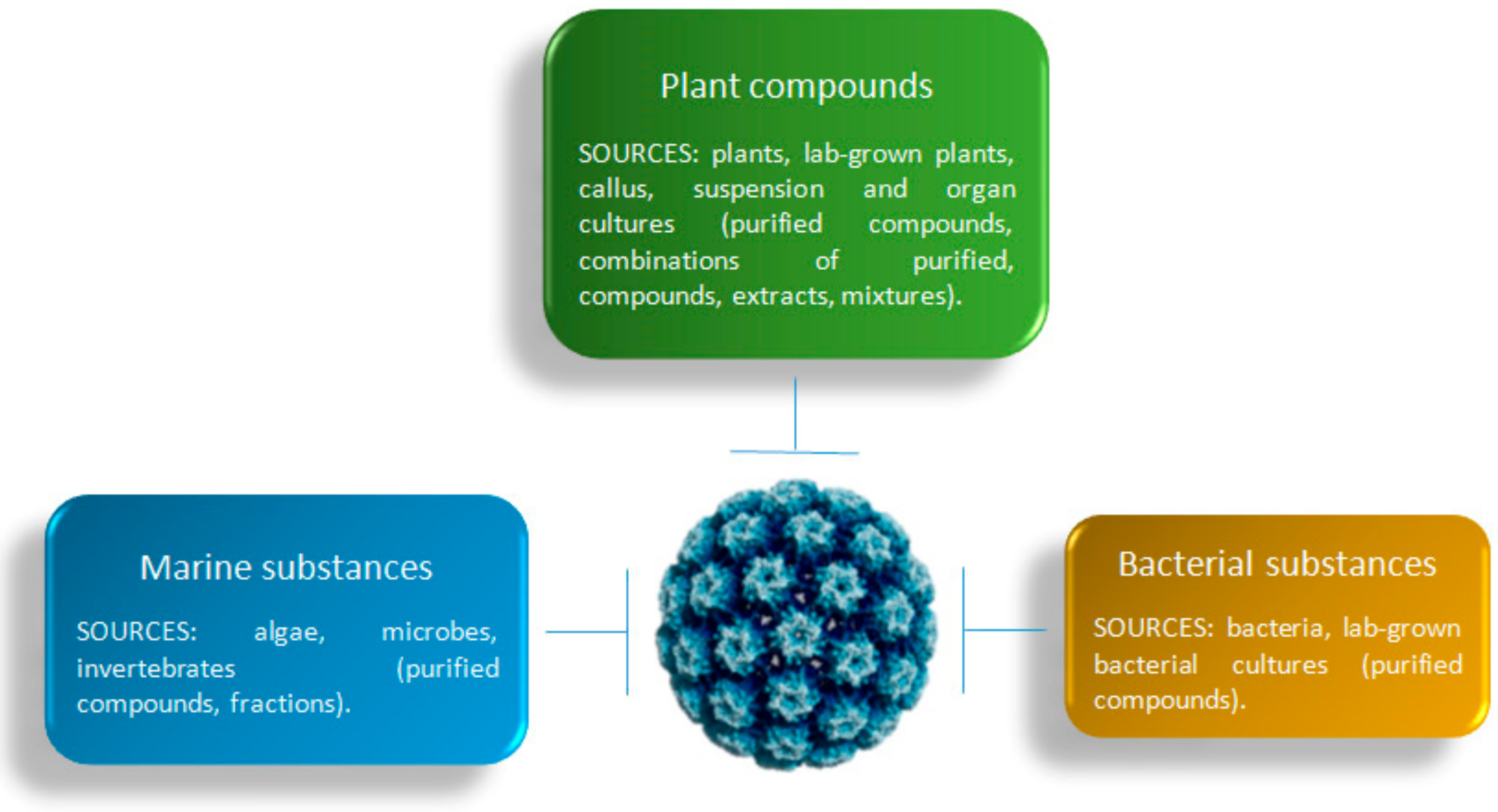
3. Results
4. Purified Phytochemicals
4.1. Polyphenols
4.1.1. Phenolic Acids
4.1.2. Flavonoids
Flavanones
Flavones
Flavonols
Isoflavonoids
Anthocyanins
Flavolignans
Lignans
4.1.3. Stilbenes
4.1.4. Curcuminoids
4.2. Other Specialized Metabolites
4.2.1. Terpenoids
4.2.2. Thiols
4.3. Miscellaneous Specialized Metabolites
4.4. Polysaccharides
4.5. Proteins and Peptides
5. Combinations of Purified Phytocompounds
6. Plant Extracts
7. Plant Extracts Mixtures
8. Phytocompounds with Anti-HPV Potential Inferred by In Silico Approaches
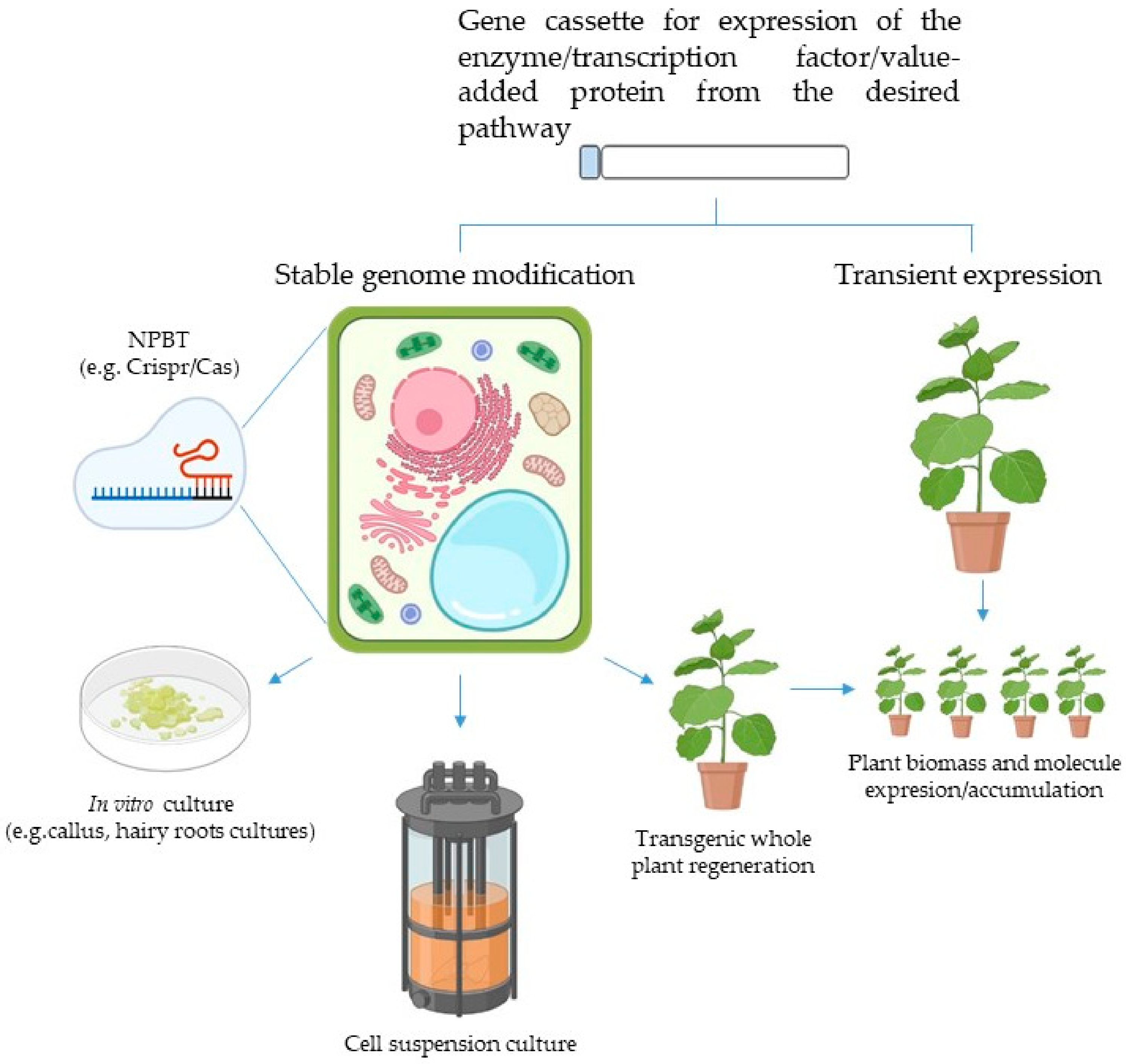
9. The (Plant) Cell Factory
10. Marine Substances
10.1. Algae
10.2. Marine Invertebrates
10.3. Marine Microbes
11. Bacterial Substances
12. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef] [PubMed]
- Ferlay, J.; Colombet, M.; Soerjomataram, I.; Mathers, C.; Parkin, D.M.; Piñeros, M.; Znaor, A.; Bray, F. Estimating the global cancer incidence and mortality in 2018: GLOBOCAN sources and methods. Int. J. Cancer 2018, 144, 1941–1953. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dagenais, G.R.; Leong, D.P.; Rangarajan, S.; Lanas, F.; Lopez-Jaramillo, P.; Gupta, R.; Diaz, R.; Avezum, A.; Oliveira, G.B.F.; Wielgosz, A.; et al. Variations in common diseases, hospital admissions, and deaths in middle-aged adults in 21 countries from five continents (PURE): A prospective cohort study. Lancet 2020, 395, 785–794. [Google Scholar] [CrossRef]
- De Martel, C.; Plummer, M.; Vignat, J.; Franceschi, S. Worldwide burden of cancer attributable to HPV by site, country and HPV type. Int. Agency Res. Cancer (IARC/WHO) 2017, 141, 664–670. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Badaracco, G.; Savarese, A.; Micheli, A.; Rizzo, C.; Paolini, F.; Carosi, M.; Cutillo, G.; Vizza, E.; Arcangeli, G.; Venuti, A. Persistence of HPV after radio-chemotherapy in locally advanced cervical cancer. Oncol. Rep. 2010, 23, 1093–1099. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burd, E.M. Human Papillomavirus and Cervical Cancer. Clin. Microbiol. Rev. 2003, 16, 1–17. [Google Scholar] [CrossRef] [Green Version]
- World Health Organisation. International Agency for Research on Cancer IARC Monographs on the Evaluation of Carcinogenic Risks to Humans. In Human Papillomaviruses; World Health Organisation: Geneva, Switzerland, 2007; Volume 90, ISBN 9789283212904. [Google Scholar]
- Kessler, T.A. Cervical Cancer: Prevention and Early Detection. Semin. Oncol. Nurs. 2017, 33, 172–183. [Google Scholar] [CrossRef]
- Ledford, L.R.C.; Lockwood, S. Scope and Epidemiology of Gynecologic Cancers: An Overview. Semin. Oncol. Nurs. 2019, 35, 147–150. [Google Scholar] [CrossRef]
- Cordeiro, M.N.; De Lima, R.d.C.P.; Paolini, F.; Melo, A.R.d.S.; Campos, A.P.F.; Venuti, A.; De Freitas, A.C. Current research into novel therapeutic vaccines against cervical cancer. Expert Rev. Anticancer Ther. 2018, 18, 365–376. [Google Scholar] [CrossRef]
- Miyauchi, S.; Sanders, P.D.; Guram, K.; Kim, S.S.; Paolini, F.; Venuti, A.; Cohen, E.E.W.; Gutkind, J.S.; Califano, J.A.; Sharabi, A.B. HPV16 E5 Mediates Resistance to PD-L1 Blockade and Can Be Targeted with Rimantadine in Head and Neck Cancer. Cancer Res. 2020, 80, 732–746. [Google Scholar] [CrossRef] [Green Version]
- Gomes, D.; Silvestre, S.; Duarte, A.P.; Venuti, A.; Soares, C.P.; Passarinha, L.; Sousa, Â. In Silico Approaches: A Way to Unveil Novel Therapeutic Drugs for Cervical Cancer Management. Pharmaceuticals 2021, 14, 741. [Google Scholar] [CrossRef] [PubMed]
- Schepisi, G.; Casadei, C.; Toma, I.; Poti, G.; Iaia, M.L.; Farolfi, A.; Conteduca, V.; Lolli, C.; Ravaglia, G.; Brighi, N.; et al. Immunotherapy and Its Development for Gynecological (Ovarian, Endometrial and Cervical) Tumors: From Immune Checkpoint Inhibitors to Chimeric Antigen Receptor (CAR)-T Cell Therapy. Cancers 2021, 13, 840. [Google Scholar] [CrossRef] [PubMed]
- Attademo, L.; Tuninetti, V.; Pisano, C.; Cecere, S.C.; Di Napoli, M.; Tambaro, R.; Valabrega, G.; Musacchio, L.; Setola, S.V.; Piccirillo, P.; et al. Immunotherapy in cervix cancer. Cancer Treat. Rev. 2020, 90, 102088. [Google Scholar] [CrossRef] [PubMed]
- Vici, P.; Pizzuti, L.; Mariani, L.; Zampa, G.; Santini, D.; Di Lauro, L.; Gamucci, T.; Natoli, C.; Marchetti, P.; Barba, M.; et al. Targeting immune response with therapeutic vaccines in premalignant lesions and cervical cancer: Hope or reality from clinical studies. Expert Rev. Vaccines 2016, 15, 1327–1336. [Google Scholar] [CrossRef] [PubMed]
- Vici, P.; Mariani, L.; Pizzuti, L.; Sergi, D.; Di Lauro, L.; Vizza, E.; Tomao, F.; Tomao, S.; Cavallotti, C.; Paolini, F.; et al. Immunologic treatments for precancerous lesions and uterine cervical cancer. J. Exp. Clin. Cancer Res. 2014, 33, 1–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, L.; Wang, Y.; Du, J. Human Papillomavirus Vaccines: An Updated Review. Vaccines 2020, 8, 39. [Google Scholar] [CrossRef] [PubMed]
- Massa, S.; Franconi, R.; Brandi, R.; Muller, A.; Mett, V.; Yusibov, V.; Venuti, A. Anti-cancer activity of plant-produced HPV16 E7 vaccine. Vaccine 2007, 25, 3018–3021. [Google Scholar] [CrossRef]
- Franconi, R.; Di Bonito, P.; Dibello, F.; Accardi, L.; Muller, A.; Cirilli, A.; Simeone, P.; Donà, M.G.; Venuti, A.; Giorgi, C. Plant-derived human papillomavirus 16 E7 oncoprotein induces immune response and specific tumor protection. Cancer Res. 2002, 62, 3654–3658. [Google Scholar]
- Di Bonito, P.; Grasso, F.; Mangino, G.; Massa, S.; Illiano, E.; Franconi, R.; Fanales-Belasio, E.; Falchi, M.; Affabris, E.; Giorgi, C.; et al. Immunomodulatory Activity of a Plant Extract Containing Human Papillomavirus 16-E7 Protein in Human Monocyte-Derived Dendritic Cells. Int. J. Immunopathol. Pharmacol. 2009, 22, 967–978. [Google Scholar] [CrossRef] [Green Version]
- Demurtas, O.C.; Massa, S.; Ferrante, P.; Venuti, A.; Franconi, R.; Giuliano, G. A Chlamydomonas-Derived Human Papillomavirus 16 E7 Vaccine Induces Specific Tumor Protection. PLoS ONE 2013, 8, e61473. [Google Scholar] [CrossRef] [Green Version]
- Franconi, R.; Massa, S.; Paolini, F.; Vici, P.; Venuti, A. Plant-Derived Natural Compounds in Genetic Vaccination and Therapy for HPV-Associated Cancers. Cancers 2020, 12, 3101. [Google Scholar] [CrossRef] [PubMed]
- Moga, M.A.; Dimienescu, O.G.; Arvatescu, C.A.; Mironescu, A.; Dracea, L.; Ples, L. The role of natural polyphenols in the prevention and treatment of cervical cancer—An overview. Molecules 2016, 21, 1055. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Drețcanu, G.; Iuhas, C.I.; Diaconeasa, Z. The involvement of natural polyphenols in the chemoprevention of cervical cancer. Int. J. Mol. Sci. 2021, 22, 8812. [Google Scholar] [CrossRef] [PubMed]
- Di Domenico, F.; Foppoli, C.; Coccia, R.; Perluigi, M. Antioxidants in cervical cancer: Chemopreventive and chemotherapeutic effects of polyphenols. Biochim. Biophys. Acta Mol. Basis Dis. 2012, 1822, 737–747. [Google Scholar] [CrossRef] [Green Version]
- Chen, H.; Liu, R.H. Potential Mechanisms of Action of Dietary Phytochemicals for Cancer Prevention by Targeting Cellular Signaling Transduction Pathways. J. Agric. Food Chem. 2018, 66, 3260–3276. [Google Scholar] [CrossRef]
- Arora, I.; Sharma, M.; Tollefsbol, T.O. Combinatorial Epigenetics Impact of Polyphenols and Phytochemicals in Cancer Prevention and Therapy. Int. J. Mol. Sci. 2019, 20, 4567. [Google Scholar] [CrossRef] [Green Version]
- Liu, C.-J.; Singh, S.; Sharma, B.; Kanwar, S.S.; Kumar, A. Lead Phytochemicals for Anticancer Drug Development. Front. Plant Sci. 2016, 7, 1667. [Google Scholar] [CrossRef] [Green Version]
- Kotecha, R.; Takami, A.; Espinoza, J.L. Dietary Phytochemicals and Cancer Chemoprevention: A Review of the Clinical Evidence. Oncotarget 2016, 7, 52517. [Google Scholar] [CrossRef] [Green Version]
- Dubensky, T.W.; Kanne, D.B.; Leong, M.L. Rationale, progress and development of vaccines utilizing STING-activating cyclic dinucleotide adjuvants. Ther. Adv. Vaccines 2013, 1, 131–143. [Google Scholar] [CrossRef]
- Barber, G.N. Cytoplasmic DNA innate immune pathways. Immunol. Rev. 2011, 243, 99–108. [Google Scholar] [CrossRef]
- Pilla, L.; Rivoltini, L.; Patuzzo, R.; Marrari, A.; Valdagni, R.; Parmiani, G. Multipeptide vaccination in cancer patients. Expert Opin. Biol. Ther. 2009, 9, 1043–1055. [Google Scholar] [CrossRef] [PubMed]
- Mett, V.; Farrance, C.E.; Green, B.J.; Yusibov, V. Plants as biofactories. Biologicals 2008, 36, 354–358. [Google Scholar] [CrossRef] [PubMed]
- Wu, T.; Kerbler, S.M.; Fernie, A.R.; Zhang, Y. Plant cell cultures as heterologous bio-factories for secondary metabolite production. Plant Commun. 2021, 2, 100235. [Google Scholar] [CrossRef] [PubMed]
- Chang, W.-C.; Hsieh, C.-H.; Hsiao, M.-W.; Lin, W.-C.; Hung, Y.-C.; Ye, J.-C. Caffeic acid induces apoptosis in human cervical cancer cells through the mitochondrial pathway. Taiwan. J. Obstet. Gynecol. 2010, 49, 419–424. [Google Scholar] [CrossRef] [Green Version]
- Anantharaju, P.G.; Reddy, D.B.; Padukudru, M.A.; Kumari Chitturi, C.M.; Vimalambike, M.G.; Madhunapantula, S.V. Induction of colon and cervical cancer cell death by cinnamic acid derivatives is mediated through the inhibition of Histone Deacetylases (HDAC). PLoS ONE 2017, 12, e0186208. [Google Scholar] [CrossRef]
- Guo, H.; Zhang, D.; Fu, Q. Inhibition of cervical cancer by promoting IGFBP7 expression using ellagic acid from pomegranate peel. Med. Sci. Monit. 2016, 22, 4881–4886. [Google Scholar] [CrossRef] [Green Version]
- Gao, J.; Yu, H.; Guo, W.; Kong, Y.; Li, Q.; Yang, S.; Zhang, Y.; Wang, Y. The anticancer effects of ferulic acid is associated with induction of cell cycle arrest and autophagy in cervical cancer cells. Cancer Cell Int. 2018, 18, 102. [Google Scholar] [CrossRef] [Green Version]
- Karthikeyan, S.; Kanimozhi, G.; Prasad, N.R.; Mahalakshmi, R. Radiosensitizing effect of ferulic acid on human cervical carcinoma cells in vitro. Toxicol. In Vitro 2011, 25, 1366–1375. [Google Scholar] [CrossRef]
- You, B.R.; Moon, H.J.; Han, Y.H.; Park, W.H. Gallic acid inhibits the growth of HeLa cervical cancer cells via apoptosis and/or necrosis. Food Chem. Toxicol. 2010, 48, 1334–1340. [Google Scholar] [CrossRef]
- Zhao, B.; Hu, M. Gallic acid reduces cell viability, proliferation, invasion and angiogenesis in human cervical cancer cells. Oncol. Lett. 2013, 6, 1749–1755. [Google Scholar] [CrossRef] [Green Version]
- Alshatwi, A.A.; Ramesh, E.; Periasamy, V.S.; Subash-Babu, P. The apoptotic effect of hesperetin on human cervical cancer cells is mediated through cell cycle arrest, death receptor, and mitochondrial pathways. Fundam. Clin. Pharmacol. 2013, 27, 581–592. [Google Scholar] [CrossRef] [PubMed]
- Roohbakhsh, A.; Parhiz, H.; Soltani, F.; Rezaee, R.; Iranshahi, M. Molecular mechanisms behind the biological effects of hesperidin and hesperetin for the prevention of cancer and cardiovascular diseases. Life Sci. 2015, 124, 64–74. [Google Scholar] [CrossRef] [PubMed]
- Ramesh, E.; Alshatwi, A.A. Naringin induces death receptor and mitochondria-mediated apoptosis in human cervical cancer (SiHa) cells. Food Chem. Toxicol. 2013, 51, 97–105. [Google Scholar] [CrossRef]
- Krishnakumar, N.; Sulfikkarali, N.; RajendraPrasad, N.; Karthikeyan, S. Enhanced anticancer activity of naringenin-loaded nanoparticles in human cervical (HeLa) cancer cells. Biomed. Prev. Nutr. 2011, 1, 223–231. [Google Scholar] [CrossRef]
- Zeng, L.; Zhen, Y.; Chen, Y.; Zou, L.; Zhang, Y.; Hu, F.; Feng, J.; Shen, J.; Wei, B. Naringin inhibits growth and induces apoptosis by a mechanism dependent on reduced activation of NF-κB/COX-2-caspase-1 pathway in HeLa cervical cancer cells. Int. J. Oncol. 2014, 45, 1929–1936. [Google Scholar] [CrossRef] [Green Version]
- Vanamala, J.; Leonardi, T.; Patil, B.S.; Taddeo, S.S.; Murphy, M.E.; Pike, L.M.; Chapkin, R.S.; Lupton, J.R.; Turner, N.D. Suppression of colon carcinogenesis by bioactive compounds in grapefruit. Carcinogenesis 2006, 27, 1257–1265. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, D.I.; Lee, S.J.; Lee, S.B.; Park, K.; Kim, W.J.; Moon, S.K. Requirement for Ras/Raf/ERK pathway in naringin-induced G1 -cell-cycle arrest via p21WAF1 expression. Carcinogenesis 2008, 29, 1701–1709. [Google Scholar] [CrossRef] [Green Version]
- Manach, C.; Scalbert, A.; Morand, C.; Rémésy, C.; Jiménez, L. Polyphenols: Food sources and bioavailability. Am. J. Clin. Nutr. 2004, 79, 727–747. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, C.S.; Landau, J.M.; Huang, M.T.; Newmark, H.L. Inhibition of carcinogenesis by dietary polyphenolic compounds. Annu. Rev. Nutr. 2001, 21, 381–406. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Czeczot, H.; Tudek, B.; Kusztelak, J.; Szymczyk, T.; Dobrowolska, B.; Glinkowska, G.; Malinowski, J.; Strzelecka, H. Isolation and studies of the mutagenic activity in the Ames test of flavonoids naturally occurring in medical herbs. Mutat. Res. Toxicol. 1990, 240, 209–216. [Google Scholar] [CrossRef]
- Del Rio, D.; Rodriguez-Mateos, A.; Spencer, J.P.E.; Tognolini, M.; Borges, G.; Crozier, A. Dietary (poly)phenolics in human health: Structures, bioavailability, and evidence of protective effects against chronic diseases. Antioxid. Redox Signal. 2013, 18, 1818–1892. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodriguez-Mateos, A.; Vauzour, D.; Krueger, C.G.; Shanmuganayagam, D.; Reed, J.; Calani, L.; Mena, P.; Del Rio, D.; Crozier, A. Bioavailability, bioactivity and impact on health of dietary flavonoids and related compounds: An update. Arch. Toxicol. 2014, 88, 1803–1853. [Google Scholar] [CrossRef] [PubMed]
- Brglez Mojzer, E.; Knez Hrnčič, M.; Škerget, M.; Knez, Ž.; Bren, U. Polyphenols: Extraction Methods, Antioxidative Action, Bioavailability and Anticarcinogenic Effects. Molecules 2016, 21, 901. [Google Scholar] [CrossRef] [PubMed]
- Zheng, P.W.; Chiang, L.C.; Lin, C.C. Apigenin induced apoptosis through p53-dependent pathway in human cervical carcinoma cells. Life Sci. 2005, 76, 1367–1379. [Google Scholar] [CrossRef]
- Czyz, J.; Madeja, Z.; Korohoda, W.; Hü Lser, D.F. Flavonoid apigenin inhibits motility and invasiveness of carcinoma cells in vitro. Int. J. Cancer 2004, 114, 12–18. [Google Scholar] [CrossRef]
- Xu, Y.; Xin, Y.; Diao, Y.; Lu, C.; Fu, J.; Luo, L.; Yin, Z. Synergistic Effects of Apigenin and Paclitaxel on Apoptosis of Cancer Cells. PLoS ONE 2011, 6, e29169. [Google Scholar] [CrossRef] [Green Version]
- Chuang, C.M.; Monie, A.; Wu, A.; Hung, C.F. Combination of apigenin treatment with therapeutic HPV DNA vaccination generates enhanced therapeutic antitumor effects. J. Biomed. Sci. 2009, 16, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Lee, H.G.; Yu, K.A.; Oh, W.K.; Baeg, T.W.; Oh, H.C.; Ahn, J.S.; Jang, W.C.; Kim, J.W.; Lim, J.S.; Choe, Y.K.; et al. Inhibitory effect of jaceosidin isolated from Artemisia argyi on the function of E6 and E7 oncoproteins of HPV 16. J. Ethnopharmacol. 2005, 98, 339–343. [Google Scholar] [CrossRef]
- Han, H.Y.; Kim, H.J.; Jeong, S.H.; Kim, J.; Jeong, S.H.; Kim, G.C.; Hwang, D.S.; Kim, U.K.; Ryu, M.H. The Flavonoid Jaceosidin from Artemisia princeps Induces Apoptotic Cell Death and Inhibits the Akt Pathway in Oral Cancer Cells. Evid.-Based Complement. Altern. Med. 2018, 2018, 5765047. [Google Scholar] [CrossRef] [Green Version]
- Cherry, J.J.; Rietz, A.; Malinkevich, A.; Liu, Y.; Xie, M.; Bartolowits, M.; Davisson, V.J.; Baleja, J.D.; Androphy, E.J. Structure Based Identification and Characterization of Flavonoids That Disrupt Human Papillomavirus-16 E6 Function. PLoS ONE 2013, 8, 84506. [Google Scholar] [CrossRef]
- Kim, M.S.; Bak, Y.; Park, Y.S.; Lee, D.H.; Kim, J.H.; Kang, J.W.; Song, H.-H.; Oh, S.-R.; Yoon, D.Y. Wogonin induces apoptosis by suppressing E6 and E7 expressions and activating intrinsic signaling pathways in HPV-16 cervical cancer cells. Cell Biol. Toxicol. 2013, 29, 259–272. [Google Scholar] [CrossRef] [PubMed]
- He, F.; Wang, Q.; Zheng, X.-L.; Yan, J.-Q.; Yang, L.; Sun, H.; Hu, L.-N.; Lin, Y.; Wang, X. Wogonin potentiates cisplatin-induced cancer cell apoptosis through accumulation of intracellular reactive oxygen species. Oncol. Rep. 2012, 28, 601–605. [Google Scholar] [CrossRef] [PubMed]
- Hussain, S.; Ashafaq, M. Epigallocatechin-3-Gallate (EGCG): Mechanisms, perspectives and clinical applications in cervical cancer. J. Cancer Prev. Curr. Res. 2018, 9, 178–182. [Google Scholar] [CrossRef]
- Qiao, Y.; Cao, J.; Xie, L.; Shi, X. Cell Growth Inhibition and Gene Expression Regulation by (-)-Epigal-locatechin-3-Gallate in Human Cervical Cancer Cells. Arch. Pharm Res. 2009, 32, 1309–1315. [Google Scholar] [CrossRef] [PubMed]
- Sharma, C.; Nusri, Q.E.A.; Begum, S.; Javed, E.; Rizvi, T.A.; Hussain, A. (-)-Epigallocatechin-3-gallate induces apoptosis and inhibits invasion and migration of human cervical cancer cells. Asian Pac. J. Cancer Prev. 2012, 13, 4815–4822. [Google Scholar] [CrossRef] [Green Version]
- Zou, C.; Liu, H.; Feugang, J.M.; Hao, Z.; Chow, H.H.S.; Garcia, F. Green tea compound in chemoprevention of cervical cancer. Int. J. Gynecol. Cancer 2010, 20, 617–624. [Google Scholar] [CrossRef] [Green Version]
- Chen, L.; Han, W.; Geng, Y.; Su, J. A genome-wide study of DNA methylation modified by epigallocatechin-3-gallate in the CAL-27 cell line. Mol. Med. Rep. 2015, 12, 5886–5890. [Google Scholar] [CrossRef] [Green Version]
- Zhu, Y.; Huang, Y.; Liu, M.; Yan, Q.; Zhao, W.; Yang, P.; Gao, Q.; Wei, J.; Zhao, W.; Ma, L. Epigallocatechin gallate inhibits cell growth and regulates miRNA expression in cervical carcinoma cell lines infected with different high-risk human papillomavirus subtypes. Exp. Ther. Med. 2019, 17, 1742–1748. [Google Scholar] [CrossRef] [Green Version]
- Alshatwi, A.A.; Periasamy, V.S.; Athinarayanan, J.; Elango, R. Synergistic anticancer activity of dietary tea polyphenols and bleomycin hydrochloride in human cervical cancer cell: Caspase-dependent and independent apoptotic pathways. Chem. Biol. Interact. 2016, 247, 1–10. [Google Scholar] [CrossRef]
- Kang, T.H.; Lee, J.H.; Song, C.K.; Han, H.D.; Shin, B.C.; Pai, S.I.; Hung, C.-F.; Trimble, C.; Lim, J.-S.; Kim, T.W.; et al. Epigallocatechin-3-Gallate Enhances CD8+ T Cell–Mediated Antitumor Immunity Induced by DNA Vaccination. Cancer Res. 2007, 67, 802–811. [Google Scholar] [CrossRef] [Green Version]
- Ying, T.H.; Yang, S.F.; Tsai, S.J.; Hsieh, S.C.; Huang, Y.C.; Bau, D.T.; Hsieh, Y.H. Fisetin induces apoptosis in human cervical cancer HeLa cells through ERK1/2-mediated activation of caspase-8-/caspase-3-dependent pathway. Arch. Toxicol. 2012, 86, 263–273. [Google Scholar] [CrossRef] [PubMed]
- Xu, W.; Liu, J.; Li, C.; Wu, H.Z.; Liu, Y.W. Kaempferol-7-O-β-d-glucoside (KG) isolated from Smilax china L. rhizome induces G2/M phase arrest and apoptosis on HeLa cells in a p53-independent manner. Cancer Lett. 2008, 264, 229–240. [Google Scholar] [CrossRef] [PubMed]
- Tu, L.Y.; Bai, H.H.; Cai, J.Y.; Deng, S.P. The mechanism of kaempferol induced apoptosis and inhibited proliferation in human cervical cancer SiHa cell: From macro to nano. Scanning 2016, 38, 644–653. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vidya Priyadarsini, R.; Senthil Murugan, R.; Maitreyi, S.; Ramalingam, K.; Karunagaran, D.; Nagini, S. The flavonoid quercetin induces cell cycle arrest and mitochondria-mediated apoptosis in human cervical cancer (HeLa) cells through p53 induction and NF-κB inhibition. Eur. J. Pharmacol. 2010, 649, 84–91. [Google Scholar] [CrossRef]
- Jakubowicz-Gil, J.; Paduch, R.; Piersiak, T.; Głowniak, K.; Gawron, A.; Kandefer-Szerszeń, M. The effect of quercetin on pro-apoptotic activity of cisplatin in HeLa cells. Biochem. Pharmacol. 2005, 69, 1343–1350. [Google Scholar] [CrossRef]
- Lin, C.; Yu, Y.; Zhao, H.G.; Yang, A.; Yan, H.; Cui, Y. Combination of quercetin with radiotherapy enhances tumor radiosensitivity in vitro and in vivo. Radiother. Oncol. 2012, 104, 395–400. [Google Scholar] [CrossRef]
- Guo, J.M.; Kang, G.Z.; Xiao, B.X.; Liu, D.H.; Zhang, S. Effect of daidzein on cell growth, cell cycle, and telomerase activity of human cervical cancer in vitro. Int. J. Gynecol. Cancer 2004, 14, 882–888. [Google Scholar] [CrossRef]
- Lo, Y.L.; Wang, W. Formononetin potentiates epirubicin-induced apoptosis via ROS production in HeLa cells in vitro. Chem. Biol. Interact. 2013, 205, 188–197. [Google Scholar] [CrossRef]
- Yashar, C.M.; Spanos, W.J.; Taylor, D.D.; Gercel-Taylor, C. Potentiation of the radiation effect with genistein in cervical cancer cells. Gynecol. Oncol. 2005, 99, 199–205. [Google Scholar] [CrossRef]
- Zhang, B.; Liu, J.-Y.; Pan, J.-S.; Han, S.-P.; Yin, X.-X.; Wang, B.; Hu, G. Combined Treatment of Ionizing Radiation With Genistein on Cervical Cancer HeLa Cells. J. Pharmacol. Sci. 2006, 102, 129–135. [Google Scholar] [CrossRef] [Green Version]
- Shin, J.I.; Shim, J.H.; Kim, K.H.; Choi, H.S.; Kim, J.W.; Lee, H.G.; Kim, B.Y.; Park, S.N.; Park, O.J.; Yoon, D.Y. Sensitization of the apoptotic effect of γ-irradiation in genistein-pretreated CaSki cervical cancer cells. J. Microbiol. Biotechnol. 2008, 18, 523–531. [Google Scholar] [PubMed]
- Zheng, W.-Y.; Nie, S.-P.; Li, W.-J.; Hu, X.-J.; Xie, M.-Y. Stimulatory effects of genistein and quercetin on the proliferation of MCF-7 cells. Food Biosci. 2013, 2, 15–23. [Google Scholar] [CrossRef]
- Liu, H.; Du, J.; Hu, C.; Qi, H.; Wang, X.; Wang, S.; Liu, Q.; Li, Z. Delayed activation of extracellular-signal-regulated kinase 1/2 is involved in genistein- and equol-induced cell proliferation and estrogen-receptor-α-mediated transcription in MCF-7 breast cancer cells. J. Nutr. Biochem. 2010, 21, 390–396. [Google Scholar] [CrossRef] [PubMed]
- Qian, S.; Jin, L.-Q. The Studies of Cyanidin 3-Glucoside-Induced Apoptosis in Human Cervical Cancer Hela Cells and its Mechanism. Chin. J. Biochem. Pharm. 2008, 6, 369–373. [Google Scholar]
- Zhang, Y.; Ge, Y.; Chen, Y.; Li, Q.; Chen, J.; Dong, Y.; Shi, W. Cellular and molecular mechanisms of silibinin induces cell-cycle arrest and apoptosis on HeLa cells. Cell Biochem. Funct. 2012, 30, 243–248. [Google Scholar] [CrossRef]
- You, Y.; He, Q.; Lu, H.; Zhou, X.; Chen, L.; Liu, H.; Lu, Z.; Liu, D.; Liu, Y.; Zuo, D.; et al. Silibinin Induces G2/M Cell Cycle Arrest by Activating Drp1-Dependent Mitochondrial Fission in Cervical Cancer. Front. Pharmacol. 2020, 11, 271. [Google Scholar] [CrossRef] [Green Version]
- Giridharan, P.; Somasundaram, S.T.; Perumal, K.; Vishwakarma, R.A.; Karthikeyan, N.P.; Velmurugan, R.; Balakrishnan, A. Novel substituted methylenedioxy lignan suppresses proliferation of cancer cells by inhibiting telomerase and activation of c-myc and caspases leading to apoptosis. Br. J. Cancer 2002, 87, 98–105. [Google Scholar] [CrossRef]
- Gao, P.; Zhai, F.; Guan, L.; Zheng, J. Nordihydroguaiaretic acid inhibits growth of cervical cancer SiHa cells by upregulating p21. Oncol. Lett. 2011, 2, 123–128. [Google Scholar] [CrossRef] [Green Version]
- Dou, H.; Yang, S.; Hu, Y.; Xu, D.; Liu, L.; Li, X. Sesamin induces ER stress-mediated apoptosis and activates autophagy in cervical cancer cells. Life Sci. 2018, 200, 87–93. [Google Scholar] [CrossRef]
- Kim, Y.S.; Sull, J.W.; Sung, H.J. Suppressing effect of resveratrol on the migration and invasion of human metastatic lung and cervical cancer cells. Mol. Biol. Rep. 2012, 39, 8709–8716. [Google Scholar] [CrossRef]
- García-Zepeda, S.P.; García-Villa, E.; Díaz-Chávez, J.; Hernández-Pando, R.; Gariglio, P. Resveratrol induces cell death in cervical cancer cells through apoptosis and autophagy. Eur. J. Cancer Prev. 2013, 22, 577–584. [Google Scholar] [CrossRef]
- Zoberi, I.; Bradbury, C.M.; Curry, H.A.; Bisht, K.S.; Goswami, P.C.; Roti, J.L.; Gius, D. Radiosensitizing and anti-proliferative effects of resveratrol in two human cervical tumor cell lines. Cancer Lett. 2002, 175, 165–173. [Google Scholar] [CrossRef]
- Shaito, A.; Posadino, A.M.; Younes, N.; Hasan, H.; Halabi, S.; Alhababi, D.; Al-Mohannadi, A.; Abdel-Rahman, W.M.; Eid, A.H.; Nasrallah, G.K.; et al. Potential adverse effects of resveratrol: A literature review. Int. J. Mol. Sci. 2020, 21, 2084. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Divya, C.S.; Pillai, M.R. Antitumor action of curcumin in human papillomavirus associated cells involves downregulation of viral oncogenes, prevention of NFkB and AP-1 translocation, and modulation of apoptosis. Mol. Carcinog. 2006, 45, 320–332. [Google Scholar] [CrossRef] [PubMed]
- Maher, D.M.; Bell, M.C.; O’Donnell, E.A.; Gupta, B.K.; Jaggi, M.; Chauhan, S.C. Curcumin suppresses human papillomavirus oncoproteins, restores p53, rb, and ptpn13 proteins and inhibits benzo[a]pyrene-induced upregulation of HPV E7. Mol. Carcinog. 2011, 50, 47–57. [Google Scholar] [CrossRef] [PubMed]
- Anto, R.J.; Mukhopadhyay, A.; Denning, K.; Aggarwal, B.B. Curcumin (diferuloylmethane) induces apoptosis through activation of caspase-8, BID cleavage and cytochrome c release: Its suppression by ectopic expression of Bcl-2 and Bcl-xl. Carcinogenesis 2002, 23, 143–150. [Google Scholar] [CrossRef] [PubMed]
- Paulraj, F.; Abas, F.; Lajis, N.H.; Othman, I.; Hassan, S.S.; Naidu, R. The curcumin analogue 1,5-bis(2-hydroxyphenyl)-1,4-pentadiene-3-one induces apoptosis and downregulates E6 and E7 oncogene expression in HPV16 and HPV18-infected cervical cancer cells. Molecules 2015, 20, 11830–11860. [Google Scholar] [CrossRef]
- Bava, S.V.; Puliappadamba, V.T.; Deepti, A.; Nair, A.; Karunagaran, D.; Anto, R.J. Sensitization of taxol-induced apoptosis by curcumin involves downregulation of nuclear factor-κB and the serine/threonine kinase Akt and is independent of tubulin polymerization. J. Biol. Chem. 2005, 280, 6301–6308. [Google Scholar] [CrossRef] [Green Version]
- Bava, S.V.; Sreekanth, C.N.; Thulasidasan, A.K.T.; Anto, N.P.; Cheriyan, V.T.; Puliyappadamba, V.T.; Menon, S.G.; Ravichandran, S.D.; Anto, R.J. Akt is upstream and MAPKs are downstream of NF-κB in paclitaxel-induced survival signaling events, which are downregulated by curcumin contributing to their synergism. Int. J. Biochem. Cell Biol. 2011, 43, 331–341. [Google Scholar] [CrossRef]
- Prusty, B.K.; Das, B.C. Constitutive activation of transcription factor AP-1 in cervical cancer and suppression of human papillomavirus (HPV) transcription and AP-1 activity in HeLa cells by curcumin. Int. J. Cancer 2005, 113, 951–960. [Google Scholar] [CrossRef]
- Limtrakul, P.; Chearwae, W.; Shukla, S.; Phisalphong, C.; Ambudkar, S.V. Modulation of function of three ABC drug transporters, P-glycoprotein (ABCB1), mitoxantrone resistance protein (ABCG2) and multidrug resistance protein 1 (ABCC1) by tetrahydrocurcumin, a major metabolite of curcumin. Mol. Cell. Biochem. 2007, 296, 85–95. [Google Scholar] [CrossRef] [PubMed]
- Javvadi, P.; Hertan, L.; Kosoff, R.; Datta, T.; Kolev, J.; Mick, R.; Tuttle, S.W.; Koumenis, C. Thioredoxin reductase-1 mediates curcumin-induced radiosensitization of squamous carcinoma cells. Cancer Res. 2010, 70, 1941–1950. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, A.L.; Hsu, C.H.; Lin, J.K.; Hsu, M.M.; Ho, Y.F.; Shen, T.S.; Ko, J.Y.; Lin, J.T.; Lin, B.R.; Ming-Shiang, W.; et al. Phase I clinical trial of curcumin, a chemopreventive agent, in patients with high-risk or pre-malignant lesions. Anticancer Res. 2001, 21, 2895–2900. [Google Scholar]
- Verreault, R.; Chu, J.; Mandelson, M.; Shy, K. A case-control study of diet and invasive cervical cancer. Int. J. Cancer 1989, 43, 1050–1054. [Google Scholar] [CrossRef] [PubMed]
- Herrero, R.; Potischman, N.; Brinton, L.A.; Reeves, W.C.; Brenes, M.M.; Tenorio, F.; de Britton, R.C.; Gaitan, E. A Case-Control Study of Nutrient Status and Invasive Cervical Cancer: I. Dietary Indicators. Am. J. Epidemiol. 1991, 134, 1335–1346. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, C.; Baker, J.A.; Moysich, K.B.; Rivera, R.; Brasure, J.R.; McCann, S.E. Dietary intakes of selected nutrients and food groups and risk of cervical cancer. Nutr. Cancer 2008, 60, 331–341. [Google Scholar] [CrossRef]
- Tomita, L.Y.; Longatto Filho, A.; Costa, M.C.; Andreoli, M.A.A.; Villa, L.L.; Franco, E.L.; Cardoso, M.A. Diet and serum micronutrients in relation to cervical neoplasia and cancer among low-income Brazilian women. Int. J. Cancer 2010, 126, 703–714. [Google Scholar] [CrossRef]
- González, C.A.; Travier, N.; Luján-Barroso, L.; Castellsagué, X.; Bosch, F.X.; Roura, E.; Bueno-de-Mesquita, H.B.; Palli, D.; Boeing, H.; Pala, V.; et al. Dietary factors and in situ and invasive cervical cancer risk in the European prospective investigation into cancer and nutrition study. Int. J. Cancer 2011, 129, 449–459. [Google Scholar] [CrossRef]
- Batieha, A.M.; Armenian, H.K.; Norkus, E.P.; Morris, J.S.; Spate, V.E.; Comstock, G.W. Serum micronutrients and the subsequent risk of cervical cancer in a population-based nested case-control study. Cancer Epidemiol. Biomark. Prev. 1993, 2, 335–339. [Google Scholar]
- Lehtinen, M.; Luostarinen, T.; Youngman, L.D.; Anttila, T.; Dillner, J.; Hakulinen, T.; Koskela, P.; Lenner, P.; Hallmans, G. Low levels of serum vitamins A and E in blood and subsequent risk for cervical cancer: Interaction with HPV seropositivity. Nutr. Cancer 1999, 34, 229–234. [Google Scholar] [CrossRef]
- Zhang, Y.Y.; Lu, L.; Abliz, G.; Mijit, F. Serum carotenoid, retinol and tocopherol concentrations and risk of cervical cancer among chinese women. Asian Pac. J. Cancer Prev. 2015, 16, 2981–2986. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ratnam, D.V.; Ankola, D.D.; Bhardwaj, V.; Sahana, D.K.; Kumar, M.N.V.R. Role of antioxidants in prophylaxis and therapy: A pharmaceutical perspective. J. Control. Release 2006, 113, 189–207. [Google Scholar] [CrossRef] [PubMed]
- Aktepe, O.H.; Şahin, T.K.; Güner, G.; Arik, Z.; Yalçin, Ş. Lycopene sensitizes the cervical cancer cells to cisplatin via targeting nuclear factor- kappa B (NF-κB) pathway. Turk. J. Med. Sci. 2021, 51, 368–374. [Google Scholar] [CrossRef] [PubMed]
- Overwijk, W.W.; Wang, E.; Marincola, F.M.; Rammensee, H.G.; Restifo, N.P. Mining the mutanome: Developing highly personalized Immunotherapies based on mutational analysis of tumors. J. Immunother. Cancer 2013, 1, 11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Q.; Zheng, X.L.; Yang, L.; Shi, F.; Gao, L.B.; Zhong, Y.J.; Sun, H.; He, F.; Lin, Y.; Wang, X. Reactive oxygen species-mediated apoptosis contributes to chemosensitization effect of saikosaponins on cisplatin-induced cytotoxicity in cancer cells. J. Exp. Clin. Cancer Res. 2010, 29, 159. [Google Scholar] [CrossRef] [Green Version]
- Yuan, B.; Yang, R.; Ma, Y.; Zhou, S.; Zhang, X.; Liu, Y. A systematic review of the active saikosaponins and extracts isolated from Radix Bupleuri and their applications. Pharm. Biol. 2017, 55, 620–635. [Google Scholar] [CrossRef] [Green Version]
- Zhang, W.; Zhang, D.; Ma, X.; Liu, Z.; Li, F.; Wu, D. Paris saponin VII suppressed the growth of human cervical cancer Hela cells. Eur. J. Med. Res. 2014, 19, 41. [Google Scholar] [CrossRef] [Green Version]
- Munagala, R.; Aqil, F.; Jeyabalan, J.; Gupta, R.C. Tanshinone IIA inhibits viral oncogene expression leading to apoptosis and inhibition of cervical cancer. Cancer Lett. 2015, 356, 536–546. [Google Scholar] [CrossRef]
- Guo, J.-L.; Han, T.; Bao, L.; Li, X.-M.; Ma, J.-Q.; Tang, L.-P. Ursolic acid promotes the apoptosis of cervical cancer cells by regulating endoplasmic reticulum stress. J. Obstet. Gynaecol. Res. 2019, 45, 877–881. [Google Scholar] [CrossRef]
- Wang, S.; Meng, X.; Dong, Y. Ursolic acid nanoparticles inhibit cervical cancer growth in vitro and in vivo via apoptosis induction. Int. J. Oncol. 2017, 50, 1330–1340. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Q.; Yang, D. Allicin suppresses the migration and invasion in cervical cancer cells mainly by inhibiting NRF2. Exp. Ther. Med. 2018, 17, 1523–1528. [Google Scholar] [CrossRef]
- Chen, D.Z.; Qi, M.; Auborn, K.J.; Carter, T.H. Indole-3-carbinol and diindolylmethane induce apoptosis of human cervical cancer cells and in murine HPV16-transgenic preneoplastic cervical epithelium. J. Nutr. 2001, 131, 3294–3302. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rosen, C.A.; Woodson, G.E.; Thompson, J.W.; Hengesteg, A.P.; Bradlow, H.L. Preliminary results of the use of indole-3-carbinol for recurrent respiratory papillomatosis. Otolaryngol. Neck Surg. Off. J. Am. Acad. Otolaryngol. Neck Surg. 1998, 118, 810–815. [Google Scholar] [CrossRef]
- Bell, M.C.; Crowley-Nowick, P.; Bradlow, H.L.; Sepkovic, D.W.; Schmidt-Grimminger, D.; Howell, P.; Mayeaux, E.J.; Tucker, A.; Turbat-Herrera, E.A.; Mathis, J.M. Placebo-controlled trial of indole-3-carbinol in the treatment of CIN. Gynecol. Oncol. 2000, 78, 123–129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rosen, C.A.; Bryson, P.C. Indole-3-carbinol for recurrent respiratory papillomatosis: Long-term results. J. Voice 2004, 18, 248–253. [Google Scholar] [CrossRef]
- Del Priore, G.; Gudipudi, D.K.; Montemarano, N.; Restivo, A.M.; Malanowska-Stega, J.; Arslan, A.A. Oral diindolylmethane (DIM): Pilot evaluation of a nonsurgical treatment for cervical dysplasia. Gynecol. Oncol. 2010, 116, 464–467. [Google Scholar] [CrossRef]
- Castañon, A.; Tristram, A.; Mesher, D.; Powell, N.; Beer, H.; Ashman, S.; Rieck, G.; Fielder, H.; Fiander, A.; Sasieni, P. Effect of diindolylmethane supplementation on low-grade cervical cytological abnormalities: Double-blind, randomised, controlled trial. Br. J. Cancer 2012, 106, 45–52. [Google Scholar] [CrossRef]
- Ashrafian, L.; Sukhikh, G.; Kiselev, V.; Paltsev, M.; Drukh, V.; Kuznetsov, I.; Muyzhnek, E.; Apolikhina, I.; Andrianova, E. Double-blind randomized placebo-controlled multicenter clinical trial (phase IIa) on diindolylmethane’s efficacy and safety in the treatment of CIN: Implications for cervical cancer prevention. EPMA J. 2015, 6, 25. [Google Scholar] [CrossRef] [Green Version]
- Wang, S.J.; Zheng, C.J.; Peng, C.; Zhang, H.; Jiang, Y.P.; Han, T.; Qin, L.P. Plants and cervical cancer: An overview. Expert Opin. Investig. Drugs 2013, 22, 1133–1156. [Google Scholar] [CrossRef]
- Cheng, Y.-M.; Tsai, C.-C.; Hsu, Y.-C. Sulforaphane, a Dietary Isothiocyanate, Induces G 2/M Arrest in Cervical Cancer Cells through CyclinB1 Downregulation and GADD45β/CDC2 Association. Int. J. Mol. Sci. Artic. 2016, 17, 1530. [Google Scholar] [CrossRef] [Green Version]
- Mahata, S.; Bharti, A.C.; Shukla, S.; Tyagi, A.; Husain, S.A.; Das, B.C. Berberine modulates AP-1 activity to suppress HPV transcription and downstream signaling to induce growth arrest and apoptosis in cervical cancer cells. Mol. Cancer 2011, 10, 39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saha, S.K.; Khuda-Bukhsh, A.R. Berberine alters epigenetic modifications, disrupts microtubule network, and modulates HPV-18 E6-E7 oncoproteins by targeting p53 in cervical cancer cell HeLa: A mechanistic study including molecular docking. Eur. J. Pharmacol. 2015, 744, 132–146. [Google Scholar] [CrossRef] [PubMed]
- Guo, Y.; Xu, L.S.; Zhang, D.; Liao, Y.P.; Wang, H.P.; Lan, Z.H.; Guan, W.J.; Liu, C.Q. Betaine effects on morphology, proliferation, and p53-induced apoptosis of hela cervical carcinoma cells in vitro. Asian Pac. J. Cancer Prev. 2015, 16, 3195–3201. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yim, N.H.; Lee, J.H.; Cho, W.K.; Yang, M.C.; Kwak, D.H.; Ma, J.Y. Decursin and decursinol angelate from Angelica gigas Nakai induce apoptosis via induction of TRAIL expression on cervical cancer cells. Eur. J. Integr. Med. 2011, 3, e299–e307. [Google Scholar] [CrossRef]
- Jafri, A.; Siddiqui, S.; Rais, J.; Sultan Ahmad, M.; Kumar, S.; Jafar, T.; Afzal, M.; Arshad, M. Induction of Apoptosis by Piperine in Human Cervical Adenocarcinoma via Ros Mediated Mitochondrial Pathway and Caspase-3 Activation. EXCLI J. 2019, 18, 154–164. [Google Scholar] [CrossRef]
- Munagala, R.; Kausar, H.; Munjal, C.; Gupta, R.C. Withaferin a induces p53-dependent apoptosis by repression of HPV oncogenes and upregulation of tumor suppressor proteins in human cervical cancer cells. Carcinogenesis 2011, 32, 1697–1705. [Google Scholar] [CrossRef] [Green Version]
- Cai, S.; Adila, A.; Alimu, A.; Chen, Q.; Li, Y. Glycyrrhiza uralensis Fisch L. crude polysaccharides enhance mouse immunity and immune responses induced by HPV-DNA vaccine. Chin. J. Microbiol. Immunol. 2018, 38, 774–781. [Google Scholar]
- Yan, Q.; Li, Y.; Jiang, Z.; Sun, Y.; Zhu, L.; Ding, Z. Antiproliferation and apoptosis of human tumor cell lines by a lectin (AMML) of Astragalus mongholicus. Phytomedicine 2009, 16, 586–593. [Google Scholar] [CrossRef]
- Wang, P.; Li, J.C. Trichosanthin-induced specific changes of cytoskeleton configuration were associated with the decreased expression level of actin and tubulin genes in apoptotic Hela cells. Life Sci. 2007, 81, 1130–1140. [Google Scholar] [CrossRef]
- Mancinelli, L.; De Angelis, P.M.; Annulli, L.; Padovini, V.; Elgjo, K.; Gianfranceschi, G.L. A class of DNA-binding peptides from wheat bud causes growth inhibition, G2 cell cycle arrest and apoptosis induction in HeLa cells. Mol. Cancer 2009, 8, 55. [Google Scholar] [CrossRef] [Green Version]
- Bhutia, S.K.; Mallick, S.K.; Maiti, T.K. In vitro immunostimulatory properties of Abrus lectins derived peptides in tumor bearing mice. Phytomedicine 2009, 16, 776–782. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Tang, X.; Lu, Q.Y.; Zhang, Z.F.; Rao, J.; Le, A.D. Green tea extract and (-)-epigallocatechin-3-gallate inhibit hypoxia- and serum-induced HIF-1α protein accumulation and VEGF expression in human cervical carcinoma and hepatoma cells. Mol. Cancer Ther. 2006, 5, 1227–1238. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, D.; Basu, S.; Parija, L.; Rout, D.; Manna, S.; Dandapat, J.; Debata, P.R. Curcumin and Ellagic acid synergistically induce ROS generation, DNA damage, p53 accumulation and apoptosis in HeLa cervical carcinoma cells. Biomed. Pharmacother. 2016, 81, 31–37. [Google Scholar] [CrossRef]
- Ahn, W.S.; Yoo, J.; Huh, S.W.; Kim, C.K.; Lee, J.M.; Namkoong, S.E.; Bae, S.M.; Lee, I.P. Protective effects of green tea extracts (polyphenon E and EGCG) on human cervical lesions. Eur. J. Cancer Prev. 2003, 12, 383–390. [Google Scholar] [CrossRef] [PubMed]
- Garcia, F.A.R.; Cornelison, T.; Nuño, T.; Greenspan, D.L.; Byron, J.W.; Hsu, C.-H.; Alberts, D.S.; Chow, H.-H.S. Results of a phase II randomized, double-blind, placebo-controlled trial of Polyphenon E in women with persistent high-risk HPV infection and low-grade cervical intraepithelial neoplasia. Gynecol. Oncol. 2014, 132, 377–382. [Google Scholar] [CrossRef] [Green Version]
- Deepika, M.S.; Thangam, R.; Sheena, T.S.; Sasirekha, R.; Sivasubramanian, S.; Babu, M.D.; Jeganathan, K.; Thirumurugan, R. A novel rutin-fucoidan complex based phytotherapy for cervical cancer through achieving enhanced bioavailability and cancer cell apoptosis. Biomed. Pharmacother. 2019, 109, 1181–1195. [Google Scholar] [CrossRef]
- Kwon, S.B.; Kim, M.J.; Yang, J.M.; Lee, H.P.; Hong, J.T.; Jeong, H.S.; Kim, E.S.; Yoon, D.Y. Cudrania tricuspidata stem extract induces apoptosis via the extrinsic pathway in SiHa cervical cancer cells. PLoS ONE 2016, 11, e0150235. [Google Scholar] [CrossRef]
- Ghanbari, A.; Le Gresley, A.; Naughton, D.; Kuhnert, N.; Sirbu, D.; Ashrafi, G.H. Biological activities of Ficus carica latex for potential therapeutics in Human Papillomavirus (HPV) related cervical cancers. Sci. Rep. 2019, 9, 1013. [Google Scholar] [CrossRef] [Green Version]
- Khavari, A.; Bolhassani, A.; Alizadeh, F.; Bathaie, S.Z.; Balaram, P.; Agi, E.; Vahabpour, R. Chemo-immunotherapy using saffron and its ingredients followed by E7-NT (gp96) DNA vaccine generates different anti-tumor effects against tumors expressing the E7 protein of human papillomavirus. Arch. Virol. 2015, 160, 499–508. [Google Scholar] [CrossRef]
- Mahata, S.; Maru, S.; Shukla, S.; Pandey, A.; Mugesh, G.; Das, B.C.; Bharti, A.C. Anticancer property of Bryophyllum pinnata (Lam.) Oken. leaf on human cervical cancer cells. BMC Complement. Altern. Med. 2012, 12, 15. [Google Scholar] [CrossRef] [Green Version]
- Li, G.L.; Jiang, W.; Xia, Q.; Chen, S.H.; Ge, X.R.; Gui, S.Q.; Xu, C.J. HPV E6 downregulation and apoptosis induction of human cervical cancer cells by a novel lipid-soluble extract (PE) from Pinellia pedatisecta Schott in vitro. J. Ethnopharmacol. 2010, 132, 56–64. [Google Scholar] [CrossRef]
- Mahata, S.; Pandey, A.; Shukla, S.; Tyagi, A.; Husain, S.A.; Das, B.C.; Bharti, A.C. Anticancer activity of Phyllanthus emblica Linn. (Indian Gooseberry): Inhibition of transcription factor ap-1 and HPV gene expression in cervical cancer cells. Nutr. Cancer 2013, 65, 88–97. [Google Scholar] [CrossRef] [PubMed]
- Talwar, G.P.; Dar, S.A.; Rai, M.K.; Reddy, K.V.R.; Mitra, D.; Kulkarni, S.V.; Doncel, G.F.; Buck, C.B.; Schiller, J.T.; Muralidhar, S.; et al. A novel polyherbal microbicide with inhibitory effect on bacterial, fungal and viral genital pathogens. Int. J. Antimicrob. Agents 2008, 32, 180–185. [Google Scholar] [CrossRef] [PubMed]
- Basu, P.; Dutta, S.; Begum, R.; Mittal, S.; Dutta, P.D.; Bharti, A.C.; Panda, C.K.; Biswas, J.; Dey, B.; Talwar, G.P.; et al. Clearance of cervical human papillomavirus infection by topical application of curcumin and curcumin containing polyherbal cream: A phase II randomized controlled study. Asian Pac. J. Cancer Prev. 2013, 14, 5753–5759. [Google Scholar] [CrossRef] [Green Version]
- Talwar, G.P.; Sharma, R.; Singh, S.; Das, B.C.; Bharti, A.C.; Sharma, K.; Singh, P.; Atrey, N.; Gupta, J.C. BASANT, a Polyherbal Safe Microbicide Eliminates HPV-16 in Women with Early Cervical Intraepithelial Lesions. J. Cancer Ther. 2015, 6, 1163. [Google Scholar] [CrossRef] [Green Version]
- Shukla, S.; Bharti, A.C.; Hussain, S.; Mahata, S.; Hedau, S.; Kailash, U.; Kashyap, V.; Bhambhani, S.; Roy, M.; Batra, S.; et al. Elimination of high-risk human papillomavirus type HPV16 infection by ‘Praneem’ polyherbal tablet in women with early cervical intraepithelial lesions. J. Cancer Res. Clin. Oncol. 2009, 135, 1701–1709. [Google Scholar] [CrossRef] [PubMed]
- Talwar, G.P.; Raghuvanshi, P.; Mishra, R.; Banerjee, U.; Rattan, A.; Whaley, K.J.; Zeitlin, L.; Achilles, S.L.; Barré-Sinoussi, F.; David, A.; et al. Polyherbal formulations with wide spectrum antimicrobial activity against reproductive tract infections and sexually transmitted pathogens. Am. J. Reprod. Immunol. 2000, 43, 144–151. [Google Scholar] [CrossRef]
- Mamgain, S.; Sharma, P.; Pathak, R.K.; Baunthiyal, M. Computer aided screening of natural compounds targeting the E6 protein of HPV using molecular docking. Bioinformation 2015, 11, 236. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Jena, L.; Mohod, K.; Daf, S.; Varma, A.K. Virtual Screening for Potential Inhibitors of High-Risk Human Papillomavirus 16 E6 Protein. Interdiscip. Sci. Comput. Life Sci. 2015, 7, 136–142. [Google Scholar] [CrossRef]
- Nabati, F.; Moradi, M.; Mohabatkar, H. In silico analyzing the molecular interactions of plant-derived inhibitors against E6AP, p53, and c-Myc binding sites of HPV type 16 E6 oncoprotein. Mol. Biol. Res. Commun. 2020, 9, 71–82. [Google Scholar] [CrossRef]
- Pennisi, M.; Russo, G.; Ravalli, S.; Pappalardo, F. Combining agent based-models and virtual screening techniques to predict the best citrus-derived vaccine adjuvants against human papilloma virus. BMC Bioinform. 2017, 18, 87–97. [Google Scholar] [CrossRef]
- Buyel, J.F. Plants as sources of natural and recombinant anti-cancer agents. Biotechnol. Adv. 2018, 36, 506–520. [Google Scholar] [CrossRef] [PubMed]
- Obembe, O.O.; Popoola, J.O.; Leelavathi, S.; Reddy, S.V. Advances in plant molecular farming. Biotechnol. Adv. 2011, 29, 210–222. [Google Scholar] [CrossRef] [PubMed]
- Spiegel, H.; Boes, A.; Voepel, N.; Beiss, V.; Edgue, G.; Rademacher, T.; Sack, M.; Schillberg, S.; Reimann, A.; Fischer, R. Application of a Scalable Plant Transient Gene Expression Platform for Malaria Vaccine Development. Front. Plant Sci. 2015, 6, 1169. [Google Scholar] [CrossRef] [PubMed]
- Commandeur, U.; Twyman, R.M.; Fischer, R. The Biosafety of Molecular Farming in Plants; CABI: Wallingford, UK, 2003. [Google Scholar] [CrossRef]
- Buyel, J.F.; Stöger, E.; Bortesi, L. Targeted genome editing of plants and plant cells for biomanufacturing. Transgenic Res. 2021, 30, 401–426. [Google Scholar] [CrossRef]
- Zhu, H.; Li, C.; Gao, C. Applications of CRISPR–Cas in agriculture and plant biotechnology. Nat. Rev. Mol. Cell Biol. 2020, 21, 661–677. [Google Scholar] [CrossRef]
- Kim, B.G.; Yang, S.M.; Kim, S.Y.; Cha, M.N.; Ahn, J.H. Biosynthesis and production of glycosylated flavonoids in Escherichia coli: Current state and perspectives. Appl. Microbiol. Biotechnol. 2015, 99, 2979–2988. [Google Scholar] [CrossRef]
- Wang, H.; Yang, Y.; Lin, L.; Zhou, W.; Liu, M.; Cheng, K.; Wang, W. Engineering Saccharomyces cerevisiae with the deletion of endogenous glucosidases for the production of flavonoid glucosides. Microb. Cell Fact. 2016, 15, 134. [Google Scholar] [CrossRef] [Green Version]
- Menezes, J.C.; Orlikova, B.; Morceau, F.; Diederich, M. Natural and Synthetic Flavonoids: Structure–Activity Relationship and Chemotherapeutic Potential for the Treatment of Leukemia. Crit. Rev. Food Sci. Nutr. 2016, 56, S4–S28. [Google Scholar] [CrossRef]
- Birchfield, A.S.; McIntosh, C.A. Metabolic engineering and synthetic biology of plant natural products—A minireview. Curr. Plant Biol. 2020, 24, 100163. [Google Scholar] [CrossRef]
- Lannoo, N.; Vervecken, W.; Proost, P.; Rougé, P.; Van Damme, E.J.M. Expression of the nucleocytoplasmic tobacco lectin in the yeast Pichia pastoris. Protein Expr. Purif. 2007, 53, 275–282. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, C.; Felix, W.; Moreira, R.A.; Teixeira, J.A.; Domingues, L. Expression of frutalin, an α-d-galactose-binding jacalin-related lectin, in the yeast Pichia pastoris. Protein Expr. Purif. 2008, 60, 188–193. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Strasser, R. Plant protein glycosylation. Glycobiology 2016, 26, 926–939. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Endo, Y.; Tsurugi, K.; Franz, H. The Site of Action of the A-Chain of Mistletoe Lectin I on Eukaryotic Ribosomes the RNA IV-Glycosidase Activity of the Protein. FEBS Lett. 1988, 231, 378–380. [Google Scholar] [CrossRef] [Green Version]
- Buck, C.B.; Thompson, C.D.; Roberts, J.N.; Müller, M.; Lowy, D.R.; Schiller, J.T. Carrageenan is a potent inhibitor of papillomavirus infection. PLoS Pathog. 2006, 2, e69. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roberts, J.N.; Buck, C.B.; Thompson, C.D.; Kines, R.; Bernardo, M.; Choyke, P.L.; Lowy, D.R.; Schiller, J.T. Genital transmission of HPV in a mouse model is potentiated by nonoxynol-9 and inhibited by carrageenan. Nat. Med. 2007, 13, 857–861. [Google Scholar] [CrossRef] [Green Version]
- Costa, L.S.; Fidelis, G.P.; Cordeiro, S.L.; Oliveira, R.M.; Sabry, D.A.; Câmara, R.B.G.; Nobre, L.T.D.B.; Costa, M.S.S.P.; Almeida-Lima, J.; Farias, E.H.C.; et al. Biological activities of sulfated polysaccharides from tropical seaweeds. Biomed. Pharmacother. 2010, 64, 21–28. [Google Scholar] [CrossRef]
- Wang, W.; Wang, S.X.; Guan, H.S. The antiviral activities and mechanisms of marine polysaccharides: An overview. Mar. Drugs 2012, 10, 2795–2816. [Google Scholar] [CrossRef]
- Stevan, F.R.; Oliveira, M.B.; Bucchi, D.F.; Noseda; Iacomini, M.; Duarte, M.E. Cytotoxic effects against HeLa cells of polysaccharides from seaweeds. J. Submicrosc. Cytol. Pathol. 2001, 33, 477–484. [Google Scholar]
- Wang, S.-X.; Zhang, X.-S.; Guan, H.-S.; Wang, W. Potential Anti-HPV and Related Cancer Agents from Marine Resources: An Overview. Mar. Drugs 2014, 12, 2019–2035. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.H.; Ryu, B.; Je, J.Y.; Kim, S.K. Diethylaminoethyl chitosan induces apoptosis in HeLa cells via activation of caspase-3 and p53 expression. Carbohydr. Polym. 2011, 84, 571–578. [Google Scholar] [CrossRef]
- Román, J.J.M.; Del Campo, M.; Villar, J.; Paolini, F.; Curzio, G.; Venuti, A.; Jara, L.; Ferreira, J.; Murgas, P.; Lladser, A.; et al. Immunotherapeutic potential of mollusk hemocyanins in combination with human vaccine adjuvants in murine models of oral cancer. J. Immunol. Res. 2019, 2019, 7076942. [Google Scholar] [CrossRef]
- Karan, D.; Dubey, S.; Pirisi, L.; Nagel, A.; Pina, I.; Choo, Y.-M.; Hamann, M.T. The Marine Natural Product Manzamine A Inhibits Cervical Cancer by Targeting the SIX1 Protein. J. Nat. Prod. 2020, 83, 286–295. [Google Scholar] [CrossRef]
- Zhang, J.; Kang, J.; Liu, F.; Fan, C.; Li, H.; Chen, Q. Effect of the polypeptides from Meretrix meretrix Linnaeus on proliferation of cervical cancer Hela cells. J. Xiamen Univ. Nat. Sci. 2009, 5, 2019–2035. [Google Scholar]
- Lu, Y.; Zhang, B.Y.; Dong, Q.; Wang, B.L.; Sun, X.B. The Effects of Stichopus japonicus Acid Mucopolysaccharide on the Apoptosis of the Human Hepatocellular Carcinoma Cell Line HepG2. Am. J. Med. Sci. 2010, 339, 141–144. [Google Scholar] [CrossRef]
- Zhang, W.; Che, Q.; Tan, H.; Qi, X.; Li, J.; Li, D.; Gu, Q.; Zhu, T.; Liu, M. Marine Streptomyces sp. derived antimycin analogues suppress HeLa cells via depletion HPV E6/E7 mediated by ROS-dependent ubiquitin-proteasome system. Sci. Rep. 2017, 7, 42180. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, V.-T.; Suck Lee, J.; Qian, Z.-J.; Li, Y.-X.; Kim, K.-N.; Heo, S.-J.; Jeon, Y.-J.; Park, W.S.; Choi, I.-W.; Je, J.-Y.; et al. Gliotoxin Isolated from Marine Fungus Aspergillus sp. Induces Apoptosis of Human Cervical Cancer and Chondrosarcoma Cells. Mar. Drugs 2014, 12, 69–87. [Google Scholar] [CrossRef]
- Wijesekara, I.; Li, Y.-X.; Vo, T.-S.; Van Ta, Q.; Ngo, D.-H.; Kim, S.-K. Induction of apoptosis in human cervical carcinoma HeLa cells by neoechinulin A from marine-derived fungus Microsporum sp. Process Biochem. 2013, 48, 68–72. [Google Scholar] [CrossRef]
- Ghosh, T.; Chattopadhyay, K.; Marschall, M.; Karmakar, P.; Mandal, P.; Ray, B. REVIEW Focus on antivirally active sulfated polysaccharides: From structure-activity analysis to clinical evaluation. Glycobiology 2009, 19, 2–15. [Google Scholar] [CrossRef]
- Vo, T.-S.; Kim, S.-K. Potential Anti-HIV Agents from Marine Resources: An Overview. Mar. Drugs 2010, 8, 2871–2892. [Google Scholar] [CrossRef] [Green Version]
- Lembo, D.; Donalisio, M.; Rusnati, M.; Bugatti, A.; Cornaglia, M.; Cappello, P.; Giovarelli, M.; Oreste, P.; Landolfo, S. Sulfated K5 Escherichia coli Polysaccharide Derivatives as Wide-Range Inhibitors of Genital Types of Human Papillomavirus. Antimicrob. Agents Chemother. 2008, 52, 1374–1381. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hussaarts, K.G.A.M.; Hurkmans, D.P.; Oomen-de Hoop, E.; van Harten, L.J.; Berghuis, S.; van Alphen, R.J.; Spierings, L.E.A.; van Rossum-Schornagel, Q.C.; Vastbinder, M.B.; van Schaik, R.H.N.; et al. Impact of Curcumin (with or without Piperine) on the Pharmacokinetics of Tamoxifen. Cancers 2019, 11, 403. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lian, H.; Hui, Y.; Xiaoping, T.; Wei, T.; Jiyi, X.; Xiaolan, Y. Baicalein suppresses the proliferation of human cervical cancer cells via Notch 1/Hes signaling pathway. J. Cancer Res. Ther. 2019, 15, 1216. [Google Scholar] [CrossRef] [PubMed]
- Hu, J.; Wang, R.; Liu, Y.; Zhou, J.; Shen, K.; Dai, Y. Baicalein represses cervical cancer cell growth, cell cycle progression and promotes apoptosis via blocking akt/mtor pathway by the regulation of circhiat1/mir-19a-3p axis. Onco-Targets Ther. 2021, 14, 905. [Google Scholar] [CrossRef] [PubMed]
- Peng, Y.; Li, Q.; Li, K.; Zhao, H.; Han, Z.; Li, F.; Sun, M.; Zhang, Y. Antitumor activity of baicalein on the mice bearing U14 cervical cancer. Afr. J. Biotechnol. 2011, 10, 14169–14176. [Google Scholar] [CrossRef] [Green Version]
- Yang, H.; Xia, J.; Li, Y.; Cao, Y.; Tang, L.; Yu, X. Baicalein inhibits the invasion of human cervical cancer cells by inhibiting the hedgehog/Gli signaling pathway. Trop. J. Pharm. Res. 2020, 19, 115–120. [Google Scholar] [CrossRef]
- Bai, X.; Ma, Y.; Zhang, G. Butein suppresses cervical cancer growth through the PI3K/AKT/mTOR pathway. Oncol. Rep. 2015, 33, 3085–3092. [Google Scholar] [CrossRef] [Green Version]
- Tong, X.-Q.; Lv, W.; Luo, W.-Q.; Lu, W.-X.; Li, N. Treatment of human cervical cancer cells with butein leads to apoptosis and DNA damage. Oncol. Rep. 2016, 9, 11084–11089. [Google Scholar]
- Yang, P.-Y.; Hu, D.-N.; Kao, Y.-H.; Lin, I.-C.; Liu, F.-S. Butein induces apoptotic cell death of human cervical cancer cells. Oncol. Lett. 2018, 16, 6615–6623. [Google Scholar] [CrossRef]
- Zhang, D.; Sun, G.; Peng, L.; Tian, J.; Zhang, H. Calycosin inhibits viability, induces apoptosis, and suppresses invasion of cervical cancer cells by upregulating tumor suppressor miR-375. Arch. Biochem. Biophys. 2020, 691, 108478. [Google Scholar] [CrossRef]
- Khoo, B.Y.; Chua, S.L.; Balaram, P. Apoptotic effects of chrysin in human cancer cell lines. Int. J. Mol. Sci. 2010, 11, 2188–2199. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jung, J. Emerging Utilization of Chrysin Using Nanoscale Modification. J. Nanomater. 2016, 2016, 2894089. [Google Scholar] [CrossRef] [Green Version]
- Pratiwi, R.; Tunjung, W.A.S.; Rumiyati, R.; Amalia, A.R. Black Rice Bran Extracts and Fractions Containing Cyanidin 3-glucoside and Peonidin 3-glucoside Induce Apoptosis in Human Cervical Cancer Cells. Indones. J. Biotechnol. 2016, 20, 69–76. [Google Scholar] [CrossRef] [Green Version]
- Lee, K.; Hyun Lee, D.; Jung, Y.J.; Shin, S.Y.; Lee, Y.H. The natural flavone eupatorin induces cell cycle arrest at the G2/M phase and apoptosis in HeLa cells. Appl. Biol. Chem. 2016, 59, 193–199. [Google Scholar] [CrossRef]
- Abd Razak, N.; Yeap, S.K.; Alitheen, N.B.; Ho, W.Y.; Yong, C.Y.; Tan, S.W.; Tan, W.S.; Long, K. Eupatorin Suppressed Tumor Progression and Enhanced Immunity in a 4T1 Murine Breast Cancer Model. Integr. Cancer Ther. 2020, 19, 1534735420935625. [Google Scholar] [CrossRef]
- Wu, Z.; Zou, B.; Zhang, X.; Peng, X. Eupatilin regulates proliferation and cell cycle of cervical cancer by regulating hedgehog signalling pathway. Cell Biochem. Funct. 2020, 38, 428–435. [Google Scholar] [CrossRef]
- Tousi, M.S.; Sepehri, H.; Khoee, S.; Farimani, M.M.; Delphi, L.; Mansourizadeh, F. Evaluation of apoptotic effects of mPEG-b-PLGA coated iron oxide nanoparticles as a eupatorin carrier on DU-145 and LNCaP human prostate cancer cell lines. J. Pharm. Anal. 2021, 11, 108–121. [Google Scholar] [CrossRef]
- Chien, S.T.; Der Shi, M.; Lee, Y.C.; Te, C.C.; Shih, Y.W. Galangin, a novel dietary flavonoid, attenuates metastatic feature via PKC/ERK signaling pathway in TPA-treated liver cancer HepG2 cells. Cancer Cell Int. 2015, 15, 15. [Google Scholar] [CrossRef] [Green Version]
- Kumar, R.; Tiku, A.B. Galangin induces cell death by modulating the expression of glyoxalase-1 and Nrf-2 in HeLa cells. Chem. Biol. Interact. 2018, 279, 1–9. [Google Scholar] [CrossRef]
- Wang, K.L.; Yu, Y.C.; Hsia, S.M. Perspectives on the role of isoliquiritigenin in cancer. Cancers 2021, 13, 115. [Google Scholar] [CrossRef]
- Hirchaud, F.; Hermetet, F.; Ablise, M.; Fauconnet, S.; Vuitton, D.A.; Prétet, J.L.; Mougin, C. Isoliquiritigenin Induces Caspase-Dependent Apoptosis via Downregulation of HPV16 E6 Expression in Cervical Cancer Ca Ski Cells. Planta Med. 2013, 79, 1628–1635. [Google Scholar] [CrossRef] [PubMed]
- Wei, J.; Su, H.; Bi, Y.; Li, J.; Feng, L.; Sheng, W. Anti-proliferative effect of isorhamnetin on HeLa cells through inducing G2/M cell cycle arrest. Exp. Ther. Med. 2018, 15, 3917–3923. [Google Scholar] [CrossRef] [PubMed]
- Gong, G.; Guan, Y.Y.; Zhang, Z.L.; Rahman, K.; Wang, S.J.; Zhou, S.; Luan, X.; Zhang, H. Isorhamnetin: A review of pharmacological effects. Biomed. Pharmacother. 2020, 128, 110301. [Google Scholar] [CrossRef] [PubMed]
- Park, C.; Cha, H.J.; Choi, E.O.; Lee, H.; Hwang-Bo, H.; Ji, S.Y.; Kim, M.Y.; Kim, S.Y.; Hong, S.H.; Cheong, J.H.; et al. Isorhamnetin induces cell cycle arrest and apoptosis via reactive oxygen species-mediated AMP-activated protein kinase signaling pathway activation in human bladder cancer cells. Cancers 2019, 11, 1494. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cai, F.; Zhang, Y.; Li, J.; Huang, S.; Gao, R. Isorhamnetin inhibited the proliferation and metastasis of androgen-independent prostate cancer cells by targeting the mitochondrion-dependent intrinsic apoptotic and PI3K/Akt/mTOR pathway. Biosci. Rep. 2020, 40, BSR20192826. [Google Scholar] [CrossRef] [Green Version]
- Zhao, X.; Zhu, W.; Zhang, R.; Zhang, M.; Zhao, J.; Hou, J.; Zhang, W. Targeted juglone blocks the invasion and metastasis of HPV-positive cervical cancer cells. J. Pharmacol. Sci. 2019, 140, 211–217. [Google Scholar] [CrossRef]
- Lu, Z.; Chen, H.; Zheng, X.M.; Chen, M.L. Experimental study on the apoptosis of cervical cancer Hela cells induced by juglone through c-Jun N-terminal kinase/c-Jun pathway. Asian Pac. J. Trop. Med. 2017, 10, 572–575. [Google Scholar] [CrossRef]
- Zhang, Q.; Zhang, F.; Thakur, K.; Wang, J.; Wang, H.; Hu, F.; Zhang, J.G.; Wei, Z.J. Molecular mechanism of anti-cancerous potential of Morin extracted from mulberry in Hela cells. Food Chem. Toxicol. 2018, 112, 466–475. [Google Scholar] [CrossRef]
- Solairaja, S.; Andrabi, M.Q.; Dunna, N.R.; Venkatabalasubramanian, S. Overview of Morin and Its Complementary Role as an Adjuvant for Anticancer Agents. Nutr. Cancer 2021, 73, 927–942. [Google Scholar] [CrossRef]
- Yi, J.L.; Shi, S.; Shen, Y.L.; Wang, L.; Chen, H.Y.; Zhu, J.; Ding, Y. Myricetin and methyl eugenol combination enhances the anticancer activity, cell cycle arrest and apoptosis induction of cis-platin against HeLa cervical cancer cell lines. Int. J. Clin. Exp. Pathol. 2015, 8, 1116–1127. [Google Scholar]
- Devi, K.P.; Rajavel, T.; Habtemariam, S.; Nabavi, S.F.; Nabavi, S.M. Molecular mechanisms underlying anticancer effects of myricetin. Life Sci. 2015, 142, 19–25. [Google Scholar] [CrossRef] [PubMed]
- Taheri, Y.; Suleria, H.A.R.; Martins, N.; Sytar, O.; Beyatli, A.; Yeskaliyeva, B.; Seitimova, G.; Salehi, B.; Semwal, P.; Painuli, S.; et al. Myricetin bioactive effects: Moving from preclinical evidence to potential clinical applications. BMC Complement. Med. Ther. 2020, 20, 241. [Google Scholar] [CrossRef] [PubMed]
- Sheng, S.Q.; Yu, L.Y.; Zhou, X.W.; Pan, H.Y.; Hu, F.Y.; Liu, J.L. Paeonol prevents migration and invasion, and promotes apoptosis of cervical cancer cells by inhibiting 5-lipoxygenase. Mol. Med. Rep. 2021, 23, 401. [Google Scholar] [CrossRef] [PubMed]
- Du, J.; Song, D.; Li, J.; Li, Y.; Li, B.; Li, L. Paeonol triggers apoptosis in HeLa cervical cancer cells: The role of mitochondria-related caspase pathway. Psychopharmacol. 2021. [Google Scholar] [CrossRef]
- Hsiao, Y.H.; Hsieh, M.J.; Yang, S.F.; Chen, S.P.; Tsai, W.C.; Chen, P.N. Phloretin suppresses metastasis by targeting protease and inhibits cancer stemness and angiogenesis in human cervical cancer cells. Phytomedicine 2019, 62, 152964. [Google Scholar] [CrossRef]
- Lagoa, R.; Silva, J.; Rodrigues, J.R.; Bishayee, A. Advances in phytochemical delivery systems for improved anticancer activity. Biotechnol. Adv. 2020, 38, 107381. [Google Scholar] [CrossRef]
- Chatterjee, K.; Mukherjee, S.; Vanmanen, J.; Banerjee, P.; Fata, J.E. Dietary polyphenols, resveratrol and pterostilbene exhibit antitumor activity on an HPV E6-positive cervical cancer model: An in vitro and in vivo analysis. Front. Oncol. 2019, 9, 352. [Google Scholar] [CrossRef] [Green Version]
- Jia, L.; Hu, Y.; Yang, G.; Li, P. Puerarin suppresses cell growth and migration in HPV-positive cervical cancer cells by inhibiting the PI3K/mTOR signaling pathway. Exp. Ther. Med. 2019, 18, 543–549. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.; Chinnathambi, A.; Alharbi, S.A.; Veeraraghavan, V.P.; Mohan, S.K.; Zhang, G. Punicalagin promotes the apoptosis in human cervical cancer (ME-180) cells through mitochondrial pathway and by inhibiting the NF-kB signaling pathway. Saudi J. Biol. Sci. 2020, 27, 1100–1106. [Google Scholar] [CrossRef]
- Jing, Z.; Fei, W.; Zhou, J.; Zhang, L.; Chen, L.; Zhang, X.; Liang, X.; Xie, J.; Fang, Y.; Sui, X.; et al. Salvianolic acid B, a novel autophagy inducer, exerts antitumor activity as a single agent in colorectal cancer cells. Oncotarget 2016, 7, 61509–61519. [Google Scholar] [CrossRef]
- Mears, B.; Shiao, F.; Toney, N.; Chen, C.-M.; Choi, K.; Chow, C.; Navin, M.; Shiau, H. Salvianolic Acid B Inhibits Growth of Cervical Cancer Cells In Vitro via Induction of Apoptosis through the Extrinsic Pathway; University of Maryland: College Park, MD, USA, 2014. [Google Scholar]
- Tian, Q.; Wang, L.; Sun, X.; Zeng, F.; Pan, Q.; Xue, M. Scopoletin exerts anticancer effects on human cervical cancer cell lines by triggering apoptosis, cell cycle arrest, inhibition of cell invasion and PI3K/AKT signalling pathway. J. BUON 2019, 24, 997–1002. [Google Scholar]
- Yong, W.K.; Abd Malek, S.N. Xanthohumol induces growth inhibition and apoptosis in Ca Ski human cervical cancer cells. Evid.-Based Complement. Altern. Med. 2015, 2015, 921306. [Google Scholar] [CrossRef] [PubMed]
- Jiang, C.H.; Sun, T.L.; Xiang, D.X.; Wei, S.S.; Li, W.Q. Anticancer activity and mechanism of xanthohumol: A prenylated flavonoid from hops (Humulus lupulus L.). Front. Pharmacol. 2018, 9, 530. [Google Scholar] [CrossRef] [PubMed]
- Su, K.; Wang, C.F.; Zhang, Y.; Cai, Y.J.; Zhang, Y.Y.; Zhao, Q. The inhibitory effects of carnosic acid on cervical cancer cells growth by promoting apoptosis via ROS-regulated signaling pathway. Biomed. Pharmacother. 2016, 82, 180–191. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Li, Q.W.; Gao, D.W.; Han, Z.S.; Lu, W.Z. Antitumor and immunomodulating effects of polysaccharides isolated from Solanum nigrum Linne. Phyther. Res. 2009, 23, 1524–1530. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Database/Platform: | MEDLINE (PubMed) |
|---|---|
| Date coverage: | From 1995 to present |
| Library: | National Library of Medicine, National Center for Biotechnology Information |
| Date of last search: | 20 October 2021 |
| Limits: | In: “Article Title, Abstract, Keywords” Published: “All years” to “Present” Document type: “All” Subject Areas: All checked (default) |
| Search query: | HPV disease + natural compounds + bioactives + phytochemicals + combination treatments + therapy + infection + name of compounds/class of compounds |
| Number of hits: | 22 |
| Database/Platform: | Science Direct (Elsevier) |
| Date coverage: | From 1998 to present |
| Library: | Free access |
| Date of last search: | 20 October 2021 |
| Limits: | In: “Article Title, Abstract, Keywords” Published: “All years” to “Present” Document type: “All” Subject Areas: All checked (default) |
| Search query: | HPV disease + natural compounds + bioactives + phytochemicals + combination treatments + therapy + infection + name of compounds/class of compounds |
| Number of hits: | 2870 |
| Database/Platform: | Scopus (Elsevier) |
| Date coverage: | From 1995 to present |
| Library: | Free access |
| Date of last search: | 20 October 2021 |
| Limits: | In: “Article Title, Abstract, Keywords” Published: “All years” to “Present” Document type: “All” Subject Areas: All checked (default) |
| Search query: | HPV disease + natural compounds + bioactives + phytochemicals + combination treatments + therapy + infection + name of compounds/class of compounds |
| Number of hits: | 20 |
| Database/Platform: | Google Scholar |
| Date coverage: | From 1995 to present |
| Library: | Free access |
| Date of last search: | 20 October 2021 |
| Limits: | In: “Article Title, Abstract, Keywords” Published: “All years” to “Present” Document type: “All” Subject Areas: All checked (default) |
| Search query: | HPV disease + natural compounds + bioactives + phytochemicals + combination treatments + therapy + infection + name of compounds/class of compounds |
| Number of hits: | 1330 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Massa, S.; Pagliarello, R.; Paolini, F.; Venuti, A. Natural Bioactives: Back to the Future in the Fight against Human Papillomavirus? A Narrative Review. J. Clin. Med. 2022, 11, 1465. https://doi.org/10.3390/jcm11051465
Massa S, Pagliarello R, Paolini F, Venuti A. Natural Bioactives: Back to the Future in the Fight against Human Papillomavirus? A Narrative Review. Journal of Clinical Medicine. 2022; 11(5):1465. https://doi.org/10.3390/jcm11051465
Chicago/Turabian StyleMassa, Silvia, Riccardo Pagliarello, Francesca Paolini, and Aldo Venuti. 2022. "Natural Bioactives: Back to the Future in the Fight against Human Papillomavirus? A Narrative Review" Journal of Clinical Medicine 11, no. 5: 1465. https://doi.org/10.3390/jcm11051465
APA StyleMassa, S., Pagliarello, R., Paolini, F., & Venuti, A. (2022). Natural Bioactives: Back to the Future in the Fight against Human Papillomavirus? A Narrative Review. Journal of Clinical Medicine, 11(5), 1465. https://doi.org/10.3390/jcm11051465