
Enzyme-Linked Immunosorbent Assay: An Adaptable Methodology to Study SARS-CoV-2 Humoral and Cellular Immune Responses



Abstract
:1. Introduction
2. Detection of Immune Response to SARS-CoV-2
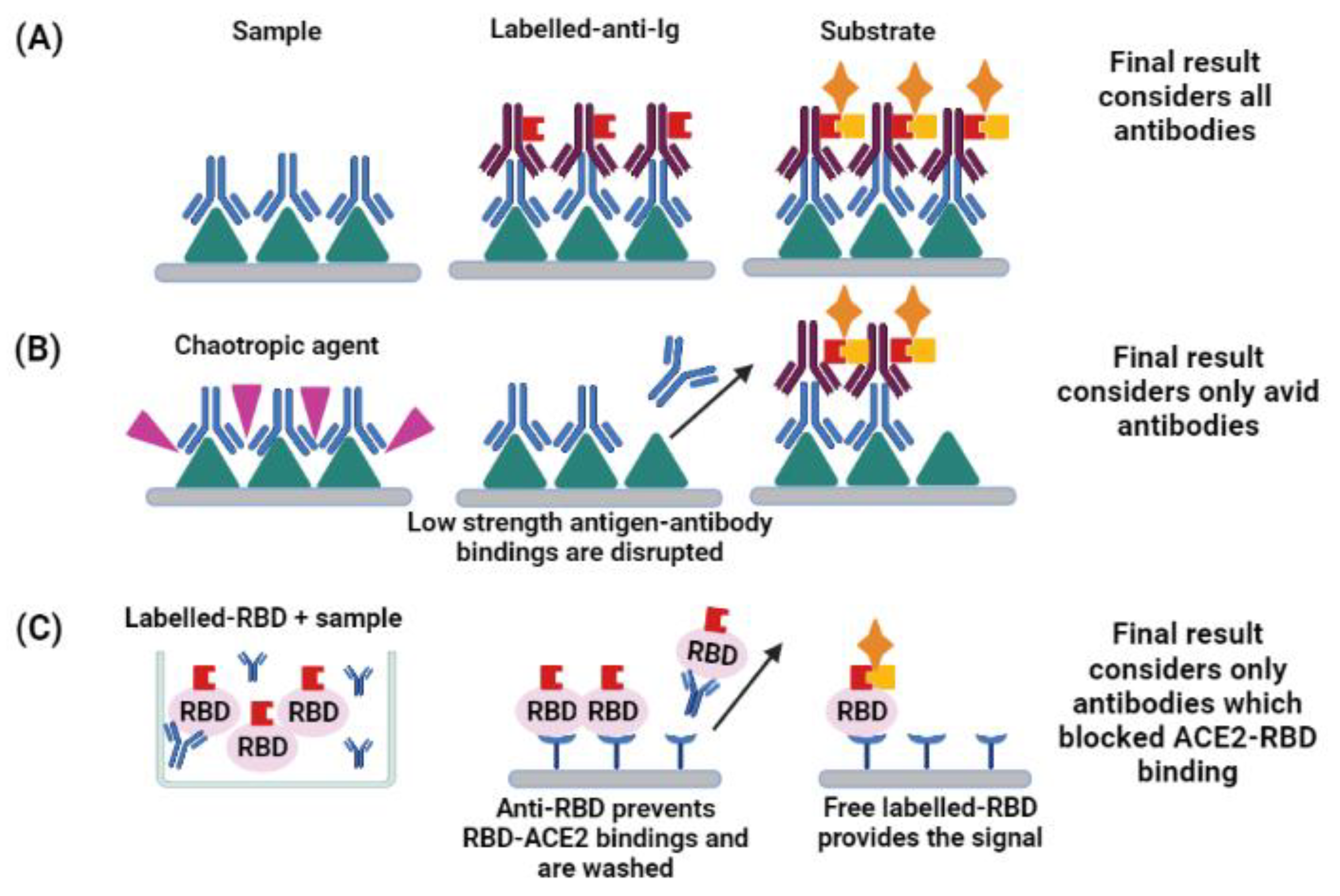
3. Enzyme-Linked Immunosorbent and Other Immunoassays
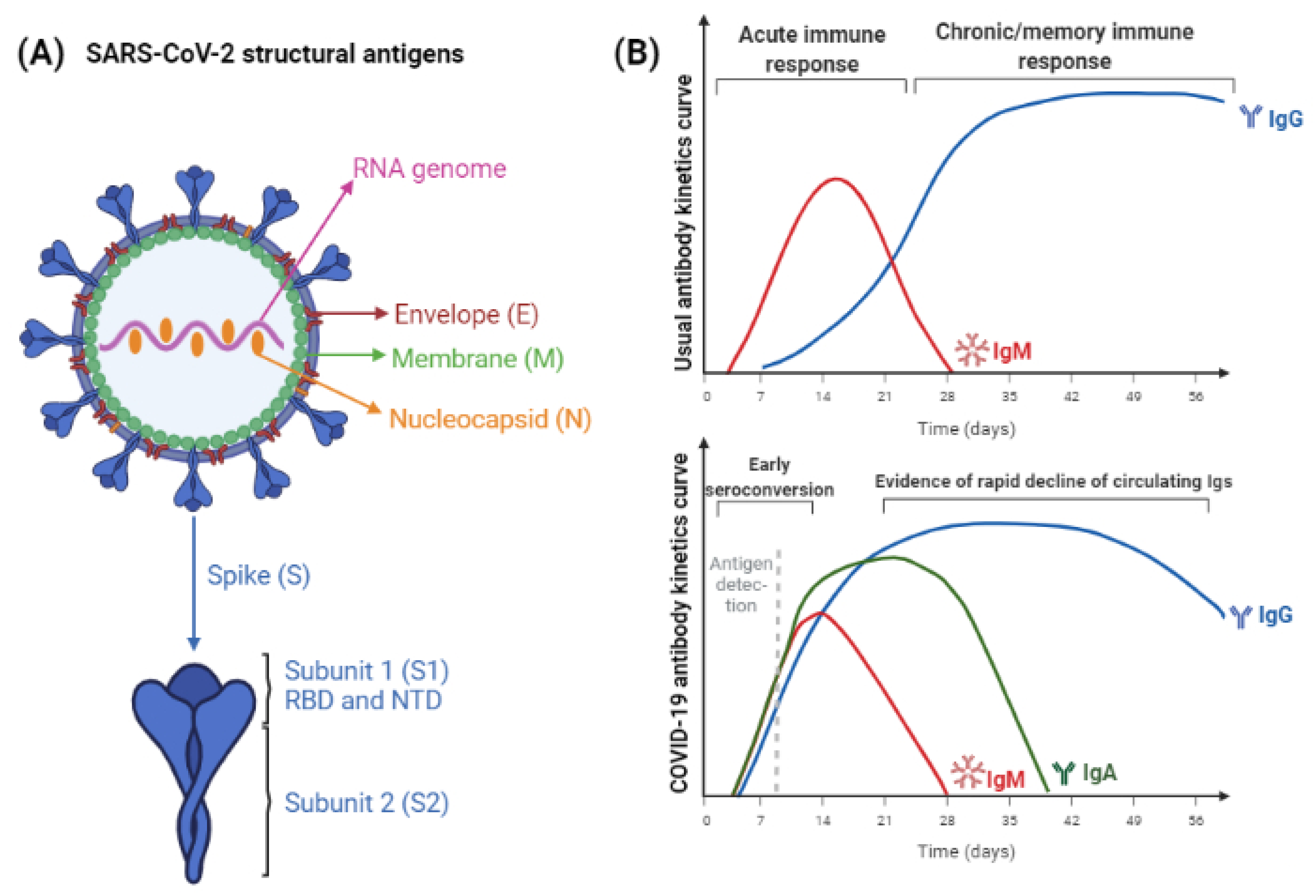
4. SARS-CoV-2 Antigens and Antibodies
5. Functionality of Antibodies
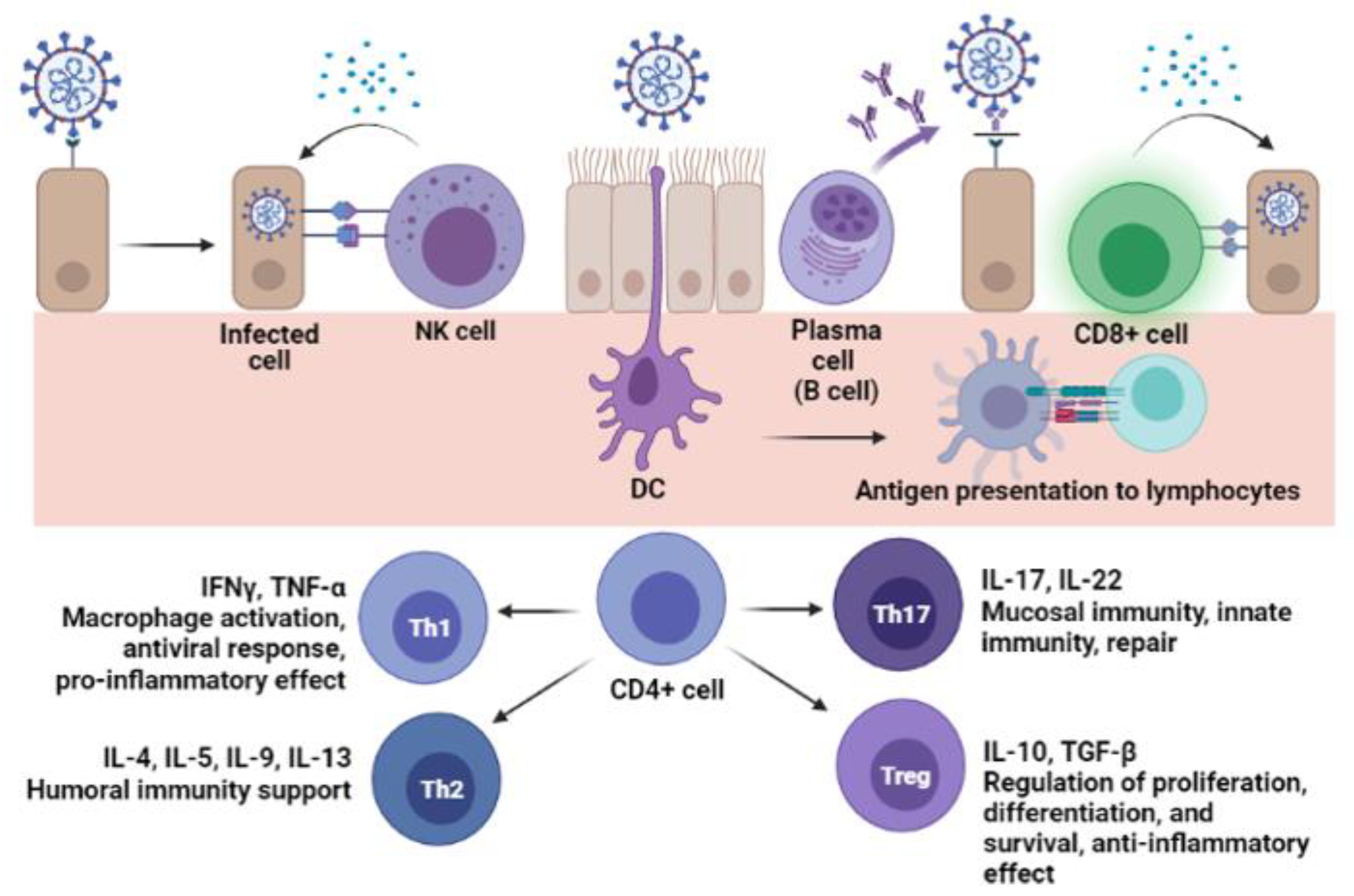
6. Cellular Immunity
7. Enzyme-Linked Techniques as Tools to Study Natural Infection and Vaccination
8. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Crowther, J.R. The ELISA Guidebook, 2nd ed.; Humana Press Inc.: Vienna, Austria, 2009; ISBN 978-1-60327-253-7. [Google Scholar]
- West, R.; Kobokovich, A.; Connell, N.; Gronvall, G.K. COVID-19 Antibody Tests: A Valuable Public Health Tool with Limited Relevance to Individuals. Trends Microbiol. 2021, 29, 214–223. [Google Scholar] [CrossRef] [PubMed]
- Maggi, E.; Canonica, G.W.; Moretta, L. COVID-19: Unanswered questions on immune response and pathogenesis. J. Allergy Clin. Immunol. 2020, 146, 18–22. [Google Scholar] [CrossRef] [PubMed]
- Assadiasl, S.; Fatahi, Y.; Zavvar, M.; Nicknam, M.H. COVID-19: Significance of antibodies. Hum. Antibodies 2020, 28, 287–297. [Google Scholar] [CrossRef] [PubMed]
- Ong, D.S.Y.; Fragkou, P.C.; Schweitzer, V.A.; Chemaly, R.F.; Moschopoulos, C.D.; Skevaki, C.; European Society of Clinical Microbiology and Infectious Diseases (ESCMID) Study Group for Respiratory Viruses (ESGREV). How to interpret and use COVID-19 serology and immunology tests. Clin. Microbiol. Infect. 2021, 27, 981–986. [Google Scholar] [CrossRef]
- Hamilton, R.G. The clinical immunology laboratory of the future. Clin. Chem. 1994, 40, 2186–2192. [Google Scholar] [CrossRef]
- Liu, G.; Rusling, J.F. COVID-19 Antibody Tests and Their Limitations. ACS Sensors 2021, 6, 593–612. [Google Scholar] [CrossRef]
- Amanat, F.; Stadlbauer, D.; Strohmeier, S.; Nguyen, T.H.O.; Chromikova, V.; McMahon, M.; Jiang, K.; Arunkumar, G.A.; Jurczyszak, D.; Polanco, J.; et al. A serological assay to detect SARS-CoV-2 seroconversion in humans. Nat. Med. 2020, 26, 1033–1036. [Google Scholar] [CrossRef]
- Moura, A.D.; Costa, H.H.M.; Correa, V.A.; Lima, A.K.S.; Lindoso, J.A.L.; De Gaspari, E.; Hong, M.A.; Cunha-Junior, J.P.; Prudencio, C.R. Assessment of avidity related to IgG subclasses in SARS-CoV-2 Brazilian infected patients. Sci. Rep. 2021, 11, 17642. [Google Scholar] [CrossRef]
- Allinson, J.L. Automated immunoassay equipment platforms for analytical support of pharmaceutical and biopharmaceutical development. Bioanalysis 2011, 3, 2803–2816. [Google Scholar] [CrossRef] [Green Version]
- Van Elslande, J.; Decru, B.; Jonckheere, S.; Van Wijngaerden, E.; Houben, E.; Vandecandelaere, P.; Indevuyst, C.; Depypere, M.; Desmet, S.; André, E.; et al. Antibody response against SARS-CoV-2 spike protein and nucleoprotein evaluated by four automated immunoassays and three ELISAs. Clin. Microbiol. Infect. 2020, 26, 1557.e1–1557.e7. [Google Scholar] [CrossRef]
- Byrum, J.R.; Waltari, E.; Janson, O.; Guo, S.-M.; Folkesson, J.; Chhun, B.B.; Vinden, J.; Ivanov, I.E.; Forst, M.L.; Li, H.; et al. multiSero: Open multiplex-ELISA platform for analyzing antibody responses to SARS-CoV-2 infection. MedRxiv 2021. [Google Scholar] [CrossRef]
- Krüttgen, A.; Cornelissen, C.G.; Dreher, M.; Hornef, M.; Imöl, M.; Kleines, M. Comparison of four new commercial serologic assays for determination of SARS-CoV-2 IgG. J. Clin. Virol. 2020, 128, 104394. [Google Scholar] [CrossRef] [PubMed]
- Olbrich, L.; Castelletti, N.; Schälte, Y.; Garí, M.; Pütz, P.; Bakuli, A.; Pritsch, M.; Kroidl, I.; Saathoff, E.; Guggenbuehl Noller, J.M.; et al. Head-to-head evaluation of seven different seroassays including direct viral neutralisation in a representative cohort for SARS-CoV-2. J. Gen. Virol. 2021, 102, 001653. [Google Scholar] [CrossRef]
- Bastos, L.M.; Tavaziva, G.; Abidi, S.K.; Campbell, J.R.; Haraoui, L.P.; Johnston, J.C.; Lan, Z.; Law, S.; MacLean, E.; Trajman, A.; et al. Diagnostic accuracy of serological tests for covid-19: Systematic review and meta-analysis. BMJ 2020, 370, m2516. [Google Scholar] [CrossRef] [PubMed]
- Saker, K.; Escuret, V.; Pitiot, V.; Massardier-Pilonchéry, M.; Paul, S.; Mokdad, B.; Langlois-Jaques, C.; Rabilloud, M.; Goncalves, D.; Fabien, N.; et al. Evaluation of commercial anti-SARS-CoV-2 antibody assays and comparison of standardized titers in vaccinated health care workers. J. Clin. Microbiol. 2021, 60, e01746-21. [Google Scholar] [CrossRef]
- Van Elslande, J.; Houben, E.; Depypere, M.; Brackenier, A.; Desmet, S.; André, E.; van Ranst, M.; Lagrou, K.; Vermeersch, P. Diagnostic performance of seven rapid IgG/IgM antibody tests and the Euroimmun IgA/IgG ELISA in COVID-19 patients. Clin. Microbiol. Infect. 2020, 26, 1082–1087. [Google Scholar] [CrossRef]
- Perez-Saez, J.; Zaballa, M.E.; Yerly, S.; Andrey, D.O.; Meyer, B.; Eckerlle, I.; Balavoine, J.-F.; Chappuis, F.; Pittet, D.; Trono, D.; et al. Persistence of anti-SARS-CoV-2 antibodies: Immunoassay heterogeneity and implications for serosurveillance. Clin. Microbiol. Infect. 2021, 27, 1695.e7–1695.e12. [Google Scholar] [CrossRef]
- Dowlatshahi, S.; Shabani, E.; Abdekhodaie, M.J. Serological assays and host antibody detection in coronavirus-related disease diagnosis. Arch. Virol. 2021, 166, 715–731. [Google Scholar] [CrossRef]
- Dai, L.; Gao, G.F. Viral targets for vaccines against COVID-19. Nat. Rev. Immunol. 2021, 21, 73–82. [Google Scholar] [CrossRef]
- Mesquita Júnior, D.; Araújo, J.A.P.; Catelan, T.T.T.; Souza, A.W.S.; Cruvinel, W.M.; Andrade, L.E.C.; Silva, N.P. Immune System Part II—Basis of the immunological response mediated by T and B lymphocytes. Brazilian J. Rheumatol. 2010, 50, 552–580. [Google Scholar] [CrossRef]
- Cancrini, G.; Iori, A. Traditional and innovative diagnostic tools: When and why should be applied. Parassitologia 2004, 46, 173–176. [Google Scholar] [PubMed]
- Ilkhani, H.; Hedayat, N.; Farhad, S. Novel approaches for rapid detection of COVID-19 during the pandemic: A review. Anal. Biochem. 2021, 634, 114362. [Google Scholar] [CrossRef]
- Matsuda, E.M.; de Campos, I.B.; de Oliveira, I.P.; Colpas, D.R.; dos Santos Carmo, A.M.; Brígido, L.F.M. Field evaluation of COVID-19 antigen tests versus RNA based detection: Potential lower sensitivity compensated by immediate results, technical simplicity, and low cost. J. Med. Virol. 2021, 93, 4405–4410. [Google Scholar] [CrossRef] [PubMed]
- Khan, J.; Al Asoom, L.I.; Khan, M.; Chakrabartty, I.; Dandoti, S.; Rudrapal, M.; Zothantluanga, J.H. Evolution of RNA viruses from SARS to SARS-CoV-2 and diagnostic techniques for COVID-19: A review. J. Basic Appl. Sci. 2021, 10, 60. [Google Scholar] [CrossRef] [PubMed]
- Lumley, S.F.; Wei, J.; O’Donnel, D.; Stoesser, N.E.; Matthews, P.C.; Howarth, A.; Hatch, S.B.; Marsden, B.D.; Cox, S.; James, T.; et al. The Duration, Dynamics, and Determinants of Severe Acute Respiratory Syndrome Coronavirus 2 (SARS-CoV-2) Antibody Responses in Individual Healthcare Workers. Clin. Infect. Dis. 2021, 73, e699–e709. [Google Scholar] [CrossRef] [PubMed]
- Villalta, D.; Moratto, A.; Salgarolo, V.; Da Re, M.; Giacomello, R.; Malipiero, G. New-Generation Quantitative Immunoassays for SARS-CoV-2 Antibody Detection: Need for Harmonization. Ann. Lab. Med. 2022, 42, 113–116. [Google Scholar] [CrossRef]
- Abusrewill, Z.; Alhudiri, I.M.; Kaal, H.H.; El Meshri, S.E.; Ebrahim, F.O.; Dalyoum, T.; Efrefer, A.A.; Ibrahim, K.; Elfghi, M.B.; Abusrewill, S.; et al. Time scale performance of rapid antigen testing for SARS-CoV-2: Evaluation of 10 rapid antigen assays. J. Med. Virol. 2021, 93, 6512–6518. [Google Scholar] [CrossRef]
- Girt, G.C.; Lakshminarayanan, A.; Huo, J.; Dormon, J.; Norman, C.; Afrough, B.; Harding, A.; James, W.; Owens, R.J.; Naismith, J.H. The use of nanobodies in a sensitive ELISA test for SARS-CoV-2 Spike 1 protein. R. Soc. Open Sci. 2021, 8, 211016. [Google Scholar] [CrossRef]
- Van der Moeren, N.; Zwart, V.F.; Goderski, G.; Rijkers, G.T.; van den Bijllaardt, W.; Veenemans, J.; Kluytmans, J.A.J.W.; Pas, S.D.; Meijer, A.; Verweij, J.J.; et al. Performance of the Diasorin SARS-CoV-2 antigen detection assay on the LIAISON XL. J. Clin. Virol. 2021, 141, 104909. [Google Scholar] [CrossRef]
- Amanat, F.; Thapa, M.; Lei, T.; Ahmed, S.M.S.; Adelsberg, D.C.; Carreño, J.M.; Strohmeier, S.; Schmitz, A.J.; Zafar, S.; Zhou, J.Q.; et al. SARS-CoV-2 mRNA vaccination induces functionally diverse antibodies to NTD, RBD, and S2. Cell 2021, 184, 3936–3948. [Google Scholar] [CrossRef]
- Krut, V.G.; Astrakhantseva, I.V.; Chuvpilo, S.A.; Efimov, G.A.; Ambaryan, S.G.; Drutskaya, M.S.; Nedospasov, S.A. Antibodies to the N-Terminal Domain of Angiotensin-Converting Enzyme (ACE2) That Block Its Interaction with SARS-CoV-2 S Protein. Dokl. Biochem. Biophys. 2021, 1–4. [Google Scholar] [CrossRef]
- Amjadi, M.F.; Adyniec, R.R.; Gupta, S.; Bashar, S.J.; Mergaert, A.M.; Braun, K.M.; Moreno, G.K.; O’Connor, D.H.; Friedrich, T.C.; Safdar, N.; et al. Anti-membrane and anti-spike antibodies are long-lasting and together discriminate between past COVID-19 infection and vaccination. medRxiv 2021. [Google Scholar] [CrossRef]
- Zhao, J.; Yuan, Q.; Wang, H.; Liu, W.; Liao, X.; Su, Y.; Wang, X.; Yuan, J.; Li, T.; Li, J.; et al. Antibody Responses to SARS-CoV-2 in Patients with Novel Coronavirus Disease 2019. Clin. Infect. Dis. 2020, 71, 2027–2034. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.; Wang, Y.; Li, X.; Ren, L.; Zhao, J.; Hu, Y.; Zhang, L.; Fan, G.; Xu, J.; Gu, X.; et al. Clinical features of patients infected with 2019 novel coronavirus in Wuhan, China. Lancet 2020, 395, 497–506. [Google Scholar] [CrossRef] [Green Version]
- Ma, H.; Zeng, W.; He, H.; Zhao, D.; Jiang, D.; Zhou, P.; Cheng, L.; Li, Y.; Ma, X.; Jin, T. Serum IgA, IgM, and IgG responses in COVID-19. Cell. Mol. Immunol. 2020, 17, 773–775. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Yu, X.; Gao, C.; Zhang, L.; Zhai, H.; Hu, Y.; Liu, E.; Wang, Q.; Gao, Y.; Wei, D.; et al. Characterization of antibody responses to SARS-CoV-2 in convalescent COVID-19 patients. J. Med. Virol. 2021, 93, 2227–2233. [Google Scholar] [CrossRef] [PubMed]
- Dan, J.M.; Mateus, J.; Kato, Y.; Hastie, K.M.; Yu, E.D.; Faliti, C.E.; Grifoni, A.; Ramirez, S.I.; Haupt, S.; Frazier, A.; et al. Immunological memory to SARS-CoV-2 assessed for up to 8 months after infection. Science 2021, 371, eabf4063. [Google Scholar] [CrossRef]
- Swadzba, J.; Anyszek, T.; Panek, A.; Martin, E. Anti-Spike SARS-CoV-2 IgG Assessment with a Commercial Assay during a 4-Month Course after COVID-19 Vaccination. Vaccines 2021, 9, 1367. [Google Scholar] [CrossRef] [PubMed]
- Ciabattini, A.; Pastore, G.; Fiorino, F.; Polvere, J.; Lucchesi, S.; Pettini, E.; Stefano, A.; Rancan, I.; Durante, M.; Miscia, M.; et al. Evidence of SARS-CoV-2-Specific Memory B Cells Six Months After Vaccination With the BNT162b2 mRNA Vaccine. Front. Immunol. 2021, 12, 740708. [Google Scholar] [CrossRef]
- Atyeo, C.; Fischinger, S.; Zohar, T.; Slein, M.D.; Burke, J.; Loos, C.; McCulloch, D.J.; Newman, K.L.; Wolf, C.; Yu, J.; et al. Distinct Early Serological Signatures Track with SARS-CoV-2 Survival. Immunity 2020, 53, 524–532.E4. [Google Scholar] [CrossRef]
- Portilho, A.I.; Silva, V.O.; Ahagon, C.M.; Matsuda, E.M.; de Oliveira, E.L.; da Silveira, E.P.R.; Lima, A.K.S.; Lindoso, J.A.L.; Campos, I.B.; Hong, M.A.; et al. Humoral response to spike S1 and S2 and nucleocapsid proteins on microarray after SARS-CoV-2 infection. J. Med. Virol. 2021, 94, 178–185. [Google Scholar] [CrossRef] [PubMed]
- Hashem, A.; Algaissi, A.; Almahboub, S.; Alfaleh, M.; Abujamel, T.; Alamri, S.; Alluhaybi, K.; Hobani, H.; AlHarbi, R.; Alsulaiman, R.; et al. Early Humoral Response Correlates with Disease Severity and Outcomes in COVID-19 Patients. Viruses 2020, 12, 1390. [Google Scholar] [CrossRef] [PubMed]
- Silva, J.; Lucas, C.; Sundaram, M.; Israelow, B.; Wong, P.; Klein, J.; Lu, P.; Venkataraman, A.; Liu, F.; Mao, T.; et al. Saliva viral load is a dynamic unifying correlate of COVID-19 severity and mortality. medRxiv 2021. [Google Scholar] [CrossRef]
- Mestecky, J.; Mcghee, J.R. Immunoglobulin A (IgA): Molecular and Cellular Interactions Involved in IgA Biosynthesis and Immune Response. Adv. Immunol. 1987, 40, 153–245. [Google Scholar] [CrossRef]
- Kerr, M.A. The structure and function of human IgA. Biochem. J. 1990, 271, 285–296. [Google Scholar] [CrossRef] [Green Version]
- Hiemstra, P.S.; Gorter, A.; Stuurman, M.E.; van Es, L.A.; Daha, M.R. Activation of the alternative pathway of complement by human serum IgA. Adv. Exp. Med. Biol. 1987, 17, 321–326. [Google Scholar] [CrossRef]
- Sterlin, D.; Mathian, A.; Miyara, M.; Mohr, A.; Anna, F.; Claër, L.; Quentric, P.; Fadlallah, J.; Ghillani, P.; Gunn, C.; et al. IgA dominates the early neutralizing antibody response to SARS-CoV-2. Sci. Transl. Med. 2021, 13, eabd2223. [Google Scholar] [CrossRef]
- Fedele, G.; Russo, G.; Schiavoni, I.; Leone, P.; Olivetta, E.; Perri, E.; Zingaropoli, M.A.; Ciardi, M.R.; Pasculli, P.; Mastroianni, C.M.; et al. Early IgG / IgA response in hospitalized COVID-19 patients is associated with a less severe disease. Diagn. Microbiol. Infect. Dis. 2022, 102, 115586. [Google Scholar] [CrossRef]
- Vossenkämper, A.; Blair, P.A.; Safinia, N.; Fraser, L.D.; Das, L.; Sanders, T.J.; Stagg, A.J.; Sanderson, J.D.; Taylor, K.; Chang, F.; et al. A role for gut-associated lymphoid tissue in shaping the human B cell repertoire. J. Exp. Med. 2013, 210, 1665–1674. [Google Scholar] [CrossRef] [Green Version]
- Victora, G.D.; Nussenzweig, M.C. Germinal Centers. Ann. Rev. Immunol. 2012, 30, 429–457. [Google Scholar] [CrossRef]
- Eisen, H.N. Affinity enhancement of antibodies: How low-affinity antibodies produced early in immune responses are followed by high-affinity antibodies later and in memory B-cell responses. Cancer Immunol. Res. 2014, 2, 381–392. [Google Scholar] [CrossRef] [Green Version]
- Correa, V.A.; Rodrigues, T.S.; Portilho, A.I.; Trzewikoswki de Lima, G.; De Gaspari, E. Modified ELISA for antibody avidity evaluation: The need for standardization. Biomed. J. 2021, 44, 433–438. [Google Scholar] [CrossRef] [PubMed]
- Bauer, G. The potential significance of high avidity immunoglobulin G (IgG) for protective immunity towards SARS-CoV-2. Int. J. Infect. Dis. 2021, 106, 61–64. [Google Scholar] [CrossRef]
- Iwasaki, A.; Yang, Y. The potential danger of suboptimal antibody responses in COVID-19. Nat. Rev. Immunol. 2020, 20, 339–341. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klasse, P.J.; Sattentau, Q.J. Occupancy and mechanism in antibody-mediated neutralization of animal viruses. J. Gen. Virol. 2002, 83, 2091–2108. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.; Wang, J.; Li, Q.; Hu, H.; Lu, J.; Chen, Z. Advances in Neutralization Assays for SARS-CoV-2. Scand. J. Immunol. 2021, 94, 1–15. [Google Scholar] [CrossRef]
- Tan, C.W.; Chia, W.N.; Qin, X.; Liu, P.; Chen, M.I.C.; Tiu, C.; Hu, Z.; Chen, V.C.W.; Young, B.E.; Sia, W.R.; et al. A SARS-CoV-2 surrogate virus neutralization test based on antibody-mediated blockage of ACE2–spike protein–protein interaction. Nat. Biotechnol. 2020, 38, 1073–1078. [Google Scholar] [CrossRef]
- Yin, Q.; Zhang, Y.; Lian, L.; Qu, Y.; Wu, W.; Chen, Z.; Pei, R.; Chen, T.; Sun, L.; Li, C.; et al. Chemiluminescence Immunoassay Based Serological Immunoassays for Detection of SARS-CoV-2 Neutralizing Antibodies in COVID-19 Convalescent Patients and Vaccinated Population. Viruses 2021, 13, 1508. [Google Scholar] [CrossRef]
- Neumann, F.; Rose, R.; Römpke, J.; Grobe, O.; Lorentz, T.; Fickenscher, H.; Krumbholz, A. Development of sars-cov-2 specific IgG and virus-neutralizing antibodies after infection with variants of concern or vaccination. Vaccines 2021, 9, 700. [Google Scholar] [CrossRef]
- von Rhein, C.; Scholz, T.; Henss, L.; Kronstein-Wiedemann, R.; Schwarz, T.; Rodionov, R.N.; Corman, V.M.; Tonn, T.; Schnierle, B.S. Comparison of potency assays to assess SARS-CoV-2 neutralizing antibody capacity in COVID-19 convalescent plasma. J. Virol. Methods 2021, 288, 114031. [Google Scholar] [CrossRef]
- Meyer, B.; Reimerink, J.; Torriani, G.; Brouwer, F.; Godeke, G.J.; Yerly, S.; Hoogerwerf, M.; Vuilleumier, N.; Kaiser, L.; Eckerle, I.; et al. Validation and clinical evaluation of a SARS-CoV-2 surrogate virus neutralisation test (sVNT). Emerg. Microbes Infect. 2020, 9, 2394–2403. [Google Scholar] [CrossRef] [PubMed]
- Cruvinel, W.M.; Júnior, D.M.; Araújo, J.A.P.; Catelan, T.T.T.; de Souza, A.W.S.; da Silva, N.P.; Andrade, L.E.C. Immune system—Part I fundamentals of innate immunity with emphasis on molecular and cellular mechanisms of inflammatory response. Rev. Bras. Reumatol. 2010, 50, 443–461. [Google Scholar] [CrossRef]
- Hogrefe, W.R. Biomarkers and assessment of vaccine responses. Biomarkers 2005, 10 (Suppl. 1), 50–57. [Google Scholar] [CrossRef] [PubMed]
- Freer, G.; Rindi, L. Intracellular cytokine detection by fluorescence-activated flow cytometry: Basic principles and recent advances. Methods 2013, 61, 30–38. [Google Scholar] [CrossRef] [PubMed]
- Czerkinsky, C.C.; Svennerholm, M.S. Ganglioside GM1 enzyme-linked immunospot assay for simple identification of heat-labile enterotoxin-producing Escherichia coli. J. Clin. Microbiol. 1983, 17, 965–969. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Czerkinsky, C.C.; Nilsson, L.A.; Nygren, H.; Ouchterlony, O.; Tarkowski, A. A solid-phase enzyme-linked immunospot (ELISPOT) assay for enumeration of specific antibody-secreting cells. J. Immunol. Methods 1983, 65, 109–121. [Google Scholar] [CrossRef]
- Janetzki, S. ELISpot for Rookies (and Experts Too), 1st ed.; Springer Nature: Cham, Switzerland, 2016; ISBN 978-3-319-45293-7. [Google Scholar]
- Lima-Junior, J.; Morgado, F.; Conceição-Silva, F. How Can Elispot Add Information to Improve Knowledge on Tropical Diseases? Cells 2017, 6, 31. [Google Scholar] [CrossRef]
- Tripathy, A.S.; Vishwakarma, S.; Trimbake, D.; Gurav, Y.K.; Potdar, V.A.; Mokashi, N.D.; Patsute, S.D.; Kaushal, H.; Choudhary, M.L.; Tilekar, B.N.; et al. Pro-inflammatory CXCL-10, TNF-α, IL-1β, and IL-6: Biomarkers of SARS-CoV-2 infection. Arch. Virol. 2021, 166, 3301–3310. [Google Scholar] [CrossRef]
- Liu, Y.; Zhang, C.; Huang, F.; Yang, Y.; Wang, F.; Yuan, J.; Zhang, Z.; Qin, Y.; Li, X.; Zhao, D.; et al. Elevated plasma levels of selective cytokines in COVID-19 patients reflect viral load and lung injury. Nat. Sci. Rev. 2020, 7, 1003–1011. [Google Scholar] [CrossRef] [Green Version]
- Bösmüller, H.; Traxler, S.; Bitzer, M.; Häberle, H.; Raiser, W.; Nann, D.; Frauenfeld, L.; Vogelsberg, A.; Klingel, K.; Fend, F.; et al. The evolution of pulmonary pathology in fatal COVID-19 disease: An autopsy study with clinical correlation. Virchows Arch. 2020, 477, 349–357. [Google Scholar] [CrossRef]
- Hassaniazad, M.; Vahedi, M.S.; Samimagham, H.R.; Gharibzadeh, A.; Beyranvand, S.; Abassi, H.; Nikpoor, A.R. Improvement of clinical outcome, laboratory findings and inflammatory cytokines levels using plasmapheresis therapy in severe COVID-19 cases. Respir. Med. 2021, 189, 106669. [Google Scholar] [CrossRef] [PubMed]
- Del Valle, D.M.; Kim-Schulze, S.; Huang, H.H.; Beckmann, N.D.; Nirenberg, S.; Wang, B.; Lavin, Y.; Swartz, T.H.; Madduri, D.; Stock, A.; et al. An inflammatory cytokine signature predicts COVID-19 severity and survival. Nat. Med. 2020, 26, 1636–1646. [Google Scholar] [CrossRef] [PubMed]
- Angioni, R.; Sánchez-Rodriguéz, R.; Munari, F.; Bertoldi, N.; Arcidiacono, D.; Cavinato, S.; Marturano, D.; Zaramella, A.; Realdon, S.; Cattelan, A.; et al. Age-severity matched cytokine profiling reveals specific signatures in Covid-19 patients. Cell Death Res. 2020, 11, 957. [Google Scholar] [CrossRef] [PubMed]
- Batista, C.M.; Foti, L. Anti-SARS-CoV-2 and anti-cytokine storm neutralizing antibody therapies against COVID-19: Update, challenges, and perspectives. Int. Immunopharmacol. 2021, 99, 108036. [Google Scholar] [CrossRef] [PubMed]
- Mortaz, E.; Tabarsi, P.; Varahram, M.; Folkerts, G.; Adcock, I.M. The Immune Response and Immunopathology of COVID-19. Front. Immunol. 2020, 11, 1–9. [Google Scholar] [CrossRef]
- Li, G.; Fan, Y.; Lai, Y.; Han, T.; Li, Z.; Zhou, P.; Pan, P.; Wang, W.; Hu, D.; Liu, X.; et al. Coronavirus infections and immune responses. J. Med. Virol. 2020, 92, 424–432. [Google Scholar] [CrossRef]
- Foulds, K.E.; Wu, C.Y.; Seder, R.A. Th1 memory: Implications for vaccine development. Immunol. Rev. 2006, 211, 58–66. [Google Scholar] [CrossRef]
- Schwarzkopf, S.; Krawczyk, A.; Knop, D.; Klump, H.; Heinold, A.; Heinemann, F.M.; Thümmler, L.; Temme, C.; Breyer, M.; Witzke, O.; et al. Cellular immunity in COVID-19 convalescents with PCR-confirmed infection but with undetectable SARS-CoV-2-specific IgG. Emerg. Infect. Dis. 2021, 27, 122–129. [Google Scholar] [CrossRef]
- Li, J.; Hui, A.; Zhang, X.; Yang, Y.; Tang, R.; Ye, H.; Ji, R.; Lin, M.; Zhu, Z.; Türeci, Ö.; et al. Safety and immunogenicity of the SARS-CoV-2 BNT162b1 mRNA vaccine in younger and older Chinese adults: A randomized, placebo-controlled, double-blind phase 1 study. Nat. Med. 2021, 27, 1062–1070. [Google Scholar] [CrossRef]
- Folegatti, P.M.; Ewer, K.J.; Aley, P.K.; Angus, B.; Becker, S.; Belij-Rammerstorfer, S.; Bellamy, D.; Bibi, S.; Bittaye, M.; Clutterbuck, E.A.; et al. Safety and immunogenicity of the ChAdOx1 nCoV-19 vaccine against SARS-CoV-2: A preliminary report of a phase 1/2, single-blind, randomised controlled trial. Lancet 2020, 396, 467–478. [Google Scholar] [CrossRef]
- Stephenson, K.E.; Le Gars, M.; Sadoff, J.; De Groot, A.M.; Heerwegh, D.; Truyers, C.; Atyeo, C.; Loos, C.; Chandrashekar, A.; McMahan, K.; et al. Immunogenicity of the Ad26.COV2.S Vaccine for COVID-19. JAMA—J. Am. Med. Assoc. 2021, 325, 1535–1544. [Google Scholar] [CrossRef] [PubMed]
- Simon, D.; Tascilar, K.; Schmidt, K.; Manger, B.; Weckwerth, L.; Sokolova, M.; Bucci, L.; Fagni, F.; Manger, K.; Schuch, F.; et al. Brief Report: Humoral and cellular immune responses to SARS-CoV-2 infection and vaccination in B cell depleted autoimmune patients. Arthritis Rheumatol. 2022, 74, 33–37. [Google Scholar] [CrossRef]
- Krammer, F. A correlate of protection for SARS-CoV-2 vaccines is urgently needed. Nat. Med. 2021, 27, 1145–1146. [Google Scholar] [CrossRef] [PubMed]
- Bauer, G.; Struck, F.; Schreiner, P.; Staschik, E.; Soutschek, E.; Motz, M. The challenge of avidity determination in SARS-CoV-2 serology. J. Med. Virol. 2021, 93, 3092–3104. [Google Scholar] [CrossRef] [PubMed]
- Struck, F.; Schreiner, P.; Staschik, E.; Wochinz-Richter, K.; Schulz, S.; Soutschek, E.; Motz, M.; Bauer, G. Vaccination versus infection with SARS-CoV-2: Establishment of a high avidity IgG response versus incomplete avidity maturation. J. Med. Virol. 2021, 93, 6765–6777. [Google Scholar] [CrossRef] [PubMed]
- Pratesi, F.; Caruso, T.; Testa, D.; Tarpanelli, T.; Gentili, A.; Gioè, D.; Migliorini, P. Bnt162b2 mrna sars-cov-2 vaccine elicits high avidity and neutralizing antibodies in healthcare workers. Vaccines 2021, 9, 672. [Google Scholar] [CrossRef]
- Taylor, S.C.; Hurst, B.; Martiszus, I.; Hausman, M.S.; Sarwat, S.; Schapiro, J.M.; Rowell, S.; Lituev, A. Semi-quantitative, high throughput analysis of SARS-CoV-2 neutralizing antibodies: Measuring the level and duration of immune response antibodies post infection/vaccination. Vaccine 2021, 39, 5688–5698. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Cytokine | Secretor Cell | Immune Function | Effect on the Host | SARS-CoV-2 Association |
|---|---|---|---|---|
| CXCL-10 | Monocytes | Monocytes, macrophages, NK cells, DCs and LT chemotaxis | Inflammation | Severe disease [70] |
| Interferon-gamma (IFN-Υ) | NK cells and LT-CD4+ (Th1) | IL-4 inhibition, Th1 differentiation, increased MHC I and II expression | Inflammation and anti-viral immune response | Lung injury [71] |
| IL-1β | Macrophages | LB proliferation and differentiation, phagocytes stimulation | Inflammation | Decreased oxygen saturation, poor outcome [72,73] |
| IL-2 | Activated LT-CD4+, LB and monocytes | NK and T cell activation and proliferation, B cell activation along with IL-4 | Inflammation and antigen-specific stimulation | ICU-hospitalization [35,72] |
| IL-4 | LT-CD4+ (Th2) | LB differentiation and proliferation, increased expression of MHC-II | Antigen-specific humoral response | Mild disease [70] |
| IL-6 | Lymphocytes and monocytes | Increased acute inflammation-cytokines release, eosinophil chemotaxis | Immune modulation (pro or anti-inflammatory), antigen-specific response, and anti-viral response | Decreased oxygen saturation, poor outcome, increased risk of death [73,74] |
| IL-8 | Macrophages | Neutrophil and granulocytes chemotaxis, phagocytosis stimulation | Inflammation | Severe disease, increased risk of death [74,75] |
| IL-10 | LT-CD8+ | Inhibition of Th1 cytokines, decreased cytolytic response | Inflammation | Severe disease and ICU hospitalization [35,75] |
| IL-17 | LT-CD4+ (Th17) | Neutrophil activation | Inflammation, mucosal activation, tissue repair | Decreased oxygen-saturation and lung injury [71,73], mild disease [70] |
| Tumor-necrosis factor (TNF)-α | Macrophages | Phagocytes chemotaxis and phagocytosis stimulation | Inflammation | Severe disease, ICU hospitalization, and increased risk of death [35,74,75] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Portilho, A.I.; Gimenes Lima, G.; De Gaspari, E. Enzyme-Linked Immunosorbent Assay: An Adaptable Methodology to Study SARS-CoV-2 Humoral and Cellular Immune Responses. J. Clin. Med. 2022, 11, 1503. https://doi.org/10.3390/jcm11061503
Portilho AI, Gimenes Lima G, De Gaspari E. Enzyme-Linked Immunosorbent Assay: An Adaptable Methodology to Study SARS-CoV-2 Humoral and Cellular Immune Responses. Journal of Clinical Medicine. 2022; 11(6):1503. https://doi.org/10.3390/jcm11061503
Chicago/Turabian StylePortilho, Amanda Izeli, Gabrielle Gimenes Lima, and Elizabeth De Gaspari. 2022. "Enzyme-Linked Immunosorbent Assay: An Adaptable Methodology to Study SARS-CoV-2 Humoral and Cellular Immune Responses" Journal of Clinical Medicine 11, no. 6: 1503. https://doi.org/10.3390/jcm11061503
APA StylePortilho, A. I., Gimenes Lima, G., & De Gaspari, E. (2022). Enzyme-Linked Immunosorbent Assay: An Adaptable Methodology to Study SARS-CoV-2 Humoral and Cellular Immune Responses. Journal of Clinical Medicine, 11(6), 1503. https://doi.org/10.3390/jcm11061503