
Is the JAK-STAT Signaling Pathway Involved in the Pathogenesis of Depression?

 , , ,
, , ,  ,
,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Subjects and Data Collection
2.2. Hamilton Depression Rating Scale (HDRS)
2.3. Biochemical Measurements
2.3.1. mRNA Expression Determination
2.3.2. Protein Expression Determination
2.3.3. Statistical Analysis
3. Results
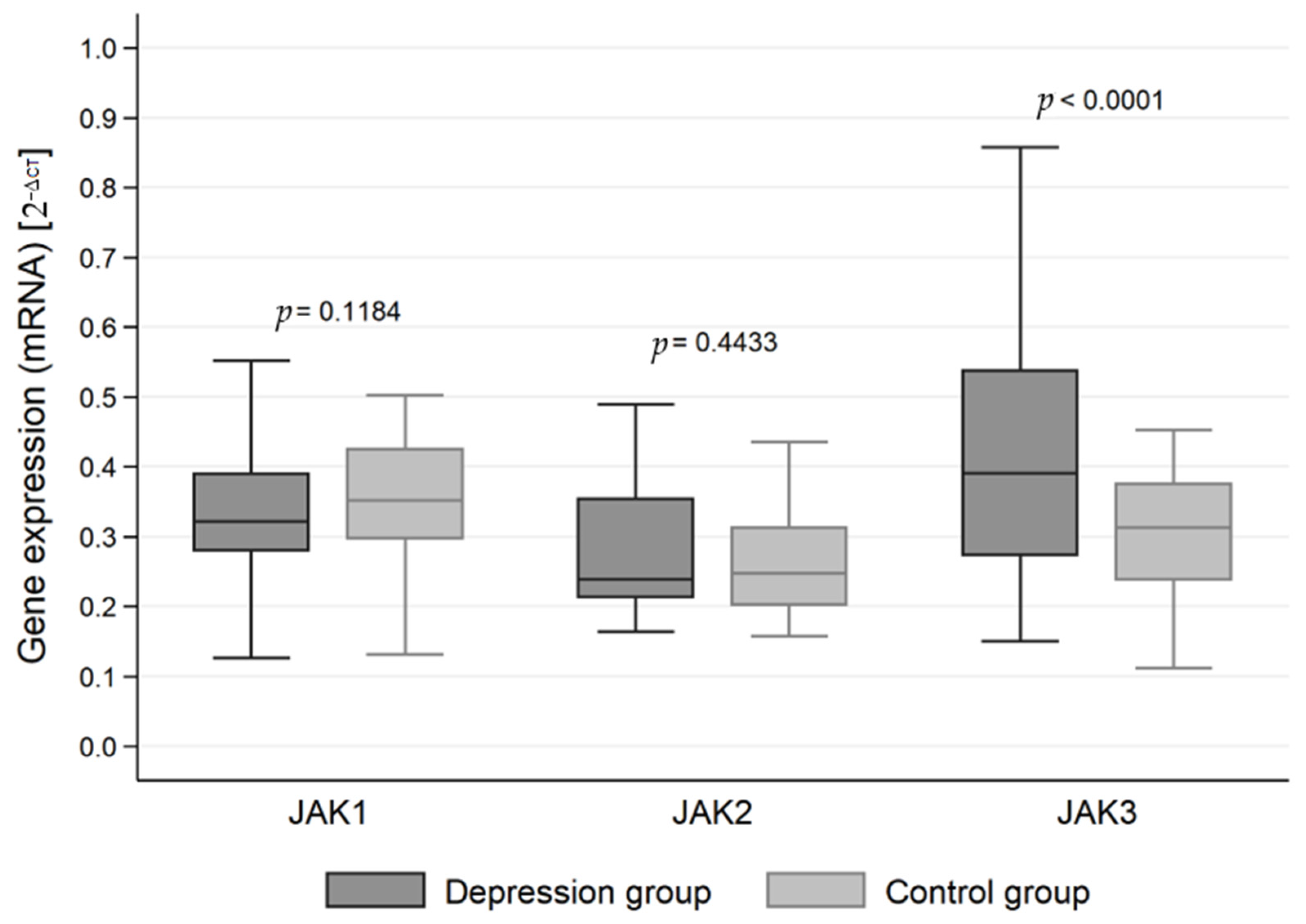
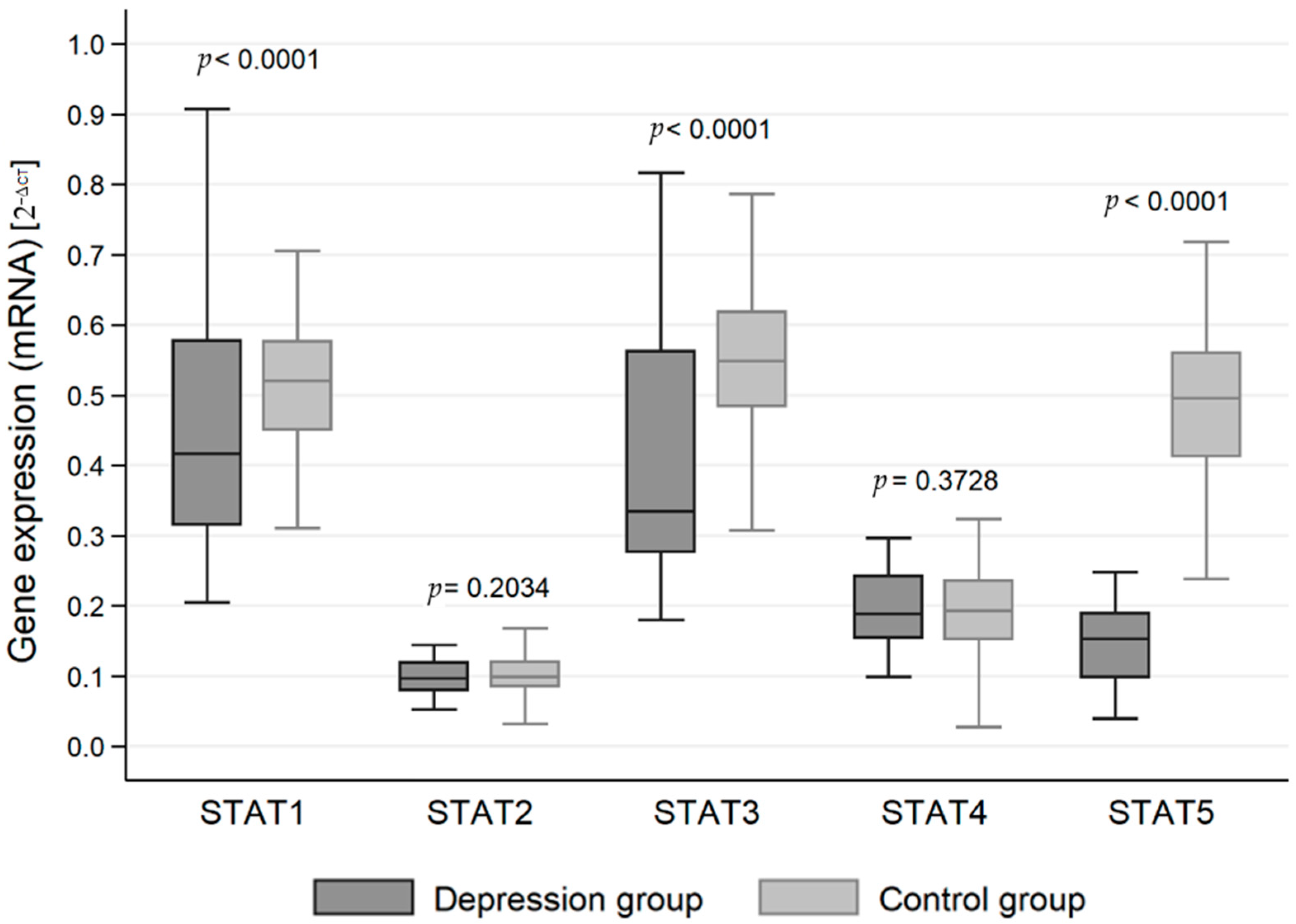
3.1. Mean mRNA Gene Expression
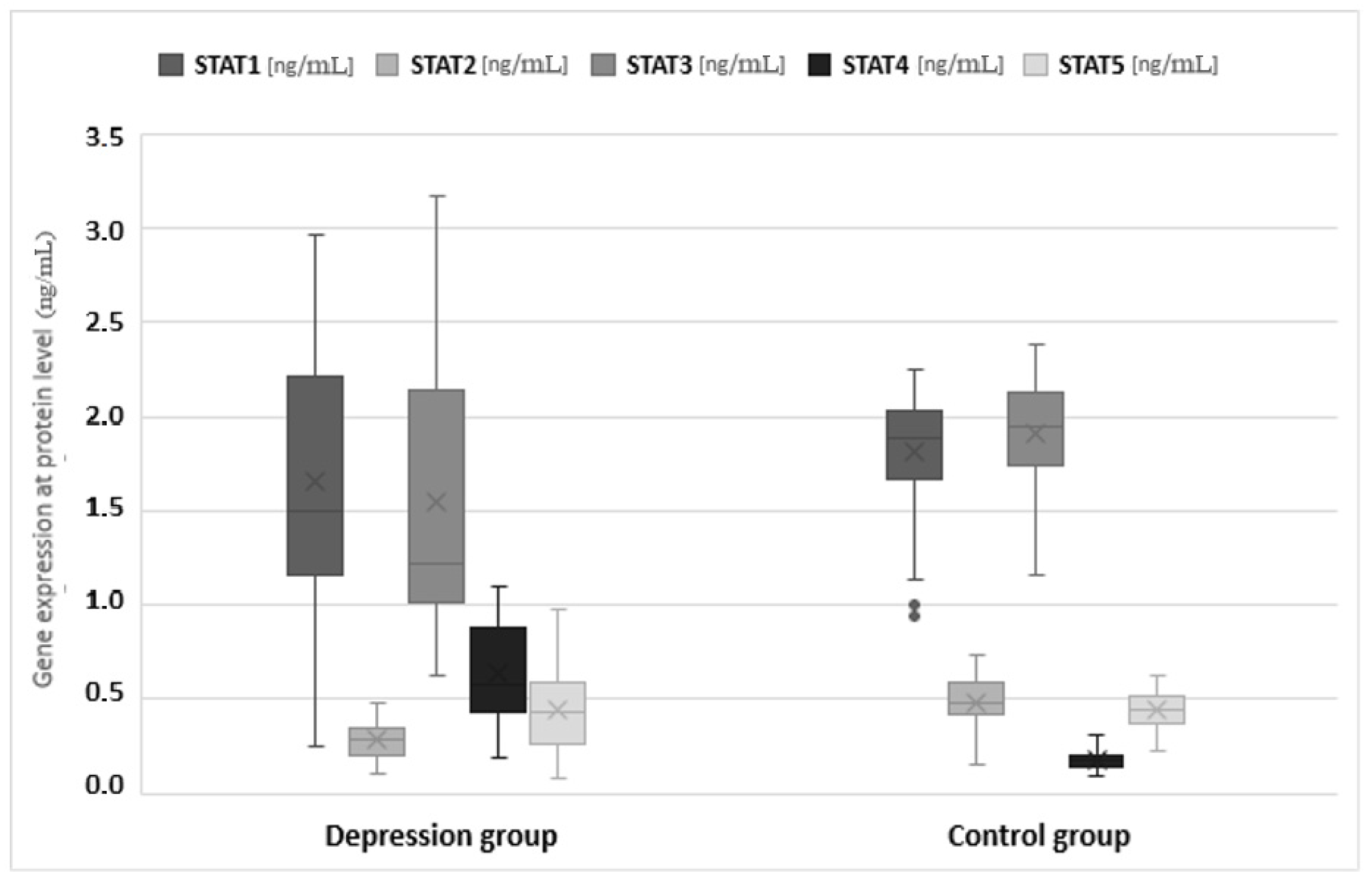
3.2. Mean Protein Expression
3.3. Correlation with Demographic and Clinical Variables
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Malhi, G.S.; Mann, J.J. Depression. Lancet 2018, 392, 2299–2312. [Google Scholar] [CrossRef]
- König, H.; König, H.H.; Konnopka, A. The excess costs of depression: A systematic review and meta-analysis. Epidemiol. Psychiatr. Sci. 2019, 29, e30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rush, A.J.; Trivedi, M.H.; Wisniewski, S.R.; Nierenberg, A.A.; Stewart, J.W.; Warden, D.; Niederehe, G.; Thase, M.E.; Lavori, P.W.; Lebowitz, B.D.; et al. Acute and Longer-Term Outcomes in Depressed Outpatients Requiring One or Several Treatment Steps: A STAR*D Report. Am. J. Psychiatry 2006, 163, 1905–1917. [Google Scholar] [CrossRef] [PubMed]
- Halaris, A.; Sohl, E.; Whitham, E.A. Treatment-Resistant Depression Revisited: A Glimmer of Hope. J. Pers. Med. 2021, 11, 155. [Google Scholar] [CrossRef] [PubMed]
- Gałecka, M.; Bliźniewska-Kowalska, K.; Orzechowska, A.; Szemraj, J.; Maes, M.; Berk, M.; Su, K.-P.; Gałecki, P. Inflammatory versus Anti-Inflammatory Profiles in Major Depressive Disorders—The Role of IL-17, IL-21, IL-23, IL-35 and Foxp3. J. Pers. Med. 2021, 11, 66. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.-H.; Giuliani, F. The Role of Inflammation in Depression and Fatigue. Front. Immunol. 2019, 10, 1696. [Google Scholar] [CrossRef] [Green Version]
- Imada, K.; Leonard, W.J. The Jak-STAT pathway. Mol. Immunol. 2000, 37, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Harrison, D.A. The Jak/STAT pathway. Cold Spring Harb. Perspect. Biol. 2012, 4, a011205. [Google Scholar] [CrossRef] [Green Version]
- Schindler, C.; Plumlee, C. Inteferons pen the JAK-STAT pathway. Semin. Cell Dev. Biol. 2008, 19, 311–318. [Google Scholar] [CrossRef] [Green Version]
- Jatiani, S.S.; Baker, S.J.; Silverman, L.R.; Reddy, E.P. JAK/STAT pathways in cytokine signaling and myeloproliferative disorders: Approaches for targeted therapies. Genes Cancer 2010, 1, 979–993. [Google Scholar] [CrossRef] [Green Version]
- Pesu, M.; Laurence, A.; Kishore, N.; Zwillich, S.H.; Chan, G.; O’Shea, J.J. Therapeutic targeting of Janus kinases. Immunol. Rev. 2008, 223, 132–142. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haan, C.; Behrmann, I.; Haan, S. Perspectives for the use of structural information and chemical genetics to develop inhibitors of Janus kinases. J. Cell. Mol. Med. 2010, 14, 504–527. [Google Scholar] [CrossRef] [PubMed]
- Kukowka, A.; Salmanowicz, M.; Mądra, M.; Banach, B. The role of janus kinases in the treatment of autoimmune skin diseases. Farm. Pol. 2021, 77, 17–24. [Google Scholar] [CrossRef]
- Malemud, C.J. The role of the JAK/STAT signal pathway in rheumatoid arthritis. Ther. Adv. Musculoskelet. Dis. 2018, 10, 117–127, Erratum in Ther. Adv. Musculoskelet. Dis. 2018, 10, 225. [Google Scholar] [CrossRef]
- Pryce, C.R.; Fontana, A. Depression in Autoimmune Diseases. Curr. Top. Behav. Neurosci. 2016, 31, 139–154. [Google Scholar]
- Uysal, S. ICD-10-CM Diagnosis Coding for Neuropsychological Assessment. Arch. Clin. Neuropsychol. 2019, 34, 721–730. [Google Scholar] [CrossRef] [PubMed]
- Hamilton, M. A rating scale for depression. J. Neurol. Neurosurg. Psychiatry 1960, 23, 56–62. [Google Scholar] [CrossRef] [Green Version]
- Schmittgen, T.D.; Livak, K.J. Analyzing real-time PCR data by the comparative CT method. Nat. Protoc. 2008, 3, 1101–1108. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Nicolas, C.S.; Amici, M.; Bortolotto, Z.A.; Doherty, A.; Csaba, Z.; Fafouri, A.; Dournaud, P.; Gressens, P.; Collingridge, G.L.; Peineau, S. The role of JAK-STAT signaling within the CNS. JAK-STAT 2013, 2, e22925. [Google Scholar] [CrossRef] [Green Version]
- Nicolas, C.S.; Peineau, S.; Amici, M.; Csaba, Z.; Fafouri, A.; Javalet, C.; Collett, V.J.; Hildebrandt, L.; Seaton, G.; Choi, S.-L.; et al. The Jak/STAT pathway is involved in synaptic plasticity. Neuron 2012, 73, 374–390. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ng, D.C.; Lin, B.H.; Lim, C.P.; Huang, G.; Zhang, T.; Poli, V.; Cao, X. Stat3 regulates microtubules by antagonizing the depolymerization activity of stathmin. J. Cell Biol. 2006, 172, 245–257. [Google Scholar] [CrossRef] [PubMed]
- Hofmann, H.D.; Kirsch, M. JAK2-STAT3 signaling: A novel function and a novel mechanism. JAK-STAT 2012, 1, 191–193. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maes, M.; Rachayon, M.; Jirakran, K.; Sodsai, P.; Klinchanhom, S.; Gałecki, P.; Sughondhabirom, A. The immune profile of Major Dysmood Disorder: Proof of concept and mechanism using the precision nomothetic psychiatry approach. bioRxiv 2020. [Google Scholar] [CrossRef]
- Planas, A.M.; Gorina, R.; Chamorro, A. Signalling pathways mediating inflammatory responses in brain ischaemia. Biochem. Soc. Trans. 2006, 34, 1267–1270. [Google Scholar] [CrossRef]
- Tsai, M.C.; Chen, W.J.; Tsai, M.S.; Ching, C.H.; Chuang, J.I. Melatonin attenuates brain contusion-induced oxidative insult, inactivation of signal transducers and activators of transcription 1, and upregulation of suppressor of cytokine signaling-3 in rats. J. Pineal Res. 2011, 51, 233–245. [Google Scholar] [CrossRef]
- Lund, I.V.; Hu, Y.; Raol, Y.H.; Benham, R.S.; Faris, R.; Russek, S.J.; Brooks-Kayal, A.R. BDNF selectively regulates GABAA receptor transcription by activation of the JAK/STAT pathway. Sci. Signal. 2008, 1, ra9. [Google Scholar] [CrossRef] [Green Version]
- Chiba, T.; Yamada, M.; Sasabe, J.; Terashita, K.; Shimoda, M.; Matsuoka, M.; Aiso, S. Amyloid-beta causes memory impairment by disturbing the JAK2/STAT3 axis in hippocampal neurons. Mol. Psychiatry 2009, 14, 206–222. [Google Scholar] [CrossRef] [Green Version]
- Mahmoud, G.S.; Grover, L.M. Growth hormone enhances excitatory synaptic transmission in area CA1 of rat hippocampus. J. Neurophysiol. 2006, 95, 2962–2974. [Google Scholar] [CrossRef]
- Orellana, D.I.; Quintanilla, R.A.; Gonzalez-Billault, C.; Maccioni, R.B. Role of the JAKs/STATs pathway in the intracellular calcium changes induced by interleukin-6 in hippocampal neurons. Neurotox. Res. 2005, 8, 295–304. [Google Scholar] [CrossRef]
- Xu, L.; Rensing, N.; Yang, X.F.; Zhang, H.X.; Thio, L.L.; Rothman, S.M.; Weisenfeld, A.E.; Wong, M.; Yamada, K.A. Leptin inhibits 4-aminopyridine- and pentylenetetrazole-induced seizures and AMPAR-mediated synaptic transmission in rodents. J. Clin. Investig. 2008, 118, 272–280. [Google Scholar] [CrossRef] [PubMed]
- Mössner, R.; Mikova, O.; Koutsilieri, E.; Saoud, M.; Ehlis, A.C.; Müller, N.; Fallgatter, A.J.; Riederer, P. Consensus paper of the WFSBP task force onbiological markers: Biological markers in depression. World J. Biol. Psychiatry 2007, 8, 141–174. [Google Scholar] [CrossRef] [PubMed]
- Maes, M.; Mihaylova, I.; Kubera, M.; Uytterhoeven, M.; Vrydags, N.; Bosmans, E. Coenzyme Q10 deficiency in myalgic encephalomyelitis/ chronic fatigue syndrome (ME/CFS) is related to fatigue, autonomic and neurocognitive symptoms and is another risk factor explaining the early mortality in ME/CFS due to cardiovascular disorder. Neuro Endocrinol. Lett. 2009, 30, 470–476. [Google Scholar] [PubMed]
- Behr, G.A.; Moreira, J.C.; Frey, B.N. Preclinical and clinical evidence of antioxidant effects of antidepressant agents: Implications for the pathophysiology of major depressive disorder. Oxid. Med. Cell. Longev. 2012, 2012, 609421. [Google Scholar] [CrossRef]
- Maes, M.; Galecki, P.; Chang, Y.S.; Berk, M. A review on the oxidative and nitrosative stress (O&NS) pathways in major depression and their possible contribution to the (neuro) degenerative processes in that illness. Prog. Neuropsychopharmacol. Biol. Psychiatry 2011, 35, 676–692. [Google Scholar]
- Nunes, S.O.V.; Reiche, E.M.V.; Morimoto, H.K.; Matsuo, T.; Itano, E.N.; Xavier, E.C.; Yamashita, C.M.; Vieira, V.R.; Menoli, A.V.; Silva, S.S.; et al. Immune and hormonal activity in adults suffering from depression. Braz. J. Med. Biol. Res. 2002, 35, 581–587. [Google Scholar] [CrossRef] [Green Version]
- Dienz, O.; Rincon, M. The effects of IL-6 on CD4 T cell responses. Clin. Immunol. 2009, 130, 27–33. [Google Scholar] [CrossRef] [Green Version]
- Talarowska, M.; Szemraj, J.; Gałecki, P. The role of interleukin genes in the course of depression. Open Med. 2016, 11, 41–48. [Google Scholar] [CrossRef]
- O’Shea, J.J.; Schwartz, D.M.; Villarino, A.V.; Gadina, M.; McInnes, I.B.; Laurence, A. The JAK-STAT pathway: Impact on human disease and therapeutic intervention. Annu. Rev. Med. 2015, 66, 311–328. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Analyzed Trait | Study Group | Statistical Parameter * | ||||||
|---|---|---|---|---|---|---|---|---|
| M | Me | Q1–Q3 (IQR) | SD | SE | 95% CI | Min.–Max. | ||
| Age (years) † | Test group | 47.51 | 51 | 41–55 (14) | 11.18 | 0.81 | 45.90–49.10 | 18–67 |
| Control group | 29.36 | 26 | 24–30 (6) | 8.71 | 0.87 | 27.63–31.10 | 20–53 | |
| Overall | 41.29 | 44 | 26–54 (28) | 13.50 | 0.79 | 39.73–42.85 | 18–67 | |
| Number of hospitalizations | Test group only | 2.01 | 1 | 1–2 (1) | 2.00 | 0.15 | 1.72–2.30 | 0–12 |
| Disease duration time (years) | Test group only | 6.18 | 4 | 1–8 (7) | 7.05 | 0.52 | 5.16–7.20 | 1–40 |
| Number of episodes | Test group only | 4.56 | 2 | 1–5 (4) | 5.33 | 0.39 | 3.79–5.34 | 1–20 |
| Hamilton Depression Rating Scale (HDRS) | Test group only | 22.82 | 23 | 18–27 (9) | 6.86 | 0.51 | 21.82–23.82 | 1–51 |
| Gene | Study Group | Statistical Parameter * | |||||||
|---|---|---|---|---|---|---|---|---|---|
| M | Trim. M | Me | Q1–Q3 (IQR) | SD | SE | 95% CI | Min.–Max. | ||
| JAK1 | Test group | 0.349 | 0.348 | 0.321 | 0.279–0.389 (0.110) | 0.107 | 0.008 | 0.334–0.365 | 0.058–0.738 |
| Control group | 0.354 | 0.354 | 0.351 | 0.296–0.425 (0.129) | 0.090 | 0.009 | 0.336–0.372 | 0.131–0.628 | |
| Overall | 0.351 | 0.350 | 0.329 | 0.282–0.422 (0.140) | 0.102 | 0.006 | 0.339–0.363 | 0.058–0.738 | |
| JAK2 | Test group | 0.281 | 0.278 | 0.238 | 0.21–0.354 (0.142) | 0.093 | 0.007 | 0.268–0.295 | 0.163–0.707 |
| Control group | 0.262 | 0.260 | 0.247 | 0.201–0.313 (0.112) | 0.070 | 0.007 | 0.248–0.276 | 0.157–0.435 | |
| Overall | 0.274 | 0.271 | 0.238 | 0.211–0.336 (0.125) | 0.086 | 0.005 | 0.264–0.284 | 0.157–0.707 | |
| JAK3 | Test group | 0.413 | 0.409 | 0.391 | 0.273–0.538 (0.265) | 0.164 | 0.012 | 0.389–0.436 | 0.149–0.858 |
| Control group | 0.306 | 0.307 | 0.312 | 0.236–0.376 (0.140) | 0.085 | 0.008 | 0.289–0.323 | 0.112–0.452 | |
| Overall | 0.376 | 0.371 | 0.333 | 0.261–0.488 (0.227) | 0.150 | 0.009 | 0.358–0.393 | 0.112–0.858 | |
| STAT1 | Test group | 0.444 | 0.441 | 0.416 | 0.315–0.578 (0.263) | 0.160 | 0.012 | 0.421–0.467 | 0.205–0.907 |
| Control group | 0.509 | 0.511 | 0.520 | 0.450–0.577 (0.127) | 0.100 | 0.010 | 0.489–0.528 | 0.233–0.705 | |
| Overall | 0.466 | 0.464 | 0.476 | 0.331–0.578 (0.247) | 0.145 | 0.008 | 0.449–0.483 | 0.205–0.907 | |
| STAT2 | Test group | 0.101 | 0.098 | 0.096 | 0.079–0.119 (0.040) | 0.049 | 0.003 | 0.094–0.108 | 0.052–0.685 |
| Control group | 0.116 | 0.103 | 0.099 | 0.084–0.120 (0.036) | 0.110 | 0.011 | 0.094–0.137 | 0.010–0.915 | |
| Overall | 0.106 | 0.099 | 0.096 | 0.081–0.120 (0.039) | 0.076 | 0.004 | 0.097–0.115 | 0.010–0.915 | |
| STAT3 | Test group | 0.413 | 0.410 | 0.334 | 0.276–0.562 (0.286) | 0.175 | 0.013 | 0.388–0.438 | 0.180–0.817 |
| Control group | 0.548 | 0.549 | 0.548 | 0.483–0.619 (0.136) | 0.100 | 0.010 | 0.529–0.569 | 0.307–0.786 | |
| Overall | 0.460 | 0.459 | 0.473 | 0.293–0.587 (0.294) | 0.166 | 0.010 | 0.441–0.479 | 0.180–0.817 | |
| STAT4 | Test group | 0.196 | 0.196 | 0.188 | 0.154–0.242 (0.088) | 0.052 | 0.004 | 0.188–0.203 | 0.099–0.296 |
| Control group | 0.195 | 0.196 | 0.192 | 0.152–0.236 (0.084) | 0.063 | 0.006 | 0.182–0.207 | 0.019–0.324 | |
| Overall | 0.196 | 0.196 | 0.189 | 0.153–0.242 (0.089) | 0.056 | 0.003 | 0.189–0.202 | 0.019–0.324 | |
| STAT5 | Test group | 0.154 | 0.147 | 0.153 | 0.098–0.189 (0.091) | 0.096 | 0.007 | 0.140–0.168 | 0.039–0.948 |
| Control group | 0.489 | 0.490 | 0.495 | 0.412–0.561 (0.149) | 0.110 | 0.011 | 0.467–0.511 | 0.238–0.718 | |
| Overall | 0.269 | 0.263 | 0.191 | 0.135–0.437 (0.302) | 0.189 | 0.011 | 0.248–0.291 | 0.039–0.948 | |
| Gene | Study Group | Statistical Parameter | ||||||
|---|---|---|---|---|---|---|---|---|
| M | Me | Q1–Q3 (IQR) | SD | SE | 95% CI | Min.–Max. | ||
| JAK1 | Test group | 1.286 | 1.175 | 1.006–1.476 (0.470) | 0.447 | 0.032 | 1.222–1.350 | 0.127–2.861 |
| Control group | 1.321 | 1.307 | 1.093–1.582 (0.489) | 0.288 | 0.029 | 1.264–1.378 | 0.783–1.883 | |
| Overall | 1.298 | 1.199 | 1.071–1.537 (0.466) | 0.399 | 0.023 | 1.252–1.344 | 0.127–2.861 | |
| JAK2 | Test group | 1.017 | 0.853 | 0.742–1.313 (0.571) | 0.353 | 0.026 | 0.966–1.068 | 0.558–1.867 |
| Control group | 1.015 | 0.971 | 0.870–1.129 (0.259) | 0.219 | 0.022 | 0.972–1.058 | 0.498–1.729 | |
| Overall | 1.016 | 0.898 | 0.773–1.221 (0.448) | 0.313 | 0.018 | 0.981–1.051 | 0.498–1.867 | |
| JAK3 | Test group | 1.547 | 1.448 | 0.997–2.079 (1.082) | 0.646 | 0.047 | 1.455–1.640 | 0.499–3.329 |
| Control group | 1.156 | 1.140 | 0.941–1.358 (0.417) | 0.273 | 0.027 | 1.102–1.210 | 0.635–1.668 | |
| Overall | 1.412 | 1.227 | 0.979–1.807 (0.828) | 0.577 | 0.034 | 1.346–1.479 | 0.499–3.329 | |
| STAT1 | Test group | 1.652 | 1.501 | 1.166–2.209 (1.043) | 0.631 | 0.046 | 1.561–1.742 | 0.251–2.958 |
| Control group | 1.811 | 1.883 | 1.667–2.023 (0.356) | 0.299 | 0.030 | 1.752–1.870 | 0.938–2.249 | |
| Overall | 1.706 | 1.762 | 1.212–2.132 (0.920) | 0.545 | 0.032 | 1.644–1.769 | 0.251–2.958 | |
| STAT2 | Test group | 0.283 | 0.289 | 0.205–0.344 (0.139) | 0.086 | 0.006 | 0.271–0295 | 0.109–0.479 |
| Control group | 0.486 | 0.480 | 0.415–0.593 (0.178) | 0.120 | 0.012 | 0.642–0.509 | 0.158–0.735 | |
| Overall | 0.353 | 0.336 | 0.235–0.441 (0.206) | 0.138 | 0.008 | 0.337–0.369 | 0.109–0.735 | |
| STAT3 | Test group | 1.543 | 1.220 | 1.010–2.140 (1.130) | 0.696 | 0.050 | 1.444–1.643 | 0.630–3.170 |
| Control group | 1.910 | 1.943 | 1.738–2.121 (0.383) | 0.274 | 0.027 | 1.857–1.963 | 1.158–2.383 | |
| Overall | 1.670 | 1.726 | 1.085–2.129 (1.044) | 0.611 | 0.036 | 1.599–1.741 | 0.630–3.170 | |
| STAT4 | Test group | 0.640 | 0.574 | 0.437–0.884 (0.447) | 0.250 | 0.018 | 0.605–0.676 | 0.192–1.095 |
| Control group | 0.175 | 0.172 | 0.140–0.208 (0.068) | 0.048 | 0.005 | 0.165–0.184 | 0.098–0.307 | |
| Overall | 0.480 | 0.432 | 0.198–0.752 (0.554) | 0.301 | 0.018 | 0.445–0.515 | 0.098–1.095 | |
| STAT5 | Test group | 0.450 | 0.436 | 0.262–0.592 (0.330) | 0.219 | 0.016 | 0.419–0.481 | 0.083–0.978 |
| Control group | 0.443 | 0.439 | 0.374–0.522 (0.148) | 0.089 | 0.009 | 0.425–0.461 | 0.231–0.628 | |
| Overall | 0.448 | 0.437 | 0.326–0.544 (0.218) | 0.185 | 0.011 | 0.426–0.470 | 0.083–0.978 | |
| Test Group | mRNA Gene Expression | |||||||
|---|---|---|---|---|---|---|---|---|
| Variables | JAK1 | JAK2 | JAK3 | STAT1 | STAT2 | STAT3 | STAT4 | STAT5 |
| Number of hospitalizations | 0.040 (−0.105)–0.183 0.5897 | 0.039 (−0.106)–0.181 0.6012 | −0.015 (−0.159)–0.129 0.8388 | −0.106 (−0.247)–0.038 0.1479 | −0.100 (−0.240)–0.045 0.1745 | 0.010 (−0.134)–0.154 0.8876 | 0.043 (−0.101)–0.186 0.5587 | 0.069 (−0.079)–0.211 0.3469 |
| Disease duration time (years) | 0.054 (−0.091)–0.196 0.4682 | 0.049 (−0.096)–0.191 0.5093 | −0.029 (−0.172)–0.115 0.6938 | −0.009 (−0.153)–0.135 0.9003 | 0.064 (−0.081)–0.206 0.3866 | 0.021 (−0.123)–0.164 0.7749 | 0.074 (−0.071)–0.216 0.3160 | 0.051 (−0.094)–0.193 0.4924 |
| Number of episodes | 0.015 (−0.129)–0.159 0.8339 | 0.118 (−0.026)–0.258 0.1075 | 0.005 (−0.139)–0.148 0.9512 | −0.104 (−0.245)–0.040 0.1558 | −0.095 (−0.236)–0.049 0.1954 | 0.125 (−0.019)–0.264 0.0891 | 0.082 (−0.063)–0.223 0.2675 | 0.047 (−0.098)–0.189 0.5271 |
| Hamilton Depression Rating Scale (HDRS) | −0.102 (−0.243)–0.043 0.1678 | 0.057 (−0.089)–0.200 0.4439 | −0.042 (−0.186)–0.103 0.5678 | 0.067 (−0.079)–0.210 0.3671 | −0.022 (−0.166)–0.123 0.7686 | 0.076 (−0.069)–0.219 0.3036 | −0.111 (−0.252)–0.034 0.1322 | −0.026 (−0.170)–0.120 0.7303 |
| Correlation with Age (Years) | mRNA Gene Expression | |||||||
|---|---|---|---|---|---|---|---|---|
| JAK1 | JAK2 | JAK3 | STAT1 | STAT2 | STAT3 | STAT4 | STAT5 | |
| Test group | −0.056 (−0.197)–0.087 = 0.4438 | −0.019 (−0.161)–0.123 = 0.7909 | −0.047 (−0.188)–0.096 = 0.5194 | −0.041 (−0.182)–0.102 = 0.5752 | −0.047 (−0.188)–0.096 = 0.5229 | 0.001 (−0.142)–0.143 = 0.9932 | −0.012 (−0.154)–0.131 = 0.8737 | −0.055 (−0.196)–0.088 = 0.4503 |
| Control group | −0.211 (−0.392)–(−0.014) = 0.0360 | −0.114 (−0.305)–0.085 = 0.2605 | 0.007 (−0.190)–0.204 = 0.9434 | −0.094 (−0.286)–0.105 = 0.3529 | −0.053 (−0.248)–0.146 = 0.6008 | 0.165 (−0.034)–0.351 = 0.1035 | 0.076 (−0.123)–0.269 = 0.4551 | 0.075 (−0.124)–0.268 = 0.4603 |
| Overall | −0.096 (−0.164)–0.066 = 0.4037 | −0.049 (−0.096)–0.019 = 0.1018 | −0.230 (−0.336)–(−0.118) = 0.0001 | 0.124 0.008–0.236 = 0.0356 | −0.006 (−0.122)–0.109 = 0.9148 | 0.296 0.187–0.398 < 0.0001 | −0.023 (−0.138)–0.093 = 0.6982 | 0.512 0.422–0.593 < 0.0001 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gałecka, M.; Szemraj, J.; Su, K.-P.; Halaris, A.; Maes, M.; Skiba, A.; Gałecki, P.; Bliźniewska-Kowalska, K. Is the JAK-STAT Signaling Pathway Involved in the Pathogenesis of Depression? J. Clin. Med. 2022, 11, 2056. https://doi.org/10.3390/jcm11072056
Gałecka M, Szemraj J, Su K-P, Halaris A, Maes M, Skiba A, Gałecki P, Bliźniewska-Kowalska K. Is the JAK-STAT Signaling Pathway Involved in the Pathogenesis of Depression? Journal of Clinical Medicine. 2022; 11(7):2056. https://doi.org/10.3390/jcm11072056
Chicago/Turabian StyleGałecka, Małgorzata, Janusz Szemraj, Kuan-Pin Su, Angelos Halaris, Michael Maes, Aleksandra Skiba, Piotr Gałecki, and Katarzyna Bliźniewska-Kowalska. 2022. "Is the JAK-STAT Signaling Pathway Involved in the Pathogenesis of Depression?" Journal of Clinical Medicine 11, no. 7: 2056. https://doi.org/10.3390/jcm11072056
APA StyleGałecka, M., Szemraj, J., Su, K. -P., Halaris, A., Maes, M., Skiba, A., Gałecki, P., & Bliźniewska-Kowalska, K. (2022). Is the JAK-STAT Signaling Pathway Involved in the Pathogenesis of Depression? Journal of Clinical Medicine, 11(7), 2056. https://doi.org/10.3390/jcm11072056