


Genome-Wide Pleiotropy Study Identifies Association of PDGFB with Age-Related Macular Degeneration and COVID-19 Infection Outcomes

 , ,
, ,  , and
, and 
Abstract
:1. Introduction
2. Methods
2.1. Genome-Wide Association Summary Statistics for AMD and COVID-19
2.2. Genetic Correlation
2.3. Pleiotropy Genome-Wide Association Study of AMD and COVID-19
2.4. Gene-Based Association Analysis
2.5. Expression/DNA Methylation Quantitative Trait Loci and Colocalization Analyses
2.6. Differential Gene Expression Analysis
2.7. Pathway Analysis Using Gene Ontology
2.8. Mendelian Randomization Analysis
3. Results
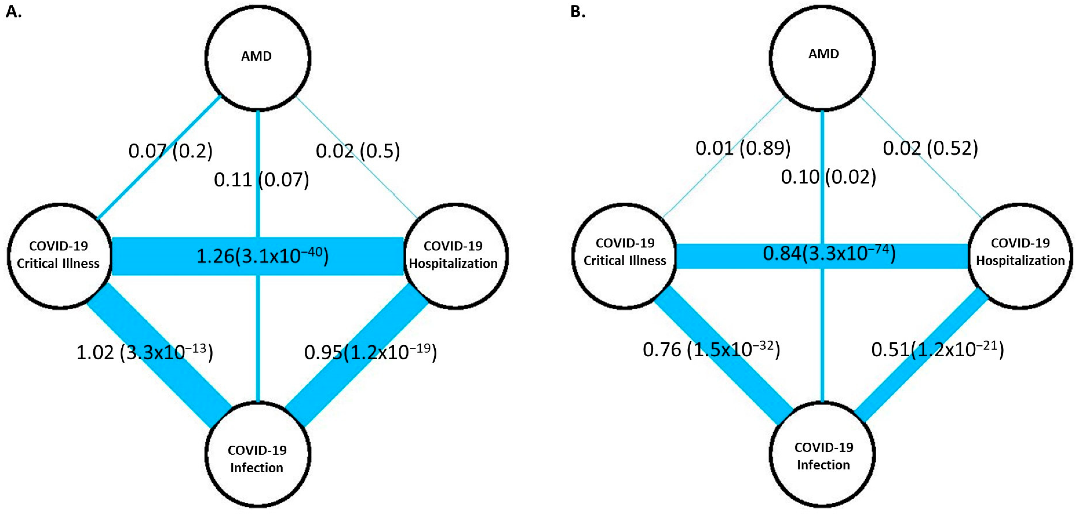
3.1. Genetic Correlation of AMD with COVID-19 Outcomes
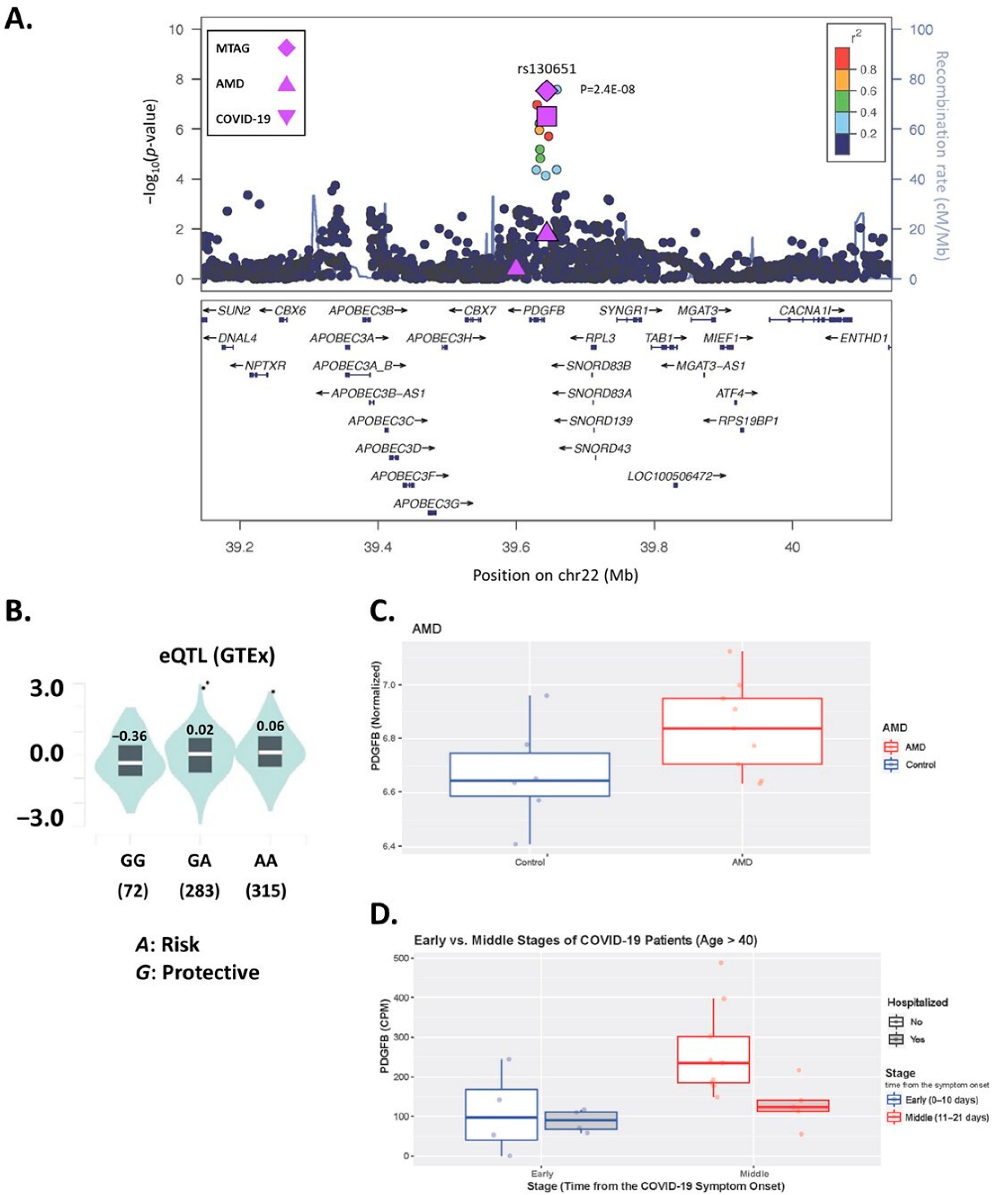
3.2. Multi-Trait Analysis of GWAS for AMD and COVID-19
3.3. PDGFB SNP rs130651 Regulates PDGFB Expression
3.4. PDGFB Is Differentially Expressed in AMD and COVID-19
3.5. Pathway Analysis
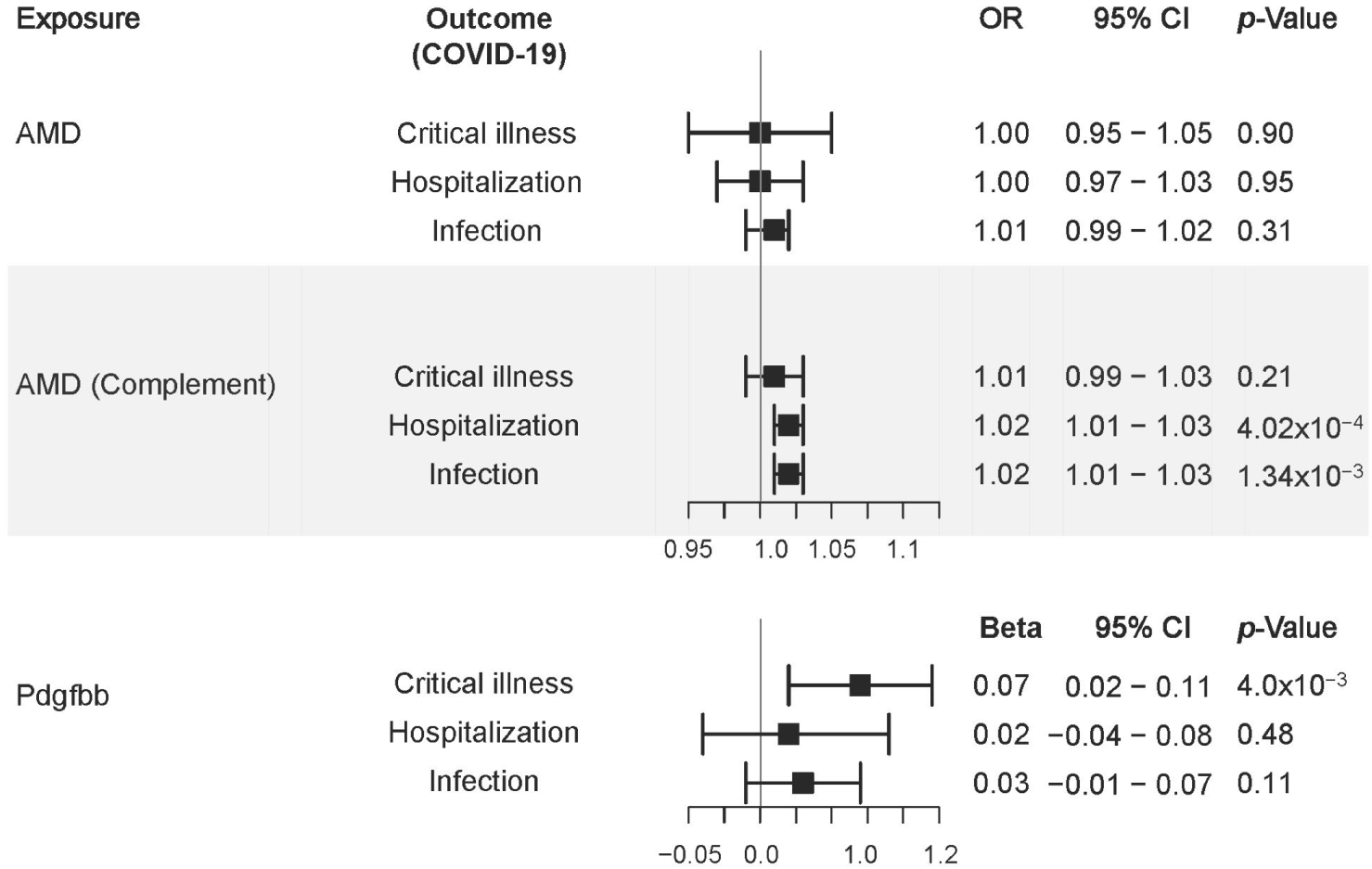
3.6. Mendelian Randomization
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Carter, P.; Anderson, M.; Mossialos, E. Health system, public health, and economic implications of managing COVID-19 from a cardiovascular perspective. Eur. Heart J. 2020, 41, 2516–2518. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Y.; Yang, Q.; Chi, J.; Dong, B.; Lv, W.; Shen, L.; Wang, Y. Comorbidities and the risk of severe or fatal outcomes associated with coronavirus disease 2019: A systematic review and meta-analysis. Int. J. Infect. Dis. 2020, 99, 47–56. [Google Scholar] [CrossRef] [PubMed]
- The Lancet. Neurology, The neurological impact of COVID-19. Lancet Neurol 2020, 19, 471. [Google Scholar] [CrossRef] [PubMed]
- Leung, E.H.; Fan, J.; Flynn, H.W., Jr.; Albini, T.A. Ocular and Systemic Complications of COVID-19: Impact on Patients and Healthcare. Clin. Ophthalmol. 2022, 16, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Ramlall, V.; Thangaraj, P.M.; Meydan, C.; Foox, J.; Butler, D.; Kim, J.; May, B.; De Freitas, J.K.; Glicksberg, B.S.; Mason, C.E.; et al. Immune complement and coagulation dysfunction in adverse outcomes of SARS-CoV-2 infection. Nat. Med. 2020, 26, 1609–1615. [Google Scholar] [CrossRef]
- Yang, J.M.; Moon, S.Y.; Lee, J.Y.; Agalliu, D.; Yon, D.K.; Lee, S.W. COVID-19 Morbidity and Severity in Patients with Age-Related Macular Degeneration: A Korean Nationwide Cohort Study. Am. J. Ophthalmol. 2021, 239, 159–169. [Google Scholar] [CrossRef]
- Smith, W.; Assink, J.; Klein, R.; Mitchell, P.; Klaver, C.C.; Klein, B.E.; Hofman, A.; Jensen, S.; Wang, J.J.; de Jong, P.T. Risk factors for age-related macular degeneration: Pooled findings from three continents. Ophthalmology 2001, 108, 697–704. [Google Scholar] [CrossRef]
- Chakravarthy, U.; Wong, T.Y.; Fletcher, A.; Piault, E.; Evans, C.; Zlateva, G.; Buggage, R.; Pleil, A.; Mitchell, P. Clinical risk factors for age-related macular degeneration: A systematic review and meta-analysis. BMC Ophthalmol. 2010, 10, 31. [Google Scholar] [CrossRef] [Green Version]
- Fritsche, L.G.; Igl, W.; Bailey, J.N.; Grassmann, F.; Sengupta, S.; Bragg-Gresham, J.L.; Burdon, K.P.; Hebbring, S.J.; Wen, C.; Gorski, M.; et al. A large genome-wide association study of age-related macular degeneration highlights contributions of rare and common variants. Nat. Genet. 2016, 48, 134–143. [Google Scholar] [CrossRef] [Green Version]
- COVID-19 Host Genetics Initiative. Mapping the human genetic architecture of COVID-19. Nature 2021, 600, 472–477. [Google Scholar] [CrossRef]
- Bulik-Sullivan, B.; Finucane, H.K.; Anttila, V.; Gusev, A.; Day, F.R.; Loh, P.R.; ReproGen Consortium; Psychiatric Genomics Consortium; Genetic Consortium for Anorexia Nervosa of the Wellcome Trust Case Control Consortium 3; Duncan, L.; et al. An atlas of genetic correlations across human diseases and traits. Nat. Genet. 2015, 47, 1236–1241. [Google Scholar] [CrossRef] [Green Version]
- Lu, Q.; Li, B.; Ou, D.; Erlendsdottir, M.; Powles, R.L.; Jiang, T.; Hu, Y.; Chang, D.; Jin, C.; Dai, W.; et al. A Powerful Approach to Estimating Annotation-Stratified Genetic Covariance via GWAS Summary Statistics. Am. J. Hum. Genet. 2017, 101, 939–964. [Google Scholar] [CrossRef] [Green Version]
- Turley, P.; Walters, R.K.; Maghzian, O.; Okbay, A.; Lee, J.J.; Fontana, M.A.; Nguyen-Viet, T.A.; Wedow, R.; Zacher, M.; Furlotte, N.A.; et al. Multi-trait analysis of genome-wide association summary statistics using MTAG. Nat. Genet. 2018, 50, 229–237. [Google Scholar] [CrossRef]
- Chung, J.; Zhang, X.; Allen, M.; Wang, X.; Ma, Y.; Beecham, G.; Montine, T.J.; Younkin, S.G.; Dickson, D.W.; Golde, T.E.; et al. Genome-wide pleiotropy analysis of neuropathological traits related to Alzheimer’s disease. Alzheimer’s Res. Ther. 2018, 10, 22. [Google Scholar] [CrossRef]
- Barbeira, A.N.; Dickinson, S.P.; Bonazzola, R.; Zheng, J.; Wheeler, H.E.; Torres, J.M.; Torstenson, E.S.; Shah, K.P.; Garcia, T.; Edwards, T.L.; et al. Exploring the phenotypic consequences of tissue specific gene expression variation inferred from GWAS summary statistics. Nat. Commun. 2018, 9, 1825. [Google Scholar] [CrossRef]
- Consortium, G.T. The Genotype-Tissue Expression (GTEx) project. Nat Genet 2013, 45, 580–585. [Google Scholar] [CrossRef]
- Kwong, A.; Boughton, A.P.; Wang, M.; VandeHaar, P.; Boehnke, M.; Abecasis, G.; Kang, H.M. FIVEx: An interactive eQTL browser across public datasets. Bioinformatics 2021, 38, 559–561. [Google Scholar] [CrossRef]
- Giambartolomei, C.; Vukcevic, D.; Schadt, E.E.; Franke, L.; Hingorani, A.D.; Wallace, C.; Plagnol, V. Bayesian test for colocalisation between pairs of genetic association studies using summary statistics. PLoS Genet. 2014, 10, e1004383. [Google Scholar] [CrossRef] [Green Version]
- Bonilla, C.; Bertoni, B.; Min, J.L.; Hemani, G.; Genetics of DNA Methylation Consortium; Elliott, H.R. Investigating DNA methylation as a potential mediator between pigmentation genes, pigmentary traits and skin cancer. Pigment Cell Melanoma Res. 2021, 34, 892–904. [Google Scholar] [CrossRef]
- Whitmore, S.S.; Braun, T.A.; Skeie, J.M.; Haas, C.M.; Sohn, E.H.; Stone, E.M.; Scheetz, T.E.; Mullins, R.F. Altered gene expression in dry age-related macular degeneration suggests early loss of choroidal endothelial cells. Mol. Vis. 2013, 19, 2274–2297. [Google Scholar]
- McClain, M.T.; Constantine, F.J.; Henao, R.; Liu, Y.; Tsalik, E.L.; Burke, T.W.; Steinbrink, J.M.; Petzold, E.; Nicholson, B.P.; Rolfe, R.; et al. Dysregulated transcriptional responses to SARS-CoV-2 in the periphery. Nat. Commun. 2021, 12, 1079. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Layana, A.; Cabrera-Lopez, F.; Garcia-Arumi, J.; Arias-Barquet, L.; Ruiz-Moreno, J.M. Early and intermediate age-related macular degeneration: Update and clinical review. Clin. Interv. Aging 2017, 12, 1579–1587. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Watanabe, K.; Taskesen, E.; van Bochoven, A.; Posthuma, D. Functional mapping and annotation of genetic associations with FUMA. Nat. Commun. 2017, 8, 1826. [Google Scholar] [CrossRef] [Green Version]
- Johnson, L.V.; Leitner, W.P.; Staples, M.K.; Anderson, D.H. Complement activation and inflammatory processes in Drusen formation and age related macular degeneration. Exp. Eye Res. 2001, 73, 887–896. [Google Scholar] [CrossRef] [PubMed]
- Hageman, G.S.; Luthert, P.J.; Victor Chong, N.H.; Johnson, L.V.; Anderson, D.H.; Mullins, R.F. An integrated hypothesis that considers drusen as biomarkers of immune-mediated processes at the RPE-Bruch’s membrane interface in aging and age-related macular degeneration. Prog. Retin. Eye Res. 2001, 20, 705–732. [Google Scholar] [CrossRef]
- Ahola-Olli, A.V.; Wurtz, P.; Havulinna, A.S.; Aalto, K.; Pitkanen, N.; Lehtimaki, T.; Kahonen, M.; Lyytikainen, L.P.; Raitoharju, E.; Seppala, I.; et al. Genome-wide Association Study Identifies 27 Loci Influencing Concentrations of Circulating Cytokines and Growth Factors. Am. J. Hum. Genet. 2017, 100, 40–50. [Google Scholar] [CrossRef] [Green Version]
- Yavorska, O.O.; Burgess, S. MendelianRandomization: An R package for performing Mendelian randomization analyses using summarized data. Int. J. Epidemiol. 2017, 46, 1734–1739. [Google Scholar] [CrossRef] [Green Version]
- Orozco, L.D.; Chen, H.H.; Cox, C.; Katschke, K.J., Jr.; Arceo, R.; Espiritu, C.; Caplazi, P.; Nghiem, S.S.; Chen, Y.J.; Modrusan, Z.; et al. Integration of eQTL and a Single-Cell Atlas in the Human Eye Identifies Causal Genes for Age-Related Macular Degeneration. Cell Rep. 2020, 30, 1246–1259. [Google Scholar] [CrossRef] [Green Version]
- Zehetner, C.; Kirchmair, R.; Neururer, S.B.; Kralinger, M.T.; Bechrakis, N.E.; Kieselbach, G.F. Systemic upregulation of PDGF-B in patients with neovascular AMD. Investig. Ophthalmol. Vis. Sci. 2014, 55, 337–344. [Google Scholar] [CrossRef] [Green Version]
- Huang, C.; Wang, Y.; Li, X.; Ren, L.; Zhao, J.; Hu, Y.; Zhang, L.; Fan, G.; Xu, J.; Gu, X.; et al. Clinical features of patients infected with 2019 novel coronavirus in Wuhan, China. Lancet 2020, 395, 497–506. [Google Scholar] [CrossRef] [Green Version]
- Petrey, A.C.; Qeadan, F.; Middleton, E.A.; Pinchuk, I.V.; Campbell, R.A.; Beswick, E.J. Cytokine release syndrome in COVID-19: Innate immune, vascular, and platelet pathogenic factors differ in severity of disease and sex. J. Leukoc. Biol. 2021, 109, 55–66. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Zhang, C.; Huang, F.; Yang, Y.; Wang, F.; Yuan, J.; Zhang, Z.; Qin, Y.; Li, X.; Zhao, D.; et al. Elevated plasma levels of selective cytokines in COVID-19 patients reflect viral load and lung injury. Natl. Sci. Rev. 2020, 7, 1003–1011. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Diaz-Lezama, N.; Wolf, A.; Koch, S.; Pfaller, A.M.; Biber, J.; Guillonneau, X.; Langmann, T.; Grosche, A. PDGF Receptor Alpha Signaling Is Key for Muller Cell Homeostasis Functions. Int. J. Mol. Sci. 2021, 22, 1174. [Google Scholar] [CrossRef] [PubMed]
- Jaffe, G.J.; Ciulla, T.A.; Ciardella, A.P.; Devin, F.; Dugel, P.U.; Eandi, C.M.; Masonson, H.; Mones, J.; Pearlman, J.A.; Quaranta-El Maftouhi, M.; et al. Dual Antagonism of PDGF and VEGF in Neovascular Age-Related Macular Degeneration: A Phase IIb, Multicenter, Randomized Controlled Trial. Ophthalmology 2017, 124, 224–234. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, D.B.; Zheng, J. Regulation of placental angiogenesis. Microcirculation 2014, 21, 15–25. [Google Scholar] [CrossRef] [Green Version]
- Skendros, P.; Mitsios, A.; Chrysanthopoulou, A.; Mastellos, D.C.; Metallidis, S.; Rafailidis, P.; Ntinopoulou, M.; Sertaridou, E.; Tsironidou, V.; Tsigalou, C.; et al. Complement and tissue factor-enriched neutrophil extracellular traps are key drivers in COVID-19 immunothrombosis. J. Clin. Investig. 2020, 130, 6151–6157. [Google Scholar] [CrossRef]
- Lo, M.W.; Kemper, C.; Woodruff, T.M. COVID-19: Complement, Coagulation, and Collateral Damage. J. Immunol. 2020, 205, 1488–1495. [Google Scholar] [CrossRef]
- Java, A.; Apicelli, A.J.; Liszewski, M.K.; Coler-Reilly, A.; Atkinson, J.P.; Kim, A.H.; Kulkarni, H.S. The complement system in COVID-19: Friend and foe? JCI Insight 2020, 5, e140711. [Google Scholar] [CrossRef]
- Klein, R.; Moss, S.E.; Meuer, S.M.; Klein, B.E. The 15-year cumulative incidence of retinal vein occlusion: The Beaver Dam Eye Study. Arch. Ophthalmol. 2008, 126, 513–518. [Google Scholar] [CrossRef] [Green Version]
- Bair, P.J.; Hsia, N.Y.; Lin, C.L.; Yang, Y.C.; Shen, T.C.; Li, C.Y. Population-based retrospective cohort study on risk of age-related macular degeneration in people with chronic obstructive pulmonary disease. Sci. Rep. 2021, 11, 15079. [Google Scholar] [CrossRef]
- Armento, A.; Ueffing, M.; Clark, S.J. The complement system in age-related macular degeneration. Cell. Mol. Life Sci. 2021, 78, 4487–4505. [Google Scholar] [CrossRef]
- Holter, J.C.; Pischke, S.E.; de Boer, E.; Lind, A.; Jenum, S.; Holten, A.R.; Tonby, K.; Barratt-Due, A.; Sokolova, M.; Schjalm, C.; et al. Systemic complement activation is associated with respiratory failure in COVID-19 hospitalized patients. Proc. Natl. Acad. Sci. USA 2020, 117, 25018–25025. [Google Scholar] [CrossRef]
- Geerlings, M.J.; de Jong, E.K.; den Hollander, A.I. The complement system in age-related macular degeneration: A review of rare genetic variants and implications for personalized treatment. Mol. Immunol. 2017, 84, 65–76. [Google Scholar] [CrossRef]
- Lakshmanan, M.; Schaffer, A.; Robbins, J.; Reynolds, J.; Norton, J. A simplified low iodine diet in I-131 scanning and therapy of thyroid cancer. Clin. Nucl. Med. 1988, 13, 866–868. [Google Scholar] [CrossRef]
- Allegrini, D.; Raimondi, R.; Montesano, G.; Borgia, A.; Sorrentino, T.; Tsoutsanis, P.; Romano, M.R. Short-Term Outcomes After COVID-19-Related Treatment Interruption Among Patients with Neovascular Age-Related Macular Degeneration Receiving Intravitreal Bevacizumab. Clin. Ophthalmol. 2021, 15, 4073–4079. [Google Scholar] [CrossRef]
- Tekola-Ayele, F.; Lee, A.; Workalemahu, T.; Sanchez-Pozos, K. Shared genetic underpinnings of childhood obesity and adult cardiometabolic diseases. Hum. Genom. 2019, 13, 17. [Google Scholar] [CrossRef] [Green Version]
- Vorstman, J.A.; Breetvelt, E.J.; Thode, K.I.; Chow, E.W.; Bassett, A.S. Expression of autism spectrum and schizophrenia in patients with a 22q11.2 deletion. Schizophr. Res. 2013, 143, 55–59. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| rs130651 (EA/NEA = G/A; EAF = 0.32) | rs4820371 (EA/NEA = C/T; EAF = 0.23) | |||||||
|---|---|---|---|---|---|---|---|---|
| Analytical Method | Trait | OR | 95% CI | p | OR | 95% CI | p | |
| Single GWAS | AMD | 0.90 | 0.87–0.94 | 1.36 × 10−7 | 0.90 | 0.86–0.93 | 1.71 × 10−7 | |
| COVID-19 | Critical Illness | 0.96 | 0.89–1.04 | 0.27 | 1.01 | 0.93–1.09 | 0.78 | |
| Hospitalization | 1.00 | 0.96–1.04 | 0.87 | 1.00 | 0.94–1.06 | 0.93 | ||
| Infections | 0.98 | 0.96–1.00 | 0.024 | 0.96 | 0.94–0.98 | 0.003 | ||
| MTAG of AMD with COVID-19 | COVID-19 | Critical Illness | 0.96 | 0.94–0.97 | 3.5 × 10−8 | 0.95 | 0.93–0.97 | 6.1 × 10−8 |
| Hospitalization | 0.96 | 0.94–0.97 | 4.4 × 10−8 | 0.95 | 0.93–0.97 | 5.7 × 10−8 | ||
| Infections | 0.96 | 0.94–0.97 | 2.4 × 10−8 | 0.95 | 0.93–0.97 | 2.6 × 10−8 | ||
| Scheme | Study | Tissue/Cell Type | Effect | SE | p-Value |
|---|---|---|---|---|---|
| rs130651 (A) | GTEx | Whole blood | 0.26 | 0.04 | 1.8 × 10−11 |
| Lepik et al. 2017 | Whole blood | 0.31 | 0.031 | 8.5 × 10−21 | |
| BLUEPRINT (Immune cells) | T cell | 0.87 | 0.083 | 7.1 × 10−20 | |
| DICE (Immune cells) | Th17 cell | 0.96 | 0.081 | 8.7 × 10−19 | |
| Tfh cell | 0.63 | 0.056 | 4.7 × 10−18 | ||
| T cell | 1.44 | 0.14 | 1.2 × 10−15 | ||
| Th2 cell | 1.11 | 0.11 | 7.1 × 10−15 | ||
| CD4 T cell (naïve) | 1.13 | 0.12 | 8.1 × 10−14 | ||
| Th1-17 cell | 0.54 | 0.07 | 5.1 × 10−11 | ||
| T regulatory cell | 0.61 | 0.087 | 9.3 × 10−10 | ||
| Th1 cell | 0.51 | 0.075 | 3.4 × 10−9 | ||
| CD8 T cell (naïve) | 0.85 | 0.13 | 6.2 × 10−9 | ||
| TwinsUK | Whole blood | 0.22 | 0.036 | 3.8 × 10−9 | |
| rs4820371 (T) | GTEx | Whole blood | 0.13 | 0.04 | 3.5 × 10−3 |
| Lepik et al. 2017 | Whole blood | 0.20 | 0.035 | 5.8 × 10−8 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chung, J.; Vig, V.; Sun, X.; Han, X.; O’Connor, G.T.; Chen, X.; DeAngelis, M.M.; Farrer, L.A.; Subramanian, M.L. Genome-Wide Pleiotropy Study Identifies Association of PDGFB with Age-Related Macular Degeneration and COVID-19 Infection Outcomes. J. Clin. Med. 2023, 12, 109. https://doi.org/10.3390/jcm12010109
Chung J, Vig V, Sun X, Han X, O’Connor GT, Chen X, DeAngelis MM, Farrer LA, Subramanian ML. Genome-Wide Pleiotropy Study Identifies Association of PDGFB with Age-Related Macular Degeneration and COVID-19 Infection Outcomes. Journal of Clinical Medicine. 2023; 12(1):109. https://doi.org/10.3390/jcm12010109
Chicago/Turabian StyleChung, Jaeyoon, Viha Vig, Xinyu Sun, Xudong Han, George T. O’Connor, Xuejing Chen, Margaret M. DeAngelis, Lindsay A. Farrer, and Manju L. Subramanian. 2023. "Genome-Wide Pleiotropy Study Identifies Association of PDGFB with Age-Related Macular Degeneration and COVID-19 Infection Outcomes" Journal of Clinical Medicine 12, no. 1: 109. https://doi.org/10.3390/jcm12010109
APA StyleChung, J., Vig, V., Sun, X., Han, X., O’Connor, G. T., Chen, X., DeAngelis, M. M., Farrer, L. A., & Subramanian, M. L. (2023). Genome-Wide Pleiotropy Study Identifies Association of PDGFB with Age-Related Macular Degeneration and COVID-19 Infection Outcomes. Journal of Clinical Medicine, 12(1), 109. https://doi.org/10.3390/jcm12010109