
Inhibition of γδ-TcR or IL17a Reduces T-Cell and Neutrophil Infiltration after Ischemia/Reperfusion Injury in Mouse Liver
 , , ,
, , , 

 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Procedure
- Sham group. Animals were anaesthetized and subjected to a sham surgical operation that involved a midline incision equivalent to the experimental groups but with no ischemia time;
- I/R group. Animals were subjected to partial hepatic ischemia for 60 min followed by 6 h of reperfusion;
- I/R + anti-γδTcR group. Animals were pretreated with an intraperitoneal injection of 500 μg of anti-γδTcR antibody [21] (Purified NA/LE Hamster anti-mouse γδ-Τ-cell Receptor, UC7-13D5, Catalog Number: 553181, BD PharmingenTM, San Jose, CA, USA) 72 h before induction of ischemia;
- I/R + anti-IL17 group. Animals were pretreated with a 100 μg anti-IL17a antibody [22] (Mouse IL-17/IL -17A Antibody Monoclonal Rat IgG2A Clone # 50104, Catalog Number: MAB421) injection into the inferior vena cava (IVC) before induction of ischemia.
2.2. Histopathology, Immunohistochemistry and Morphometry
2.3. Quantitative Gene Expression Analysis
2.4. Determination of Malondialdehyde (MDA) in Liver Tissues
2.5. Statistical Analyses
3. Results
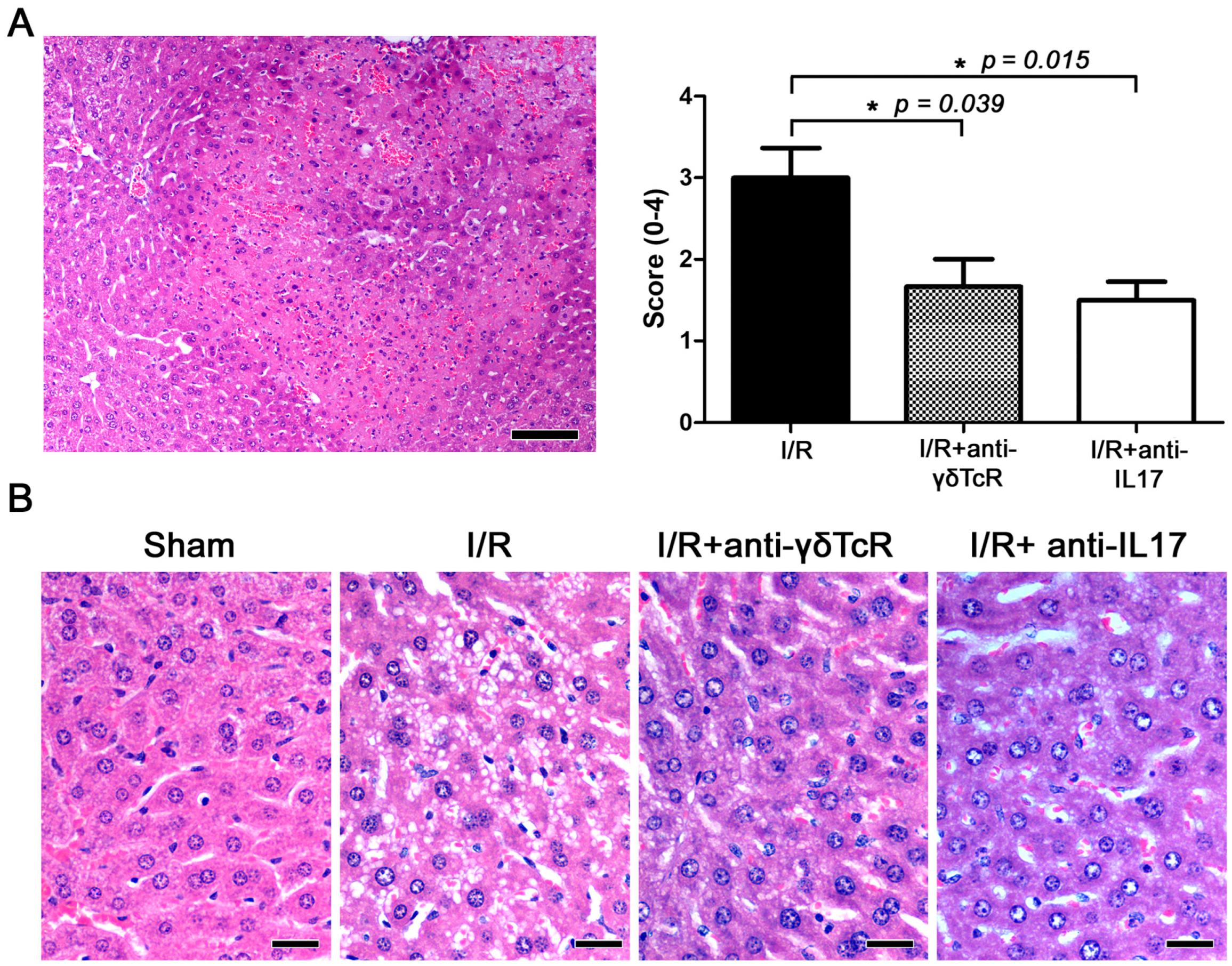
3.1. Depletion of Either γδ-T Cells or IL17 Ameliorates I/R-Induced Liver Injury
3.2. Anti-γδ-TcR and Anti-IL17 Treatment Reduce I/R-Induced Inflammatory Cell Infiltration
3.3. Anti-γδTcR and Anti-IL17 Suppress I/R-Associated Inflammatory Cytokines
3.4. Anti-γδTcR and Anti-IL17 Alter the Expression of I/R-Associated Inflammatory and Cell Stress Factors
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Abu-Amara, M.; Yang, S.Y.; Tapuria, N.; Fuller, B.; Davidson, B.; Seifalian, A. Liver Ischemia/Reperfusion Injury: Processes in Inflammatory Networks—A Review. Liver Transplant. 2010, 16, 1016–1032. [Google Scholar] [CrossRef] [PubMed]
- Datta, G.; Fuller, B.J.; Davidson, B.R. Molecular Mechanisms of Liver Ischemia Reperfusion Injury: Insights from Transgenic Knockout Models. World J. Gastroenterol. 2013, 19, 1683–1698. [Google Scholar] [CrossRef] [PubMed]
- Dar, W.A.; Sullivan, E.; Bynon, J.S.; Eltzschig, H.; Ju, C. Ischaemia Reperfusion Injury in Liver Transplantation: Cellular and Molecular Mechanisms. Liver Int. 2019, 39, 788–801. [Google Scholar] [CrossRef] [Green Version]
- Konishi, T.; Lentsch, A.B. Hepatic Ischemia/Reperfusion: Mechanisms of Tissue Injury, Repair, and Regeneration. Gene Expr. 2017, 17, 277–287. [Google Scholar] [CrossRef]
- Linfert, D.; Chowdhry, T.; Rabb, H. Lymphocytes and Ischemia-Reperfusion Injury. Transplant. Rev. 2009, 23, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zwacka, R.M.; Zhou, W.; Zhang, Y.; Darby, C.J.; Dudus, L.; Halldorson, J.; Oberley, L.; Engelhardt, J.F. Redox Gene Therapy for Ischemia/Reperfusion Injury of the Liver Reduces AP1 and NF-ΚB Activation. Nat. Med. 1998, 4, 698–704. [Google Scholar] [CrossRef] [PubMed]
- Satpute, S.R.; Park, J.M.; Jang, H.R.; Agreda, P.; Liu, M.; Gandolfo, M.T.; Racusen, L.; Rabb, H. The Role for T Cell Repertoire/Antigen-Specific Interactions in Experimental Kidney Ischemia Reperfusion Injury. J. Immunol. 2009, 183, 984–992. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shen, X.; Wang, Y.; Gao, F.; Ren, F.; Busuttil, R.W.; Kupiec-Weglinski, J.W.; Zhai, Y. CD4 T Cells Promote Tissue Inflammation via CD40 Signaling without de Novo Activation in a Murine Model of Liver Ischemia/Reperfusion Injury. Hepatology 2009, 50, 1537–1546. [Google Scholar] [CrossRef] [Green Version]
- Abe, Y.; Hines, I.N.; Zibari, G.; Pavlick, K.; Gray, L.; Kitagawa, Y.; Grisham, M.B. Mouse Model of Liver Ischemia and Reperfusion Injury: Method for Studying Reactive Oxygen and Nitrogen Metabolites In Vivo. Free Radic. Biol. Med. 2009, 46, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Yao, Z.; Painter, S.L.; Fanslow, W.C.; Ulrich, D.; Macduff, B.M.; Spriggs, M.K.; Armitage, R.J. Human IL-17: A Novel Cytokine Derived from T Cells. J. Immunol. 1995, 155, 5483–5486. [Google Scholar] [CrossRef]
- Kono, H.; Fujii, H.; Ogiku, M.; Hosomura, N.; Amemiya, H.; Tsuchiya, M.; Hara, M. Role of IL-17A in Neutrophil Recruitment and Hepatic Injury after Warm Ischemia–Reperfusion Mice. J. Immunol. 2011, 187, 4818–4825. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hayday, A.C. Γδ T Cell Update: Adaptate Orchestrators of Immune Surveillance. J. Immunol. 2019, 203, 311–320. [Google Scholar] [CrossRef] [PubMed]
- Hammerich, L.; Tacke, F. Role of Gamma-Delta T Cells in Liver Inflammation and Fibrosis. World J. Gastrointest. Pathophysiol. 2014, 5, 107–113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuboki, S.; Shin, T.; Huber, N.; Eismann, T.; Galloway, E.; Schuster, R.; Blanchard, J.; Zingarelli, B.; Lentsch, A.B. Peroxisome Proliferator-Activated Receptor-Gamma Protects against Hepatic Ischemia/Reperfusion Injury in Mice. Hepatology 2008, 47, 215–224. [Google Scholar] [CrossRef]
- Marion-Letellier, R.; Savoye, G.; Ghosh, S. Fatty Acids, Eicosanoids and PPAR Gamma. Eur. J. Pharmacol. 2016, 785, 44–49. [Google Scholar] [CrossRef]
- G Bardallo, R.; Panisello-Roselló, A.; Sanchez-Nuno, S.; Alva, N.; Roselló-Catafau, J.; Carbonell, T. Nrf2 and Oxidative Stress in Liver Ischemia/Reperfusion Injury. FEBS J. 2022, 289, 5463–5479. [Google Scholar] [CrossRef]
- Ke, B.; Shen, X.-D.; Zhang, Y.; Ji, H.; Gao, F.; Yue, S.; Kamo, N.; Zhai, Y.; Yamamoto, M.; Busuttil, R.W.; et al. KEAP1-NRF2 complex in ischemia-induced hepatocellular damage of mouse liver transplants. J. Hepatol. 2013, 59, 1200–1207. [Google Scholar] [CrossRef] [Green Version]
- Kudoh, K.; Uchinami, H.; Yoshioka, M.; Seki, E.; Yamamoto, Y. Nrf2 Activation Protects the Liver from Ischemia/Reperfusion Injury in Mice. Ann. Surg. 2014, 260, 118–127. [Google Scholar] [CrossRef] [Green Version]
- Xu, D.; Chen, L.; Chen, X.; Wen, Y.; Yu, C.; Yao, J.; Wu, H.; Wang, X.; Xia, Q.; Kong, X. The Triterpenoid CDDO-Imidazolide Ameliorates Mouse Liver Ischemia-Reperfusion Injury through Activating the Nrf2/HO-1 Pathway Enhanced Autophagy. Cell Death Dis. 2017, 8, e2983. [Google Scholar] [CrossRef] [Green Version]
- National Research Council (US) Committee for the Update of the Guide for the Care and Use of Laboratory Animals Guide for the Care and Use of Laboratory Animals. In The National Academies Collection: Reports Funded by National Institutes of Health, 8th ed.; National Academies Press: Washington, DC, USA, 2011; ISBN 978-0-309-15400-0.
- Kaufmann, S.H.; Blum, C.; Yamamoto, S. Crosstalk between Alpha/Beta T Cells and Gamma/Delta T Cells in Vivo: Activation of Alpha/Beta T-Cell Responses after Gamma/Delta T-Cell Modulation with the Monoclonal Antibody GL3. Proc. Natl. Acad. Sci. USA 1993, 90, 9620–9624. [Google Scholar] [CrossRef] [Green Version]
- Feng, M.; Li, G.; Qian, X.; Fan, Y.; Huang, X.; Zhang, F.; Lu, L. IL-17A-Producing NK Cells Were Implicated in Liver Injury Induced by Ischemia and Reperfusion. Int. Immunopharmacol. 2012, 13, 135–140. [Google Scholar] [CrossRef]
- Lentsch, A.B.; Yoshidome, H.; Cheadle, W.G.; Miller, F.N.; Edwards, M.J. Chemokine Involvement in Hepatic Ischemia/Reperfusion Injury in Mice: Roles for Macrophage Inflammatory Protein-2 and Kupffer Cells. Hepatology 1998, 27, 507–512. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, S.; Nakamura, S.; Koizumi, T.; Sakaguchi, S.; Baba, S.; Muro, H.; Fujise, Y. The Beneficial Effect of a Prostaglandin I2 Analog on Ischemic Rat Liver. Transplantation 1991, 52, 979–983. [Google Scholar] [CrossRef] [PubMed]
- Ouzounidis, N.; Giakoustidis, A.; Poutahidis, T.; Angelopoulou, K.; Iliadis, S.; Chatzigiagkos, A.; Zacharioudaki, A.; Angelopoulos, S.; Papalois, A.; Papanikolaou, V.; et al. Interleukin 18 Binding Protein Ameliorates Ischemia/Reperfusion-Induced Hepatic Injury in Mice. Liver Transplant. Liver Transplant. 2016, 22, 237–246. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pfaffl, M.W. A New Mathematical Model for Relative Quantification in Real-Time RT-PCR. Nucleic Acids Res. 2001, 29, e45. [Google Scholar] [CrossRef] [PubMed]
- Heinhuis, B.; Netea, M.G.; van den Berg, W.B.; Dinarello, C.A.; Joosten, L.A.B. Interleukin-32: A Predominantly Intracellular Proinflammatory Mediator That Controls Cell Activation and Cell Death. Cytokine 2012, 60, 321–327. [Google Scholar] [CrossRef]
- Funken, D.; Yu, Y.; Feng, X.; Imvised, T.; Gueler, F.; Prinz, I.; Madadi-Sanjani, O.; Ure, B.M.; Kuebler, J.F.; Klemann, C. Lack of Gamma Delta T Cells Ameliorates Inflammatory Response after Acute Intestinal Ischemia Reperfusion in Mice. Sci. Rep. 2021, 11, 18628. [Google Scholar] [CrossRef]
- Hochegger, K.; Schätz, T.; Eller, P.; Tagwerker, A.; Heininger, D.; Mayer, G.; Rosenkranz, A.R. Role of α/β and γ/δ T Cells in Renal Ischemia-Reperfusion Injury. Am. J. Physiol.-Ren. Physiol. 2007, 293, F741–F747. [Google Scholar] [CrossRef] [Green Version]
- Savransky, V.; Molls, R.R.; Burne-Taney, M.; Chien, C.-C.; Racusen, L.; Rabb, H. Role of the T-Cell Receptor in Kidney Ischemia-Reperfusion Injury. Kidney Int. 2006, 69, 233–238. [Google Scholar] [CrossRef] [Green Version]
- Gelderblom, M.; Weymar, A.; Bernreuther, C.; Velden, J.; Arunachalam, P.; Steinbach, K.; Orthey, E.; Arumugam, T.V.; Leypoldt, F.; Simova, O.; et al. Neutralization of the IL-17 Axis Diminishes Neutrophil Invasion and Protects from Ischemic Stroke. Blood 2012, 120, 3793–3802. [Google Scholar] [CrossRef] [Green Version]
- Eggenhofer, E.; Rovira, J.; Sabet-Baktach, M.; Groell, A.; Scherer, M.N.; Dahlke, M.-H.; Farkas, S.A.; Loss, M.; Koehl, G.E.; Lang, S.A.; et al. Unconventional RORγt+ T Cells Drive Hepatic Ischemia Reperfusion Injury. J. Immunol. 2013, 191, 480–487. [Google Scholar] [CrossRef] [Green Version]
- Shichita, T.; Sugiyama, Y.; Ooboshi, H.; Sugimori, H.; Nakagawa, R.; Takada, I.; Iwaki, T.; Okada, Y.; Iida, M.; Cua, D.J.; et al. Pivotal Role of Cerebral Interleukin-17-Producing GammadeltaT Cells in the Delayed Phase of Ischemic Brain Injury. Nat. Med. 2009, 15, 946–950. [Google Scholar] [CrossRef] [PubMed]
- Sutton, C.E.; Lalor, S.J.; Sweeney, C.M.; Brereton, C.F.; Lavelle, E.C.; Mills, K.H.G. Interleukin-1 and IL-23 Induce Innate IL-17 Production from Gammadelta T Cells, Amplifying Th17 Responses and Autoimmunity. Immunity 2009, 31, 331–341. [Google Scholar] [CrossRef] [Green Version]
- Lemasters, J.J.; Thurman, R.G. Reperfusion Injury after Liver Preservation for Transplantation. Annu. Rev. Pharmacol. Toxicol. 1997, 37, 327–338. [Google Scholar] [CrossRef] [PubMed]
- Hanschen, M.; Zahler, S.; Krombach, F.; Khandoga, A. Reciprocal Activation between CD4+ T Cells and Kupffer Cells during Hepatic Ischemia-Reperfusion. Transplantation 2008, 86, 710–718. [Google Scholar] [CrossRef] [PubMed]
- Khandoga, A.; Kessler, J.S.; Hanschen, M.; Khandoga, A.G.; Burggraf, D.; Reichel, C.; Hamann, G.F.; Enders, G.; Krombach, F. Matrix Metalloproteinase-9 Promotes Neutrophil and T Cell Recruitment and Migration in the Postischemic Liver. J. Leukoc. Biol. 2006, 79, 1295–1305. [Google Scholar] [CrossRef] [PubMed]
- Schofield, Z.V.; Woodruff, T.M.; Halai, R.; Wu, M.C.-L.; Cooper, M.A. Neutrophils—A Key Component of Ischemia-Reperfusion Injury. Shock 2013, 40, 463–470. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jaeschke, H.; Farhood, A. Neutrophil and Kupffer Cell-Induced Oxidant Stress and Ischemia-Reperfusion Injury in Rat Liver. Am. J. Physiol. 1991, 260, G355–G362. [Google Scholar] [CrossRef]
- Caldwell, C.; Okaya, T.; Martignoni, A.; Husted, T.; Schuster, R.; Lentsch, A. Divergent Functions of CD4+ T Lymphocytes in Acute Liver Inflammation and Injury after Ischemia-Reperfusion. Am. J. Physiol. Gastrointest. Liver Physiol. 2005, 289, G969–G976. [Google Scholar] [CrossRef]
- Liang, S.C.; Long, A.J.; Bennett, F.; Whitters, M.J.; Karim, R.; Collins, M.; Goldman, S.J.; Dunussi-Joannopoulos, K.; Williams, C.M.M.; Wright, J.F.; et al. An IL-17F/A Heterodimer Protein Is Produced by Mouse Th17 Cells and Induces Airway Neutrophil Recruitment. J. Immunol. 2007, 179, 7791–7799. [Google Scholar] [CrossRef] [Green Version]
- Liao, Y.; Xia, N.; Zhou, S.; Tang, T.; Yan, X.; Lv, B.-J.; Nie, S.; Wang, J.; Iwakura, Y.; Xiao, H.; et al. Interleukin-17A Contributes to Myocardial Ischemia/Reperfusion Injury by Regulating Cardiomyocyte Apoptosis and Neutrophil Infiltration. J. Am. Coll. Cardiol. 2012, 59, 420–429. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nguyen, C.T.; Furuya, H.; Das, D.; Marusina, A.I.; Merleev, A.A.; Ravindran, R.; Jalali, Z.; Khan, I.H.; Maverakis, E.; Adamopoulos, I.E. Peripheral Γδ T Cells Regulate Neutrophil Expansion and Recruitment in Experimental Psoriatic Arthritis. Arthritis Rheumatol. 2022, 74, 1524–1534. [Google Scholar] [CrossRef] [PubMed]
- Toth, B.; Alexander, M.; Daniel, T.; Chaudry, I.H.; Hubbard, W.J.; Schwacha, M.G. The Role of Gammadelta T Cells in the Regulation of Neutrophil-Mediated Tissue Damage after Thermal Injury. J. Leukoc. Biol. 2004, 76, 545–552. [Google Scholar] [CrossRef] [PubMed]
- Fábrega, E.; López-Hoyos, M.; San Segundo, D.; Casafont, F.; Pons-Romero, F. Changes in the Serum Levels of Interleukin-17/Interleukin-23 during Acute Rejection in Liver Transplantation. Liver Transplant. 2009, 15, 629–633. [Google Scholar] [CrossRef]
- Lu, L.; Li, G.; Rao, J.; Pu, L.; Yu, Y.; Wang, X.; Zhang, F. In Vitro Induced CD4(+)CD25(+)Foxp3(+) Tregs Attenuate Hepatic Ischemia-Reperfusion Injury. Int. Immunopharmacol. 2009, 9, 549–552. [Google Scholar] [CrossRef]
- Kuboki, S.; Sakai, N.; Tschöp, J.; Edwards, M.J.; Lentsch, A.B.; Caldwell, C.C. Distinct Contributions of CD4+ T Cell Subsets in Hepatic Ischemia/Reperfusion Injury. Am. J. Physiol.-Gastrointest. Liver Physiol. 2009, 296, G1054–G1059. [Google Scholar] [CrossRef] [Green Version]
- Wanner, G.A.; Ertel, W.; Müller, P.; Höfer, Y.; Leiderer, R.; Menger, M.D.; Messmer, K. Liver Ischemia and Reperfusion Induces a Systemic Inflammatory Response through Kupffer Cell Activation. Shock 1996, 5, 34–40. [Google Scholar] [CrossRef]
- Korn, T.; Bettelli, E.; Oukka, M.; Kuchroo, V.K. IL-17 and Th17 Cells. Annu. Rev. Immunol. 2009, 27, 485–517. [Google Scholar] [CrossRef]
- Littman, D.R.; Rudensky, A.Y. Th17 and Regulatory T Cells in Mediating and Restraining Inflammation. Cell 2010, 140, 845–858. [Google Scholar] [CrossRef] [Green Version]
- Rao, J.; Lu, L.; Zhai, Y. T Cells in Organ Ischemia Reperfusion Injury. Curr. Opin. Organ Transplant. 2014, 19, 115–120. [Google Scholar] [CrossRef] [Green Version]
- Zou, X.M.; Yagihashi, A.; Hirata, K.; Tsuruma, T.; Matsuno, T.; Tarumi, K.; Asanuma, K.; Watanabe, N. Downregulation of Cytokine-Induced Neutrophil Chemoattractant and Prolongation of Rat Liver Allograft Survival by Interleukin-10. Surg. Today 1998, 28, 184–191. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Li, C.; Ng, K.T.-P.; Liu, J.; Liu, H.; Zhang, W.; Xiao, F.; Li, X.; Lo, C.M.; Lu, L.; et al. IL-17a Exacerbates Hepatic Ischemia-Reperfusion Injury in Fatty Liver by Promoting Neutrophil Infiltration and Mitochondria-Driven Apoptosis. J. Leukoc. Biol. 2020, 108, 1603–1613. [Google Scholar] [CrossRef] [PubMed]
- Yoshidome, H.; Kato, A.; Edwards, M.J.; Lentsch, A.B. Interleukin-10 Suppresses Hepatic Ischemia/Reperfusion Injury in Mice: Implications of a Central Role for Nuclear Factor ΚB. Hepatology 1999, 30, 203–208. [Google Scholar] [CrossRef] [PubMed]
- Ricote, M.; Li, A.C.; Willson, T.M.; Kelly, C.J.; Glass, C.K. The Peroxisome Proliferator-Activated Receptor-Gamma Is a Negative Regulator of Macrophage Activation. Nature 1998, 391, 79–82. [Google Scholar] [CrossRef]
- Bouhlel, M.A.; Derudas, B.; Rigamonti, E.; Dièvart, R.; Brozek, J.; Haulon, S.; Zawadzki, C.; Jude, B.; Torpier, G.; Marx, N.; et al. PPARgamma Activation Primes Human Monocytes into Alternative M2 Macrophages with Anti-Inflammatory Properties. Cell Metab. 2007, 6, 137–143. [Google Scholar] [CrossRef] [Green Version]
- Linares, I.; Farrokhi, K.; Echeverri, J.; Kaths, J.M.; Kollmann, D.; Hamar, M.; Urbanellis, P.; Ganesh, S.; Adeyi, O.A.; Yip, P.; et al. PPAR-Gamma Activation Is Associated with Reduced Liver Ischemia-Reperfusion Injury and Altered Tissue-Resident Macrophages Polarization in a Mouse Model. PLoS ONE 2018, 13, e0195212. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer | Sequence (5′-3′) | GenBank Access No | Positions | Amplicon Size (bp) | Primer Conc. (nM) | Annealing T (°C) |
|---|---|---|---|---|---|---|
| IL1-β-F | TGGACCTTCCAGGATGAGGACA | NM_008361.4 | 331–352 | 148 | 100 | 61 |
| IL1-β-R | GTTCATCTCGGAGCCTGTAGTG | 457–487 | ||||
| IL6-F | TACCACTTCACAAGTCGGAGGC | NM_031168.2 | 189–209 | 144 | 250 | 60 |
| IL6-R | CTGCAAGTGCATCATCGTTGTTC | 315–332 | ||||
| IL10-F | TCAGCCAGGTGAAGACTTTCT | NM_010548.2 | 210–230 | 147 | 200 | 63 |
| ILA10-R | GGGGCATCACTTCTACCAGG | 337–356 | ||||
| PPAR-γ-F | GTACTGTCGGTTTCAGAAGTGCC | NM_011146 | 618–640 | 104 | 250 | 60 |
| PPAR-γ-R | ATCTCCGCCAACAGCTTCTCCT | 698–719 | ||||
| NRF2-F | CAGCATAGAGCAGGACATGGAG | NM_010902 | 765–786 | 107 | 250 | 60 |
| NRF2-R | GAACAGCGGTAGTATCAGCCAG | 850–871 | ||||
| TNF-α-F | GTCCCCAAAGGGATGAGAAG | NM_013693.3 | 337–356 | 134 | 300 | 59 |
| TNF-α-R | CACTTGGTGGTTTGCTACGA | 451–470 | ||||
| TGF-β-F | GGAGAGCCCTGGATACCAAC | NM_011577.2 | 1697–1716 | 149 | 250 | 59 |
| TGF-β-R | GCAGGGTCCCAGACAGAAG | 1827–1845 | ||||
| GAPDH-F | CATCAAATGGGGTGAGGCCG | NM_001289726.1 | 338–357 | 149 | 250 | 63 |
| GAPDH-R | CCATCACAAACATGGGGGCA | 467–486 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Al Mogrampi, S.; Boumpoureka, C.; Afaloniati, H.; Lagou, M.; Angelopoulou, K.; Anestakis, D.; Tampouratzi, Z.G.; Iliadis, S.; Antoniadis, N.; Giakoustidis, A.; et al. Inhibition of γδ-TcR or IL17a Reduces T-Cell and Neutrophil Infiltration after Ischemia/Reperfusion Injury in Mouse Liver. J. Clin. Med. 2023, 12, 1751. https://doi.org/10.3390/jcm12051751
Al Mogrampi S, Boumpoureka C, Afaloniati H, Lagou M, Angelopoulou K, Anestakis D, Tampouratzi ZG, Iliadis S, Antoniadis N, Giakoustidis A, et al. Inhibition of γδ-TcR or IL17a Reduces T-Cell and Neutrophil Infiltration after Ischemia/Reperfusion Injury in Mouse Liver. Journal of Clinical Medicine. 2023; 12(5):1751. https://doi.org/10.3390/jcm12051751
Chicago/Turabian StyleAl Mogrampi, Saant, Christina Boumpoureka, Hara Afaloniati, Maria Lagou, Katerina Angelopoulou, Doxakis Anestakis, Zoi Gerasimina Tampouratzi, Stavros Iliadis, Nikolaos Antoniadis, Alexandros Giakoustidis, and et al. 2023. "Inhibition of γδ-TcR or IL17a Reduces T-Cell and Neutrophil Infiltration after Ischemia/Reperfusion Injury in Mouse Liver" Journal of Clinical Medicine 12, no. 5: 1751. https://doi.org/10.3390/jcm12051751
APA StyleAl Mogrampi, S., Boumpoureka, C., Afaloniati, H., Lagou, M., Angelopoulou, K., Anestakis, D., Tampouratzi, Z. G., Iliadis, S., Antoniadis, N., Giakoustidis, A., Papalois, A., Papadopoulos, V., Poutahidis, T., & Giakoustidis, D. (2023). Inhibition of γδ-TcR or IL17a Reduces T-Cell and Neutrophil Infiltration after Ischemia/Reperfusion Injury in Mouse Liver. Journal of Clinical Medicine, 12(5), 1751. https://doi.org/10.3390/jcm12051751