
Vascular Progenitor Cells: From Cancer to Tissue Repair

Abstract
:1. Introduction
2. Vascular Progenitor Cells
2.1. Endothelial Progenitor Cells
2.2. Smooth Muscle Progenitor Cells
2.3. Mesangiogenic Progenitor Cells
2.4. Pericytes
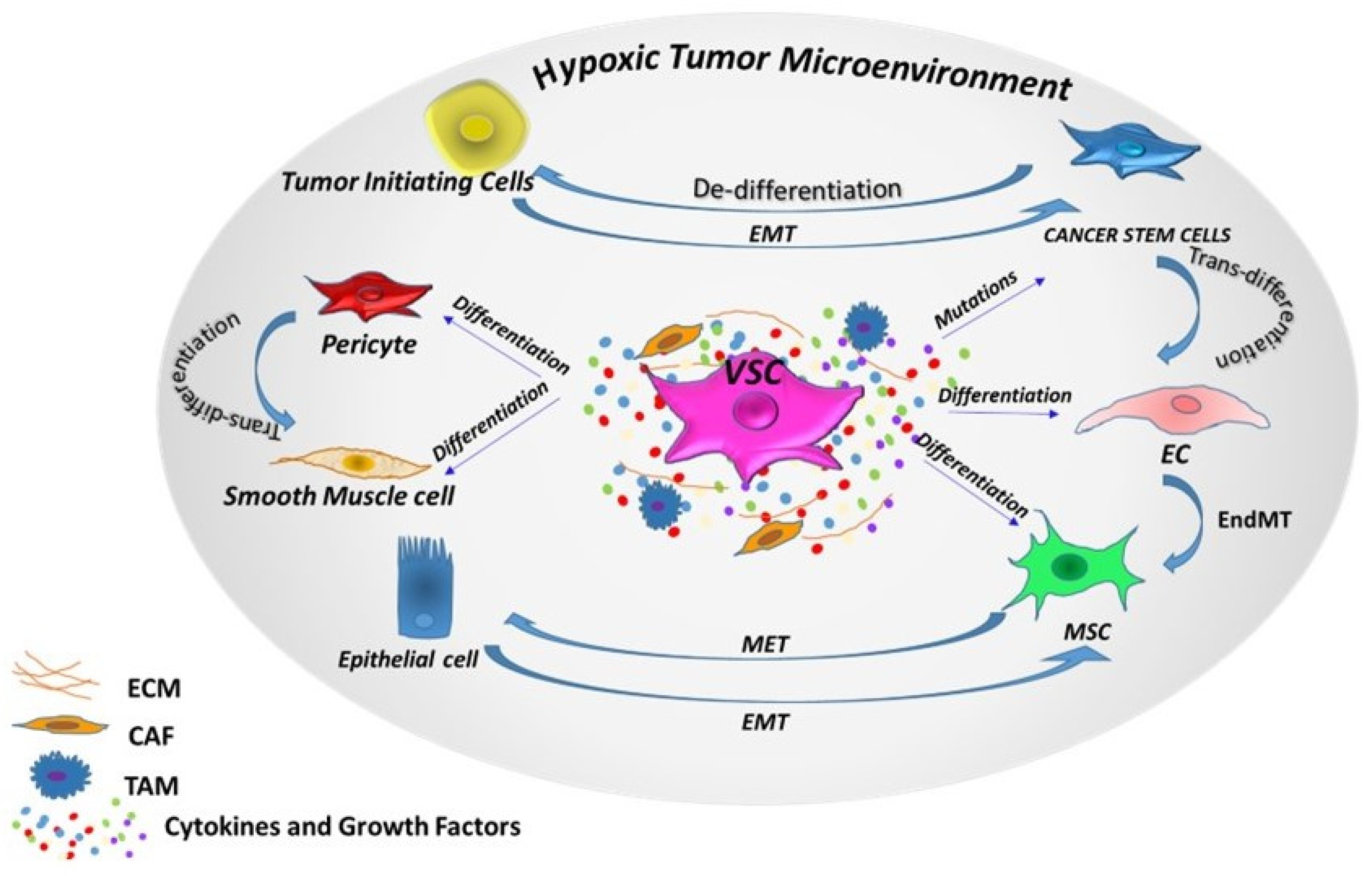
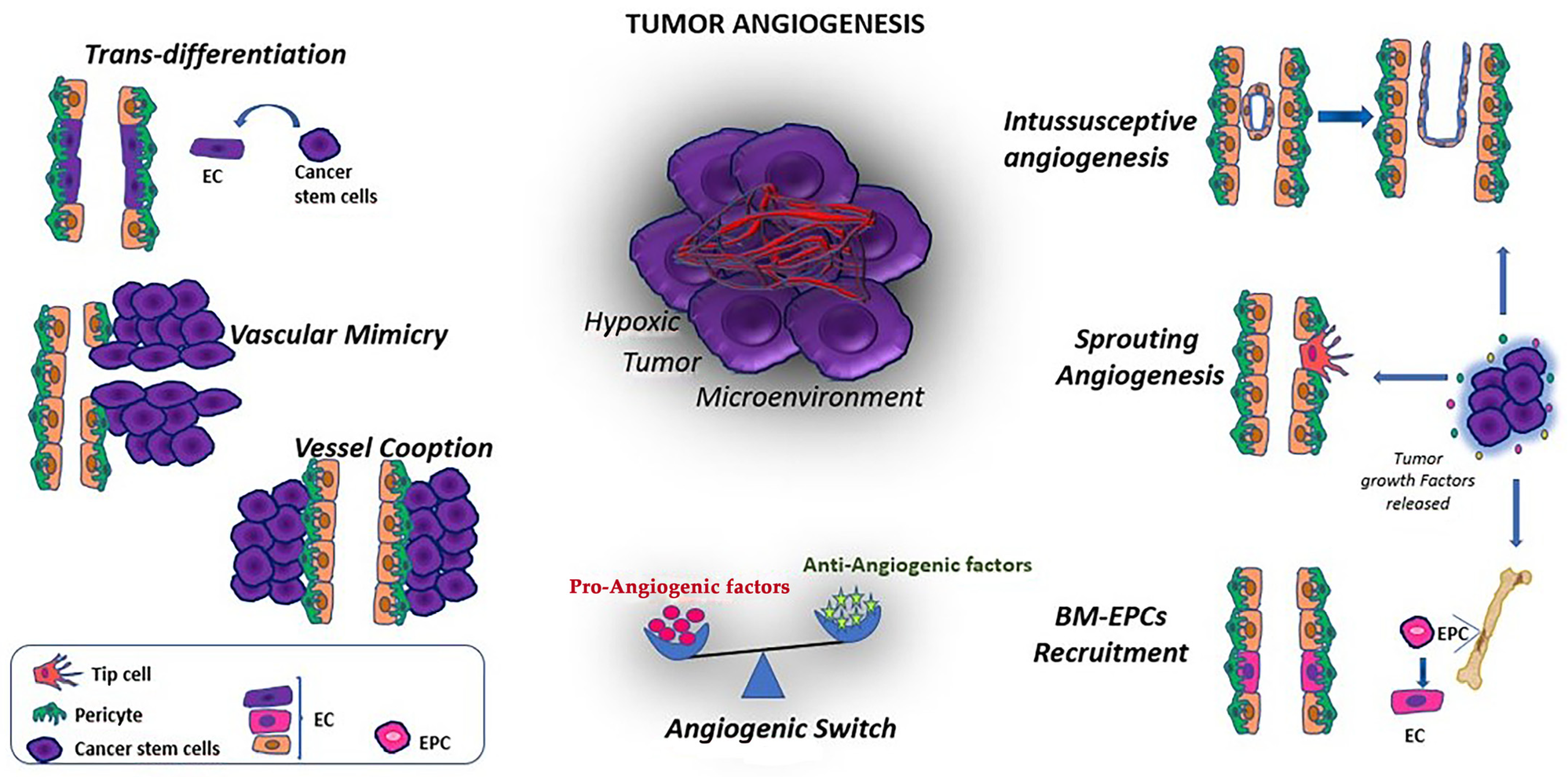
3. Vascular Progenitor Cells in Cancers
4. Future Perspectives
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Tao, J.; Cao, X.; Yu, B.; Qu, A. Vascular Stem/Progenitor Cells in Vessel Injury and Repair. Front. Cardiovasc. Med. 2022, 9, 845070. [Google Scholar] [CrossRef] [PubMed]
- da Silva, C.; Guedes, I.H.L.; de Lima, J.C.S.; Sobrinho, J.; Dos Santos, A.A. Responses triggered by the immune system in hypertensive conditions and repercussions on target-organ damage: A review. Curr. Cardiol. Rev. 2022, 19, 56–65. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Xu, L.; Zuo, Y.X.; Chang, X.Q.; Chi, H.T. Potential intervention target of atherosclerosis: Ferroptosis (Review). Mol. Med. Rep. 2022, 26, 12859. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, S.; Hahn, Y.; Silverstein, B.; Singh, M.; Fleitz, A.; Van, J.; Chen, H.; Liang, Q. Lysosomal dysfunction in diabetic cardiomyopathy. Front. Aging 2023, 4, 1113200. [Google Scholar] [CrossRef]
- Ramsey, D.J.; Kwan, J.T.; Sharma, A. Keeping an eye on the diabetic foot: The connection between diabetic eye disease and wound healing in the lower extremity. World J. Diabetes 2022, 13, 1035–1048. [Google Scholar] [CrossRef]
- Li, S.; Dinh, H.T.P.; Matsuyama, Y.; Sato, K.; Yamagishi, S. Molecular Mechanisms in the Vascular and Nervous Systems following Traumatic Spinal Cord Injury. Life 2022, 13, 9. [Google Scholar] [CrossRef]
- Kohli, S.; Shahzad, K.; Jouppila, A.; Holthöfer, H.; Isermann, B.; Lassila, R. Thrombosis and Inflammation—A Dynamic Interplay and the Role of Glycosaminoglycans and Activated Protein C. Front. Cardiovasc. Med. 2022, 9, 866751. [Google Scholar] [CrossRef]
- Jiang, Y.; Huang, J.; Xia, Y.; Sun, Z.; Hu, P.; Wang, D.; Liu, Y.; Tao, T.; Liu, Y. Hypoxia activates GPR146 which participates in pulmonary vascular remodeling by promoting pyroptosis of pulmonary artery endothelial cells. Eur. J. Pharmacol. 2023, 941, 175502. [Google Scholar] [CrossRef]
- Hida, K.; Maishi, N.; Takeda, R.; Hida, Y. The Roles of Tumor Endothelial Cells in Cancer Metastasis. In Metastasis; Sergi, C.M., Ed.; Exon Publications: Brisbane, Australia, 2022. [Google Scholar]
- Sirivella, M.L.; Rahinj, G.B.; Chauhan, H.S.; Satyanarayana, M.V.; Ramanan, L. Numerical Methodology to Evaluate Trackability and Pushability of PTCA Balloon Catheter. Cardiovasc. Eng. Technol. 2023, 1–16. [Google Scholar] [CrossRef]
- Ejiri, K.; Ogawa, A.; Shimokawahara, H.; Matsubara, H. Treatment of Vascular Injury During Balloon Pulmonary Angioplasty in Patients with Chronic Thromboembolic Pulmonary Hypertension. JACC Asia 2022, 2, 831–842. [Google Scholar] [CrossRef]
- Dight, J.; Zhao, J.; Styke, C.; Khosrotehrani, K.; Patel, J. Resident vascular endothelial progenitor definition and function: The age of reckoning. Angiogenesis 2021, 25, 15–33. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.; Li, Y.; Yang, Y.; Li, P. The Microenvironment That Regulates Vascular Wall Stem/Progenitor Cells in Vascular Injury and Repair. BioMed Res. Int. 2022, 2022, 9377965. [Google Scholar] [CrossRef] [PubMed]
- Medina, R.J.; Barber, C.L.; Sabatier, F.; Dignat-George, F.; Melero-Martin, J.M.; Khosrotehrani, K.; Ohneda, O.; Randi, A.M.; Chan, J.K.Y.; Yamaguchi, T.; et al. Endothelial progenitors: A consensus statement on nomenclature. Stem Cells Transl. Med. 2017, 6, 1316–1320. [Google Scholar] [CrossRef]
- Dzobo, K.; Senthebane, D.A.; Dandara, C. The Tumor Microenvironment in Tumorigenesis and Therapy Resistance Revisited. Cancers 2023, 15, 376. [Google Scholar] [CrossRef] [PubMed]
- Lin, C.S.; Lue, T.F. Defining vascular stem cells. Stem Cells Dev. 2013, 22, 1018–1026. [Google Scholar] [CrossRef] [Green Version]
- Carmeliet, P. Angiogenesis in health and disease. Nat. Med. 2003, 9, 653–660. [Google Scholar] [CrossRef] [PubMed]
- Chopra, H.; Hung, M.K.; Kwong, D.L.; Zhang, C.F.; Pow, E.H.N. Insights into endothelial progenitor cells: Origin, classification, potentials, and prospects. Stem Cells Int. 2018, 2018, 9847015. [Google Scholar] [CrossRef] [Green Version]
- Troidl, K.; Schaper, W. Arteriogenesis versus angiogenesis in peripheral artery disease. Diabetes/Metab. Res. Rev. 2012, 28, 27–29. [Google Scholar] [CrossRef]
- Simons, M. Angiogenesis: Where do we stand now? Circulation 2005, 111, 1556–1566. [Google Scholar] [CrossRef] [Green Version]
- Liu, M.; Gomez, D. Smooth Muscle Cell Phenotypic Diversity. Arterioscler. Thromb. Vasc. Biol. 2019, 39, 1715–1723. [Google Scholar] [CrossRef]
- Madonna, R.; De Caterina, R. Circulating endothelial progenitor cells: Do they live up to their name? Vascul. Pharmacol. 2015, 67–69, 2–5. [Google Scholar] [CrossRef] [PubMed]
- Craig, D.J.; James, A.W.; Wang, Y.; Tavian, M.; Crisan, M.; Péault, B.M. Blood Vessel Resident Human Stem Cells in Health and Disease. STEM CELLS Transl. Med. 2022, 11, 35–43. [Google Scholar] [CrossRef] [PubMed]
- Mangialardi, G.; Cordaro, A.; Madeddu, P. The bone marrow pericyte: An orchestrator of vascular niche. Regen. Med. 2016, 11, 883–895. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Psaltis, P.J.; Simari, R.D. Vascular Wall Progenitor Cells in Health and Disease. Circ. Res. 2015, 116, 1392–1412. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lugano, R.; Ramachandran, M.; Dimberg, A. Tumor angiogenesis: Causes, consequences, challenges and opportu-nities. Cell. Mol. Life Sci. 2020, 77, 1745–1770. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Asahara, T.; Murohara, T.; Sullivan, A.; Silver, M.; van der Zee, R.; Li, T.; Witzenbichler, B.; Schatteman, G.; Isner, J.M. Isolation of putative progenitor endothelial cells for angiogenesis. Science 1997, 275, 964–966. [Google Scholar] [CrossRef]
- Moccia, F.; Zuccolo, E.; Poletto, V.; Cinelli, M.; Bonetti, E.; Guerra, G.; Rosti, V. Endothelial progenitor cells support tumour growth and metastatisation: Implications for the resistance to anti-angiogenic therapy. Tumor Biol. 2015, 36, 6603–6614. [Google Scholar] [CrossRef]
- Choi, K.; Kennedy, M.; Kazarov, A.; Papadimitriou, J.C.; Keller, G. A common precursor for hematopoietic and en-dothelial cells. Development 1998, 125, 725–732. [Google Scholar] [CrossRef]
- Asahara, T.; Kawamoto, A. Endothelial progenitor cells for postnatal vasculogenesis. Am. J. Physiol. Physiol. 2004, 287, C572–C579. [Google Scholar] [CrossRef] [Green Version]
- Barone, C.; Orsenigo, R.; Meneveri, R.; Brunelli, S.; Azzoni, E. One Size Does Not Fit All: Heterogeneity in Develop-mental Hematopoiesis. Cells 2022, 11, 1061. [Google Scholar] [CrossRef]
- Zhu, Q.; Gao, P.; Tober, J.; Bennett, L.; Chen, C.; Uzun, Y.; Li, Y.; Howell, E.D.; Mumau, M.; Yu, W.; et al. Developmental trajectory of prehematopoietic stem cell formation from endothelium. Blood 2020, 136, 845–856. [Google Scholar] [CrossRef] [PubMed]
- Zhao, S.; Feng, S.; Tian, Y.; Wen, Z. Hemogenic and aortic endothelium arise from a common hemogenic angioblast precursor and are specified by the Etv2 dosage. Proc. Natl. Acad. Sci. USA 2022, 119, e2119051119. [Google Scholar] [CrossRef] [PubMed]
- Baltrunaite, K.; Craig, M.P.; Desai, S.P.; Chaturvedi, P.; Pandey, R.N.; Hegde, R.S.; Sumanas, S. ETS transcription factors Etv2 and Fli1b are required for tumor angiogenesis. Angiogenesis 2017, 20, 307–323. [Google Scholar] [CrossRef] [Green Version]
- Friedrich, E.B.; Walenta, K.; Scharlau, J.; Nickenig, G.; Werner, N. CD34-/CD133+/VEGFR-2+ endothelial progenitor cell subpopulation with poten vasoregenerative capacities. Circ. Res. 2006, 98, e20–e25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.; Wang, R.; Jiang, L.; Xu, Q.; Guo, X. Endothelial repair by stem and progenitor cells. J. Mol. Cell. Cardiol. 2021, 163, 133–146. [Google Scholar] [CrossRef] [PubMed]
- Ingram, D.A.; Mead, L.E.; Moore, D.B.; Woodard, W.; Fenoglio, A.; Yoder, M.C. Vessel wall–derived endothelial cells rapidly proliferate because they contain a complete hierarchy of endothelial progenitor cells. Blood 2005, 105, 2783–2786. [Google Scholar] [CrossRef] [Green Version]
- Naito, H.; Kidoya, H.; Sakimoto, S.; Wakabayashi, T.; Takakura, N. Identification and characterization of a resident vascular stem/progenitor cell population in preexisting blood vessels. EMBO J. 2011, 31, 842–855. [Google Scholar] [CrossRef] [Green Version]
- Ratajczak, M.Z.; Shin, D.-M.; Liu, R.; Mierzejewska, K.; Ratajczak, J.; Kucia, M.; Zuba-Surma, E.K. Very small embryonic/epiblast-like stem cells (VSELs) and their potential role in aging and organ rejuvenation—An update and comparison to other primitive small stem cells isolated from adult tissues. Aging 2012, 4, 235–246. [Google Scholar] [CrossRef]
- Ratajczak, M.Z.; Ratajczak, J.; Suszynska, M.; Miller, D.M.; Kucia, M.; Shin, D.-M. A Novel View of the Adult Stem Cell Compartment from the Perspective of a Quiescent Population of Very Small Embryonic-Like Stem Cells. Circ. Res. 2017, 120, 166–178. [Google Scholar] [CrossRef] [Green Version]
- Schwarz, T.M.; Leicht, S.F.; Radic, T.; Rodriguez-Arabaolaza, I.; Hermann, P.C.; Berger, F.; Saif, J.; Böcker, W.; Ellwart, J.W.; Aicher, A.; et al. Vascular incorporation of endothelial colony-forming cells is essential for functional recovery of murine ischemic tissue following cell therapy. Arter. Thromb. Vasc. Biol. 2012, 32, e13–e21. [Google Scholar] [CrossRef] [Green Version]
- Kang, K.T.; Lin, R.Z.; Kuppermann, D.; Melero-Martin, J.M.; Bischoff, J. Endothelial colony forming cells and mesen-chymal progenitor cells form blood vessels and increase blood flow in ischemic muscle. Sci. Rep. 2017, 7, 770. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, G.; Jacquet, L.; Karamariti, E.; Xu, Q. Origin and differentiation of vascular smooth muscle cells. J. Physiol. 2015, 593, 3013–3030. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burggren, W.W. Phenotypic Switching Resulting from Developmental Plasticity: Fixed or Reversible? Front. Physiol. 2019, 10, 1634. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sata, M.; Saiura, A.; Kunisato, A.; Tojo, A.; Okada, S.; Tokuhisa, T.; Hirai, H.; Makuuchi, M.; Hirata, Y.; Nagai, R. Hematopoietic stem cells differentiate into vascular cells that participate in the pathogenesis of atherosclerosis. Nat. Med. 2002, 8, 403–409. [Google Scholar] [CrossRef]
- Bentzon, J.F.; Weile, C.; Sondergaard, C.S.; Hindkjaer, J.; Kassem, M.; Falk, E. Smooth Muscle Cells in Atherosclerosis Originate from the Local Vessel Wall and Not Circulating Progenitor Cells in ApoE Knockout Mice. Arter. Thromb. Vasc. Biol. 2006, 26, 2696–2702. [Google Scholar] [CrossRef] [Green Version]
- Lu, W.; Li, X. Vascular stem/progenitor cells: Functions and signaling pathways. Cell. Mol. Life Sci. 2017, 75, 859–869. [Google Scholar] [CrossRef]
- Hu, Y.; Zhang, Z.; Torsney, E.; Afzal, A.R.; Davison, F.; Metzler, B.; Xu, Q. Abundant progenitor cells in the adventitia contribute to atherosclerosis of vein grafts in ApoE-deficient mice. J. Clin. Investig. 2004, 113, 1258–1265. [Google Scholar] [CrossRef] [Green Version]
- Kramann, R.; Goettsch, C.; Wongboonsin, J.; Iwata, H.; Schneider, R.K.; Kuppe, C.; Kaesler, N.; Chang-Panesso, M.; Machado, F.G.; Gratwohl, S.; et al. Adventitial MSC-like Cells Are Progenitors of Vascular Smooth Muscle Cells and Drive Vascular Calcification in Chronic Kidney Disease. Cell Stem Cell 2016, 19, 628–642. [Google Scholar] [CrossRef] [Green Version]
- Majesky, M.W.; Horita, H.; Ostriker, A.; Lu, S.; Regan, J.N.; Bagchi, A.; Dong, X.R.; Poczobutt, J.; Nemenoff, R.A.; Weiser-Evans, M.C. Differentiated Smooth Muscle Cells Generate a Subpopulation of Res-ident Vascular Progenitor Cells in the Adventitia Regulated by Klf4. Circ. Res. 2017, 120, 296–311. [Google Scholar] [CrossRef]
- Tang, J.; Wang, H.; Huang, X.; Li, F.; Zhu, H.; Li, Y.; He, L.; Zhang, H.; Pu, W.; Liu, K.; et al. Arterial Sca1+ Vascular Stem Cells Generate De Novo Smooth Muscle for Artery Repair and Regeneration. Cell Stem Cell 2019, 26, 81–96. [Google Scholar] [CrossRef]
- Zhang, L.; Bhaloo, S.I.; Chen, T.; Zhou, B.; Xu, Q. Role of Resident Stem Cells in Vessel Formation and Arteriosclerosis. Circ. Res. 2018, 122, 1608–1624. [Google Scholar] [CrossRef] [PubMed]
- Frismantiene, A.; Philippova, M.; Erne, P.; Resink, T.J. Smooth muscle cell-driven vascular diseases and molecular mechanisms of VSMC plasticity. Cell. Signal. 2018, 52, 48–64. [Google Scholar] [CrossRef] [PubMed]
- Passman, J.N.; Dong, X.R.; Wu, S.P.; Maguire, C.T.; Hogan, K.A.; Bautch, V.L.; Majesky, M.W. A sonic hedgehog signaling domain in the arterial adventitia supports res-ident Sca1+ smooth muscle progenitor cells. Proc. Natl. Acad. Sci. USA 2008, 105, 9349–9354. [Google Scholar] [CrossRef] [Green Version]
- Yamashita, J.; Itoh, H.; Hirashima, M.; Ogawa, M.; Nishikawa, S.; Yurugi, T.; Naito, M.; Nakao, K.; Nishikawa, S.-I. Flk1-positive cells derived from embryonic stem cells serve as vascular progenitors. Nature 2000, 408, 92–96. [Google Scholar] [CrossRef] [PubMed]
- Chappell, J.; Harman, J.L.; Narasimhan, V.M.; Yu, H.; Foote, K.; Simons, B.D.; Bennett, M.R.; Jørgensen, H.F. Extensive Proliferation of a Subset of Differentiated, yet Plastic, Medial Vascular Smooth Muscle Cells Contributes to Neointimal Formation in Mouse Injury and Atherosclerosis Models. Circ. Res. 2016, 119, 1313–1323. [Google Scholar] [CrossRef] [PubMed]
- In’t Anker, P.S.; Scherjon, S.A.; Kleijburg-van der Keur, C.; de Groot-Swings, G.M.; Claas, F.H.; Fibbe, W.E.; Kanhai, H.H. Isolation of mesenchymal stem cells of fetal or ma-ternal origin from human placenta. Stem Cells 2004, 22, 1338–1345. [Google Scholar] [CrossRef]
- Barachini, S.; Trombi, L.; Danti, S.; D’Alessandro, D.; Battolla, B.; Legitimo, A.; Nesti, C.; Mucci, I.; D′acunto, M.; Cascone, M.G.; et al. Morpho-Functional Characterization of Human Mesenchymal Stem Cells from Umbilical Cord Blood for Potential Uses in Regenerative Medicine. Stem Cells Dev. 2009, 18, 293–306. [Google Scholar] [CrossRef]
- Nesti, C.; Pardini, C.; Barachini, S.; D’Alessandro, D.; Siciliano, G.; Murri, L.; Petrini, M.; Vaglini, F. Human dental pulp stem cells protect mouse dopaminergic neurons against MPP+ or rotenone. Brain Res. 2011, 1367, 94–102. [Google Scholar] [CrossRef]
- Barachini, S.; Danti, S.; Pacini, S.; D’Alessandro, D.; Carnicelli, V.; Trombi, L.; Moscato, S.; Mannari, C.; Cei, S.; Petrini, M. Plasticity of human dental pulp stromal cells with bioengineering platforms: A versatile tool for regenerative medicine. Micron 2014, 67, 155–168. [Google Scholar] [CrossRef]
- Sacchetti, B.; Funari, A.; Michienzi, S.; Di Cesare, S.; Piersanti, S.; Saggio, I.; Tagliafico, E.; Ferrari, S.; Robey, P.G.; Riminucci, M.; et al. Self-Renewing Osteoprogenitors in Bone Marrow Sinusoids Can Organize a Hematopoietic Microenvironment. Cell 2007, 131, 324–336. [Google Scholar] [CrossRef] [Green Version]
- Tormin, A.; Li, O.; Brune, J.C.; Walsh, S.; Schütz, B.; Ehinger, M.; Ditzel, N.; Kassem, M.; Scheding, S. CD146 expression on primary nonhematopoietic bone marrow stem cells is cor-related with in situ localization. Blood 2011, 117, 5067–5077. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Campagnolo, P.; Cesselli, D.; Zen, A.A.H.; Beltrami, A.P.; Kraenkel, N.; Katare, R.; Angelini, G.; Emanueli, C.; Madeddu, P. Human Adult Vena Saphena Contains Perivascular Progenitor Cells Endowed with Clonogenic and Proangiogenic Potential. Circulation 2010, 121, 1735–1745. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maacha, S.; Sidahmed, H.; Jacob, S.; Gentilcore, G.; Calzone, R.; Grivel, J.-C.; Cugno, C. Paracrine Mechanisms of Mesenchymal Stromal Cells in Angiogenesis. Stem Cells Int. 2020, 2020, 4356359. [Google Scholar] [CrossRef] [PubMed]
- Petrini, M.; Pacini, S.; Trombi, L.; Fazzi, R.; Montali, M.; Ikehara, S.; Abraham, N.G. Identification and Purification of Mesodermal Progenitor Cells from Human Adult Bone Marrow. Stem Cells Dev. 2009, 18, 857–866. [Google Scholar] [CrossRef]
- Barachini, S.; Montali, M.; Panvini, F.M.; Carnicelli, V.; Gatti, G.L.; Piolanti, N.; Bonicoli, E.; Scaglione, M.; Buda, G.; Parchi, P.D. Mesangiogenic Progenitor Cells Are Tissue Specific and Cannot Be Isolated from Adipose Tissue or Umbilical Cord Blood. Front. Cell Dev. Biol. 2021, 9, 669381. [Google Scholar] [CrossRef]
- Barachini, S.; Pacini, S.; Montali, M.; Panvini, F.M.; Carnicelli, V.; Piolanti, N.; Bonicoli, E.; Scaglione, M.; Parchi, P.D. Mesangiogenic Progenitor Cells and musculoskeletal tissue regeneration: Differences between adipose-derived and bone marrow-derived cells? J. Biol. Regul. Homeost. Agents 2020, 34, 33–38. [Google Scholar]
- Montali, M.; Barachini, S.; Panvini, F.M.; Carnicelli, V.; Fulceri, F.; Petrini, I.; Pacini, S. Growth Factor Content in Human Sera Affects the Isolation of Mesangiogenic Progenitor Cells (MPCs) from Human Bone Marrow. Front. Cell Dev. Biol. 2016, 4, 114. [Google Scholar] [CrossRef] [Green Version]
- Montali, M.; Barachini, S.; Pacini, S.; Panvini, F.M.; Petrini, M. Isolating Mesangiogenic Progenitor Cells (MPCs) from Human Bone Marrow. J. Vis. Exp. 2016, 15. [Google Scholar] [CrossRef] [Green Version]
- Pacini, S.; Carnicelli, V.; Trombi, L.; Montali, M.; Fazzi, R.; Lazzarini, E.; Giannotti, S.; Petrini, M. Constitutive Expression of Pluripotency-Associated Genes in Mesodermal Progenitor Cells (MPCs). PLoS ONE 2010, 5, e9861. [Google Scholar] [CrossRef]
- Fazzi, R.; Pacini, S.; Carnicelli, V.; Trombi, L.; Montali, M.; Lazzarini, E.; Petrini, M. Mesodermal progenitor cells (MPCs) differentiate into mesenchymal stromal cells (MSCs) by activation of Wnt5/calmodulin signalling pathway. PLoS ONE 2011, 6, e25600. [Google Scholar] [CrossRef] [Green Version]
- Gentry, T.; Foster, S.; Winstead, L.; Deibert, E.; Fiordalisi, M.; Balber, A. Simultaneous isolation of human BM hematopoietic, endothelial and mesenchymal progenitor cells by flow sorting based on aldehyde dehydrogenase activity: Implications for cell therapy. Cytotherapy 2007, 9, 259–274. [Google Scholar] [CrossRef] [PubMed]
- Roehrich, M.-E.; Spicher, A.; Milano, G.; Vassalli, G. Characterization of Cardiac-Resident Progenitor Cells Expressing High Aldehyde Dehydrogenase Activity. BioMed Res. Int. 2013, 2013, 503047. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nagano, M.; Yamashita, T.; Hamada, H.; Ohneda, K.; Kimura, K.-I.; Nakagawa, T.; Shibuya, M.; Yoshikawa, H.; Ohneda, O. Identification of functional endothelial progenitor cells suitable for the treatment of ischemic tissue using human umbilical cord blood. Blood 2007, 110, 151–160. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Montali, M.; Panvini, F.M.; Barachini, S.; Ronca, F.; Carnicelli, V.; Mazzoni, S.; Petrini, I.; Pacini, S. Human adult mesangiogenic progenitor cells reveal an early angiogenic potential, which is lost after mesengenic differentiation. Stem Cell Res. Ther. 2017, 8, 106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pacini, S.; Barachini, S.; Montali, M.; Carnicelli, V.; Fazzi, R.; Parchi, P.; Petrini, M. Mesangiogenic Progenitor Cells Derived from One Novel CD64(bright)CD31(bright)CD14(neg) Population in Human Adult Bone Marrow. Stem Cells Dev. 2016, 25, 661–673. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suzuki, S.; Namiki, J.; Shibata, S.; Mastuzaki, Y.; Okano, H. The Neural Stem/Progenitor Cell Marker Nestin Is Expressed in Proliferative Endothelial Cells, but Not in Mature Vasculature. J. Histochem. Cytochem. 2010, 58, 721–730. [Google Scholar] [CrossRef] [Green Version]
- Panvini, F.M.; Pacini, S.; Montali, M.; Barachini, S.; Mazzoni, S.; Morganti, R.; Ciancia, E.M.; Carnicelli, V.; Petrini, M. High NESTIN Expression Marks the Endosteal Capillary Network in Human Bone Marrow. Front. Cell Dev. Biol. 2020, 8, 596452. [Google Scholar] [CrossRef] [PubMed]
- Méndez-Ferrer, S.; Michurina, T.V.; Ferraro, F.; Mazloom, A.R.; MacArthur, B.D.; Lira, S.A.; Scadden, D.T.; Ma’Ayan, A.; Enikolopov, G.N.; Frenette, P.S. Mesenchymal and haematopoietic stem cells form a unique bone marrow niche. Nature 2010, 466, 829–834. [Google Scholar] [CrossRef] [Green Version]
- Kunisaki, Y.; Bruns, I.; Scheiermann, C.; Ahmed, J.; Pinho, S.; Zhang, D.; Mizoguchi, T.; Wei, Q.; Lucas, D.; Ito, K.; et al. Arteriolar niches maintain haematopoietic stem cell quiescence. Nature 2013, 502, 637–643. [Google Scholar] [CrossRef] [Green Version]
- Baccin, C.; Al-Sabah, J.; Velten, L.; Helbling, P.M.; Grünschläger, F.; Hernández-Malmierca, P.; Nombela-Arrieta, C.; Steinmetz, L.M.; Trumpp, A.; Haas, S. Combined single-cell and spatial transcriptomics reveal the molecular, cellular and spatial bone marrow niche organization. Nat. Cell Biol. 2020, 22, 38–48. [Google Scholar] [CrossRef]
- Dabravolski, S.A.; Markin, A.M.; Andreeva, E.R.; Eremin, I.I.; Orekhov, A.N.; Melnichenko, A.A. Emerging role of pericytes in therapy of cardiovascular diseases. Biomed. Pharmacother. 2022, 156, 113928. [Google Scholar] [CrossRef] [PubMed]
- Avolio, E.; Mangialardi, G.; Slater, S.C.; Alvino, V.V.; Gu, Y.; Cathery, W.; Beltrami, A.P.; Katare, R.; Heesom, K.; Caputo, M.; et al. Secreted Protein Acidic and Cysteine Rich Matricellular Protein is En-riched in the Bioactive Fraction of the Human Vascular Pericyte Secretome. Antioxid. Redox. Signal. 2021, 34, 1151–1164. [Google Scholar] [CrossRef] [PubMed]
- Omorphos, N.P.; Gao, C.; Tan, S.S.; Sangha, M.S. Understanding angiogenesis and the role of angiogenic growth factors in the vascularisation of engineered tissues. Mol. Biol. Rep. 2021, 48, 941–950. [Google Scholar] [CrossRef]
- Sweeney, M.; Foldes, G. It Takes Two: Endothelial-Perivascular Cell Cross-Talk in Vascular Development and Disease. Front. Cardiovasc. Med. 2018, 5, 154. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; D’Souza, S.S.; Moskvin, O.V.; Toh, H.; Wang, B.; Zhang, J.; Swanson, S.; Guo, L.-W.; Thomson, J.A.; Slukvin, I.I. Specification and Diversification of Pericytes and Smooth Muscle Cells from Mesenchymoangioblasts. Cell Rep. 2017, 19, 1902–1916. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Herrmann, M.; Bara, J.; Sprecher, C.; Menzel, U.; Jalowiec, J.; Osinga, R.; Scherberich, A.; Alini, M.; Verrier, S. Pericyte plasticity—Comparative investigation of the angiogenic and multilineage potential of pericytes from different human tissues. Eur. Cells Mater. 2016, 31, 236–249. [Google Scholar] [CrossRef] [PubMed]
- Guimarães-Camboa, N.; Cattaneo, P.; Sun, Y.; Moore-Morris, T.; Gu, Y.; Dalton, N.D.; Rockenstein, E.; Masliah, E.; Peterson, K.L.; Stallcup, W.B.; et al. Pericytes of Multiple Organs Do Not Behave as Mesenchymal Stem Cells In Vivo. Cell Stem Cell 2017, 20, 345–359. [Google Scholar] [CrossRef] [Green Version]
- Fanelli, G.N.; Naccarato, A.G.; Scatena, C. Recent Advances in Cancer Plasticity: Cellular Mechanisms, Surveillance Strategies, and Therapeutic Optimization. Front. Oncol. 2020, 10, 569. [Google Scholar] [CrossRef] [Green Version]
- Hass, R.; Von Der Ohe, J.; Ungefroren, H. The Intimate Relationship Among EMT, MET and TME: A T(ransdifferentiation) E(nhancing) M(ix) to Be Exploited for Therapeutic Purposes. Cancers 2020, 12, 3674. [Google Scholar] [CrossRef]
- Ghajar, C.M.; Peinado, H.; Mori, H.; Matei, I.R.; Evason, K.J.; Brazier, H.; Almeida, D.; Koller, A.; Hajjar, K.A.; Stainier, D.Y.; et al. The perivascular niche regulates breast tumour dormancy. Nat. Cell Biol. 2013, 15, 807–817. [Google Scholar] [CrossRef]
- Reya, T.; Morrison, S.J.; Clarke, M.F.; Weissman, I.L. Stem cells, cancer, and cancer stem cells. Nature 2001, 414, 105–111. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lawson, D.A.; Kessenbrock, K.; Davis, R.T.; Pervolarakis, N.; Werb, Z. Tumour heterogeneity and metastasis at sin-gle-cell resolution. Nat. Cell Biol. 2018, 20, 1349–1360. [Google Scholar] [CrossRef] [PubMed]
- Liu, T.J.; Sun, B.C.; Zhao, X.L.; Zhao, X.M.; Sun, T.; Gu, Q.; Yao, Z.; Dong, X.Y.; Zhao, N.; Liu, N. CD133+ cells with cancer stem cell characteristics associates with vasculogenic mimicry in triple-negative breast cancer. Oncogene 2013, 32, 544–553. [Google Scholar] [CrossRef] [Green Version]
- Roma-Rodrigues, C.; Mendes, R.; Baptista, P.V.; Fernandes, A.R. Targeting Tumor Microenvironment for Cancer Therapy. Int. J. Mol. Sci. 2019, 20, 840. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Siemann, D.W. The unique characteristics of tumor vasculature and preclinical evidence for its selective disruption by Tumor-Vascular Disrupting Agents. Cancer Treat. Rev. 2011, 37, 63–74. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carmeliet, P.; Jain, R.K. Angiogenesis in cancer and other diseases. Nature 2000, 407, 249–257. [Google Scholar] [CrossRef] [PubMed]
- Saman, H.; Raza, S.S.; Uddin, S.; Rasul, K. Inducing Angiogenesis, a Key Step in Cancer Vascularization, and Treatment Approaches. Cancers 2020, 12, 1172. [Google Scholar] [CrossRef] [PubMed]
- Motz, G.T.; Coukos, G. The parallel lives of angiogenesis and immunosuppression: Cancer and other tales. Nat. Rev. Immunol. 2011, 11, 702–711. [Google Scholar] [CrossRef]
- Matsumoto, K.; Rambow, F.; Stanchi, F.; Mathivet, T.; Qian, J.; Giese, W.; He, L.; Lambrechts, D.; Zhou, B.; Betsholtz, C.; et al. Emerging single cell endothelial heterogeneity supports sprouting tumour angiogenesis and growth. bioRxiv 2021. [Google Scholar] [CrossRef]
- Melero-Martin, J.M.; Dudley, A.C. Concise Review: Vascular Stem Cells and Tumor Angiogenesis. Stem Cells 2011, 29, 163–168. [Google Scholar] [CrossRef]
- Chen, W.Z.; Jiang, J.X.; Yu, X.Y.; Xia, W.J.; Yu, P.X.; Wang, K.; Zhao, Z.Y.; Chen, Z.G. Endothelial cells in colorectal cancer. World J. Gastrointest. Oncol. 2019, 11, 946–956. [Google Scholar] [CrossRef] [PubMed]
- Moschetta, M.; Mishima, Y.; Sahin, I.; Manier, S.; Glavey, S.; Vacca, A.; Roccaro, A.M.; Ghobrial, I.M. Role of endothelial progenitor cells in cancer progression. Biochim. Biophys. Acta (BBA)-Rev. Cancer 2014, 1846, 26–39. [Google Scholar] [CrossRef] [PubMed]
- Yan, F.; Liu, X.; Ding, H.; Zhang, W. Paracrine mechanisms of endothelial progenitor cells in vascular repair. Acta Histochem. 2022, 124, 151833. [Google Scholar] [CrossRef] [PubMed]
- Armani, G.; Pozzi, E.; Pagani, A.; Porta, C.; Rizzo, M.; Cicognini, D.; Rovati, B.; Moccia, F.; Pedrazzoli, P.; Ferraris, E. The heterogeneity of cancer endothelium: The relevance of angiogenesis and endothelial progenitor cells in cancer microenvironment. Microvasc. Res. 2021, 138, 104189. [Google Scholar] [CrossRef]
- Pacini, S.; Montali, M.; Mazziotta, F.; Schifone, C.P.; Macchia, L.; Carnicelli, V.; Panvini, F.M.; Barachini, S.; Notarfranchi, L.; Previti, G.B.; et al. Mesangiogenic progenitor cells are forced toward the angiogenic fate, in multiple myeloma. Oncotarget 2019, 10, 6781–6790. [Google Scholar] [CrossRef] [Green Version]
- Bhartiya, D.; Jha, N.; Tripathi, A.; Tripathi, A. Very small embryonic-like stem cells have the potential to win the three-front war on tissue damage, cancer, and aging. Front. Cell Dev. Biol. 2023, 10, 1061022. [Google Scholar] [CrossRef]
- Singh, P.; Metkari, S.M.; Bhartiya, D. Mice Uterine Stem Cells are Affected by Neonatal Endocrine Disruption & In-itiate Uteropathies in Adult Life Independent of Circulatory Ovarian Hormones. Stem Cell Rev. Rep. 2022, 18, 1686–1701. [Google Scholar]
- Saravanan, S.; Vimalraj, S.; Pavani, K.; Nikarika, R.; Sumantran, V.N. Intussusceptive angiogenesis as a key therapeutic target for cancer therapy. Life Sci. 2020, 252, 117670. [Google Scholar] [CrossRef]
- Fathi Maroufi, N.; Taefehshokr, S.; Rashidi, M.R.; Taefehshokr, N.; Khoshakhlagh, M.; Isazadeh, A.; Mokarizadeh, N.; Baradaran, B.; Nouri, M. Vascular mimicry: Changing the therapeutic paradigms in cancer. Mol. Biol. Rep. 2020, 47, 4749–4765. [Google Scholar] [CrossRef]
- Fernandez-Cortes, M.; Delgado-Bellido, D.; Oliver, F.J. Vasculogenic Mimicry: Become an Endothelial Cell “But Not So Much”. Front. Oncol. 2019, 9, 803. [Google Scholar] [CrossRef] [Green Version]
- Klopp, A.H.; Gupta, A.; Spaeth, E.; Andreeff, M.; Marini, F., 3rd. Concise review: Dissecting a discrepancy in the liter-ature: Do mesenchymal stem cells support or suppress tumor growth? Stem Cells 2011, 29, 11–19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, Q.; Eichten, A.; A Parveen, A.; Adler, C.; Huang, Y.; Wang, W.; Ding, Y.; Adler, A.; Nevins, T.; Ni, M.; et al. Single-Cell Transcriptome Analyses Reveal Endothelial Cell Heterogeneity in Tumors and Changes following Antiangiogenic Treatment. Cancer Res 2018, 78, 2370–2382. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thijssen, V.L.; Paulis, Y.W.; Nowak-Sliwinska, P.; Deumelandt, K.L.; Hosaka, K.; Soetekouw, P.M.; Cimpean, A.M.; Raica, M.; Pauwels, P.; Oord, J.J.V.D.; et al. Targeting PDGF-mediated recruitment of pericytes blocks vascular mimicry and tumor growth. J. Pathol. 2018, 246, 447–458. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pinto, M.P.; Sotomayor, P.; Carrasco-Avino, G.; Corvalan, A.H.; Owen, G.I. Escaping Antiangiogenic Therapy: Strategies Employed by Cancer Cells. Int. J. Mol. Sci. 2016, 17, 1489. [Google Scholar] [CrossRef] [Green Version]
- Donnem, T.; Hu, J.; Ferguson, M.; Adighibe, O.; Snell, C.; Harris, A.L.; Gatter, K.C.; Pezzella, F. Vessel co-option in primary human tumors and metastases: An obstacle to effective anti-angiogenic treatment? Cancer Med. 2013, 2, 427–436. [Google Scholar] [CrossRef] [Green Version]
- Karthik, S.; Djukic, T.; Kim, J.-D.; Zuber, B.; Makanya, A.; Odriozola, A.; Hlushchuk, R.; Filipovic, N.; Jin, S.W.; Djonov, V. Synergistic interaction of sprouting and intussusceptive angiogenesis during zebrafish caudal vein plexus development. Sci. Rep. 2018, 8, 9840. [Google Scholar] [CrossRef] [Green Version]
- Javanmard, S.H.; Zarrin, B.; Zarifi, F.; Vaseghi, G. Acquired tumor resistance to antiangiogenic therapy: Mechanisms at a glance. J. Res. Med. Sci. 2017, 22, 117. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Marker | Clinical Trials (Analyzed Disease) | Clinical Issue | PMID |
|---|---|---|---|
| cKit+ | COVID-19 and bioprosthetic total artificial heart implantation, glioblastoma growth glioblastoma | Atherosclerosis, angiogenesis in cancer, vasculogenesis in cardiovascular diseases | 33205351, 24311637, 20821342 |
| CD19+ | COVID-19 and bioprosthetic total artificial heart implantation | Vasculogenesis in cardiovascular diseases | 33205351 |
| CD31+ | Metastatic Renal Cell Carcinoma, ovarian cancer, Hepatocellular Carcinoma, ovarian or peritoneal cancer, epithelial ovarian cancer, metastatic pancreatic cancer, stent implantation | Angiogenesis in cancer, angiogenic sprouting and vessel permeability, tumor microvessel density, microvascularization, metastasis, injury by implantation | 34921022, 33103699, 29059426, 26210785, 20870280, 20388201, 20130876, 25915510 |
| CD34+ | COVID-19 and bioprosthetic total artificial heart implantation, epithelial ovarian cancer, metastatic pancreatic cancer, hypoxic stress, arterial erectile dysfunction | Vasculogenesis in cardiovascular diseases, metastasis, injury by hypoxia, cavernous arterial insufficiency | 33205351, 20388201, 20130876, 27761657, 21868743 |
| CD45+ | COVID-19 and bioprosthetic total artificial heart implantation, electronic cigarettes | Vasculogenesis in cardiovascular diseases, increase of endothelial progenitor cells | 33205351, 27693003 |
| CD14+ | Angiogenic Homeostasis in Diabetes¸ hypoxic stress, electronic cigarettes | Injury in diabetes, injury by hypoxia, increase of endothelial progenitor cells | 31102457, 27761657, 27693003 |
| CD105+ | Epithelial ovarian cancer, ovarian cancer | Cancer angiogenesis, neoangiogenesis | 20388201, 19135712 |
| CD144/VE-cadherin+ | Atrial fibrillation, electronic cigarettes¸ oxidative stress and hypoxia by extreme altitude, stent implantation, arterial erectile dysfunction, ingestion of saturated fatty acids | Protrombotic injury, increase of endothelial progenitor cells, injury by hypoxia and oxidative stress, injury by implantation, cavernous arterial insufficiency, injury by saturated fatty acids | 33237804, 27693003, 26820158, 25915510, 21868743, 19846183 |
| CD146+ | Mediterranean diet with nuts (MedDiet-nuts), coronary artery disease, metastatic colorectal cancer, coronary intervention | Cardiovascular risk, stress-induced arteriogenesis, prognosis, extent of endothelial injury | 27052787, 23056467, 21825101, 20846600 |
| CD45- | Metastatic pancreatic cancer, arterial erectile dysfunction | Metastasis formation, cavernous arterial insufficiency | 20130876, 21868743 |
| CD146- | Cancer therapy | Prognosis | 22028623 |
| Tie2- | Angiogenesis in glioblastoma | Cancer angiogenesis | 20821342 |
| vWF+ | Hematological malignancy | Thrombosis and bleeding complications. | 31390488 |
| VEGFR2+ | Angiogenesis in glioblastoma, tumor shrinkage clear-cell metastatic renal cell carcinoma, melanoma, acute myeloid leukemia, advanced cancer | Cancer angiogenesis | 20821342, 20215520, 19284623, 19188183, 17169805, 15867205 |
| Flk-1 | Metastatic breast cancer, seasonal allergic rhinitis | Tumor progression, angiogenesis in patients with asthma | 17651148, 22585426 |
| Marker | Clinical Trials (Analyzed Disease) | Clinical Issue | PMID |
|---|---|---|---|
| Gli1+ | Metastatic pancreatic cancer, advanced solid tumors, head and neck squamous cell carcinoma | Metastasis formation, prognosis | 31787526, 28317088, 21357786 |
| PDGFRβ | Thymic epithelial tumors, meningiomas, advanced renal-cell carcinoma, macular degeneration, malignant meningiomas, metastatic breast cancer, metastatic salivary gland carcinomas | Tumor regression, prognosis, survival, better outcomes, pharmacokinetics, antigiogenic effects | 34706060, 31220093, 30374686, 31791663, 31220093, 28464908, 27821319 |
| Marker | Clinical Trials (Analyzed Disease) | Clinical Issue | PMID |
|---|---|---|---|
| Sox2 | Myeloma; multiple myeloma; antimyeloma immunity, non-small-cell lung cancer | Tumor progression, tumor prevention (antimyeloma immunity), metastasis formation, prognosis | 30830874, 26827660, 23430442, 22837720 |
| Ki-67- | Node-Positive Breast Cancer | Prognosis | 31407967 |
| Nestin+ | Breast Cancer; myeloproliferative neoplasms; dermatofibrosarcoma protuberans; glioblastoma multiforme | Tumor progression, prognosis, tumor invasion, | 34615722, 30409796, 23962157, 20063522 |
| Marker | Clinical Trials | Pathological Issue | PMID |
|---|---|---|---|
| CD13+ | Diffuse large B-cell lymphoma, primary central nervous system lymphoma, B cell lymphoma, solid tumors | Vascular permeability and CNS access of anticancer drugs, blood-brain barrier (BBB) penetration, diagnostic subtyping, cancer angiogenesis | 32766857, 31118164, 30066366, 19900802 |
| α-SMA | Colorectal cancer, nasopharyngeal carcinoma, chronic allograft nephropathy, usual interstitial pneumonia and nonspecific interstitial pneumonia | Metastasis and immune cell activity, neoangiogenesis, allograft injury (transplantation), fibrotic diseases | 27248825, 24877105, 16236802, 15955241 |
| NG-2 | Neurologic and neurometabolic diseases | Replacement in neurologic diseases | 24558163 |
| RGS 5 | Metastatic colorectal cancer | Tumor maturation | 25069475 |
| Desmin | Calcifying fibrous tumor, esophageal cancer, childhood sarcomas | Diagnosis, tumor progression and depth of invasion, diagnosis | 31355265, 23801316, 1710539 |
| DLK-1 | Central precocious puberty, lung cancer | Timely diagnosis and treatment, prognostic factor | 36577869, 33508526 |
| Endosialin | Metastatic soft-tissue sarcomas, solid tumors, treatment-refractory solid tumors | Tumor angiogenesis, tumor shrinkage, | 31034598, 30623276, 25398449 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Barachini, S.; Ghelardoni, S.; Madonna, R. Vascular Progenitor Cells: From Cancer to Tissue Repair. J. Clin. Med. 2023, 12, 2399. https://doi.org/10.3390/jcm12062399
Barachini S, Ghelardoni S, Madonna R. Vascular Progenitor Cells: From Cancer to Tissue Repair. Journal of Clinical Medicine. 2023; 12(6):2399. https://doi.org/10.3390/jcm12062399
Chicago/Turabian StyleBarachini, Serena, Sandra Ghelardoni, and Rosalinda Madonna. 2023. "Vascular Progenitor Cells: From Cancer to Tissue Repair" Journal of Clinical Medicine 12, no. 6: 2399. https://doi.org/10.3390/jcm12062399
APA StyleBarachini, S., Ghelardoni, S., & Madonna, R. (2023). Vascular Progenitor Cells: From Cancer to Tissue Repair. Journal of Clinical Medicine, 12(6), 2399. https://doi.org/10.3390/jcm12062399