Sub-Inhibitory Clindamycin and Azithromycin reduce S. aureus Exoprotein Induced Toxicity, Inflammation, Barrier Disruption and Invasion




Abstract
:1. Introduction
2. Experimental Section
2.1. Cells, Bacteria and Antibiotics
2.2. Bacterial Cell Culture Supernatant Collection and Protein Concentration
2.3. Antibiotic Sensitivity Test
2.4. Primary HNECs Culture and Treatment
2.5. Air Liquid Interface (ALI) Culture and Treatment
2.6. Cytotoxity Assay
2.7. IL6 and IL8 ELISA Assay
2.8. SDS Page
2.9. Biofilm Culture, Treatment and Minimum Biofilm Eradication Concentration (MBEC) Assay
2.10. S. aureus Biofilm Protein Collection and Protein Concentration
2.11. Transepithelial Electrical Resistance (TEER)
2.12. Permeability Assay
2.13. Immunofluorescence Staining
2.14. Intracellular S. aureus Infection
2.15. Statistical Analysis
3. Results
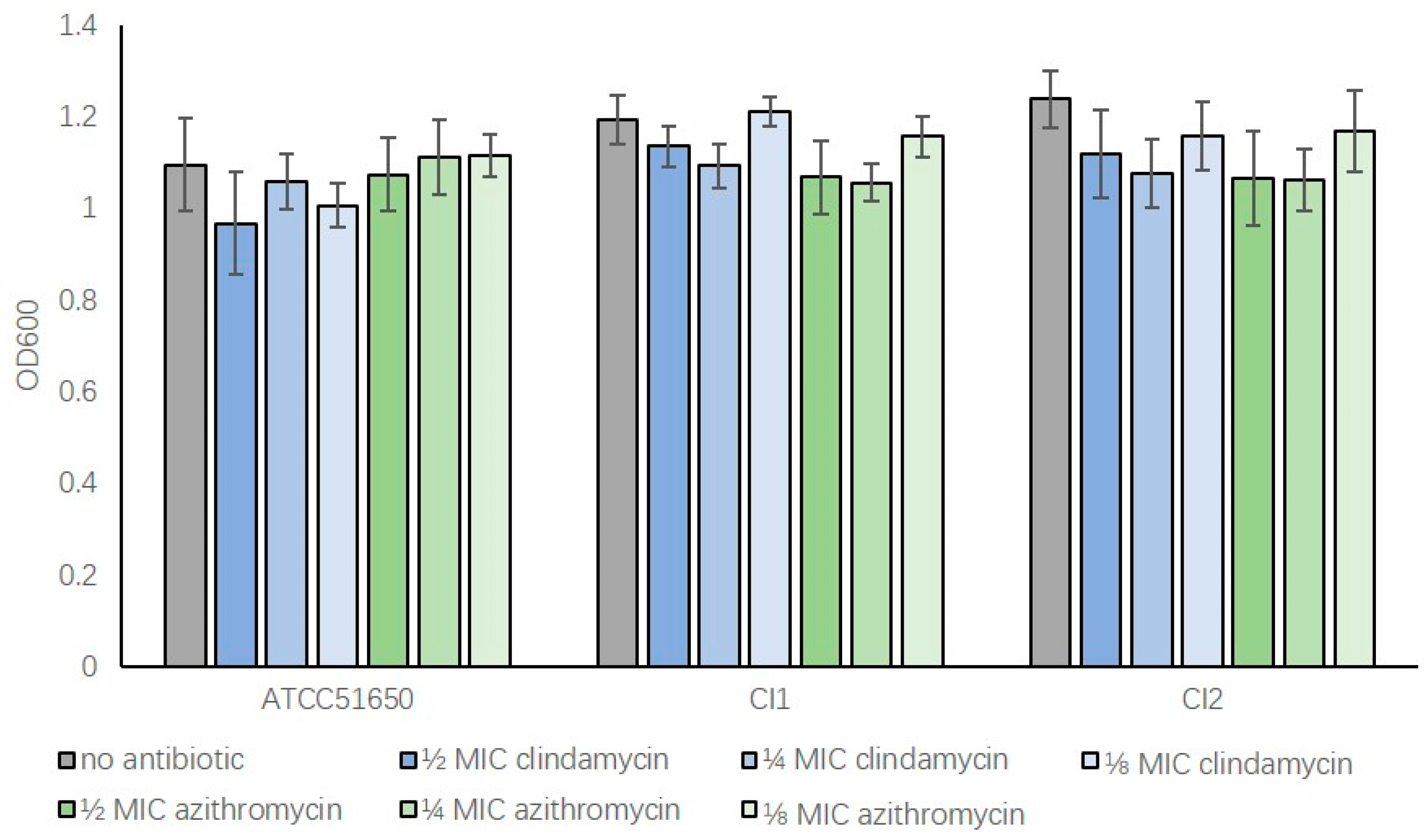
3.1. S. aureus MIC and Bacterial Growth with Subinhibitory Clindamycin and Azithromycin
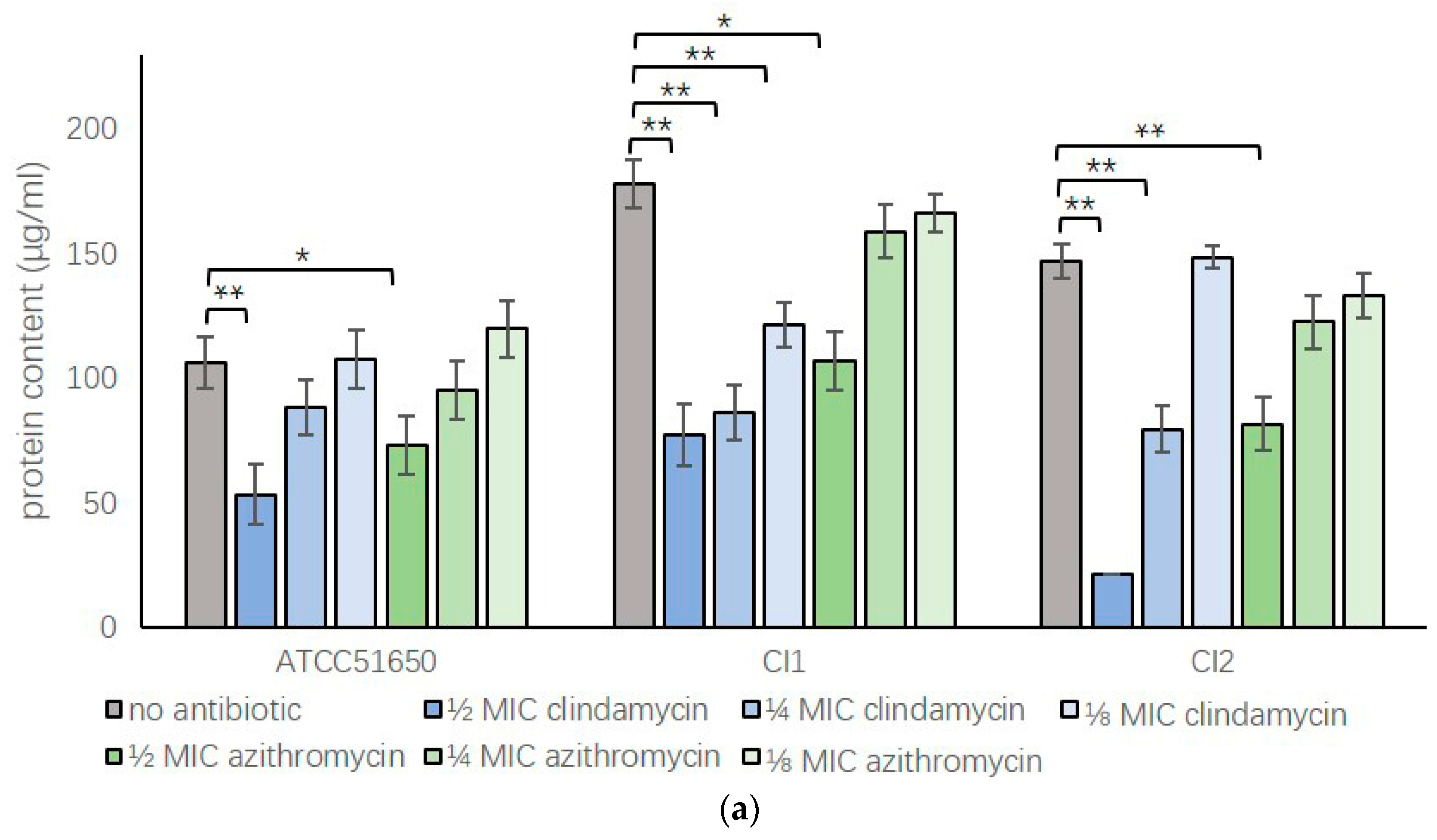
3.2. Sub-Inhibitory Clindamycin and Azithromycin Reduced S. aureus Exoprotein Secretion
3.3. Sub-Inhibitory Clindamycin Reduced S. aureus Biofilm Protein Content
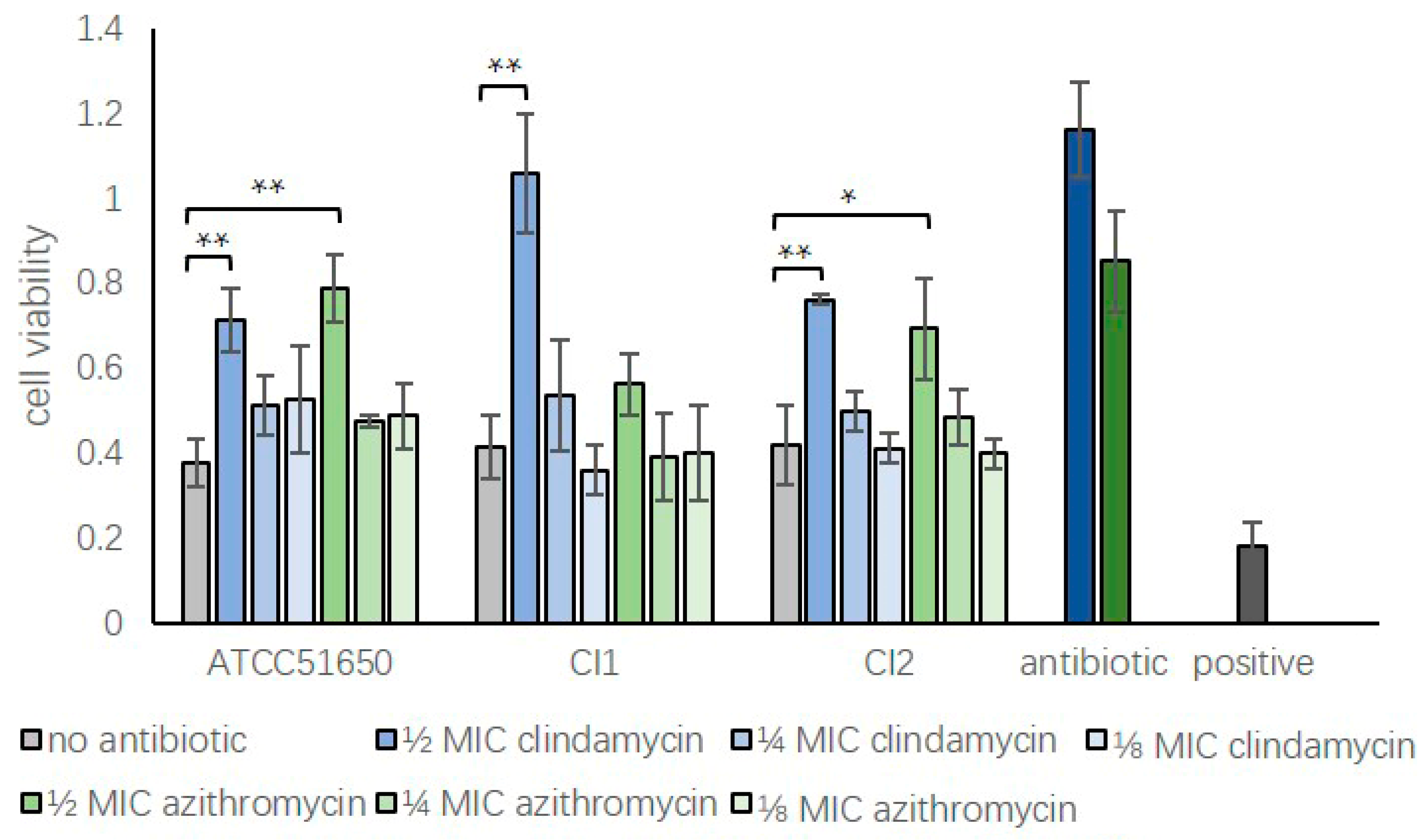
3.4. Sub-Inhibitory Clindamycin Reduced Bacterial Exoprotein-Induced Cytotoxicity of HNECs
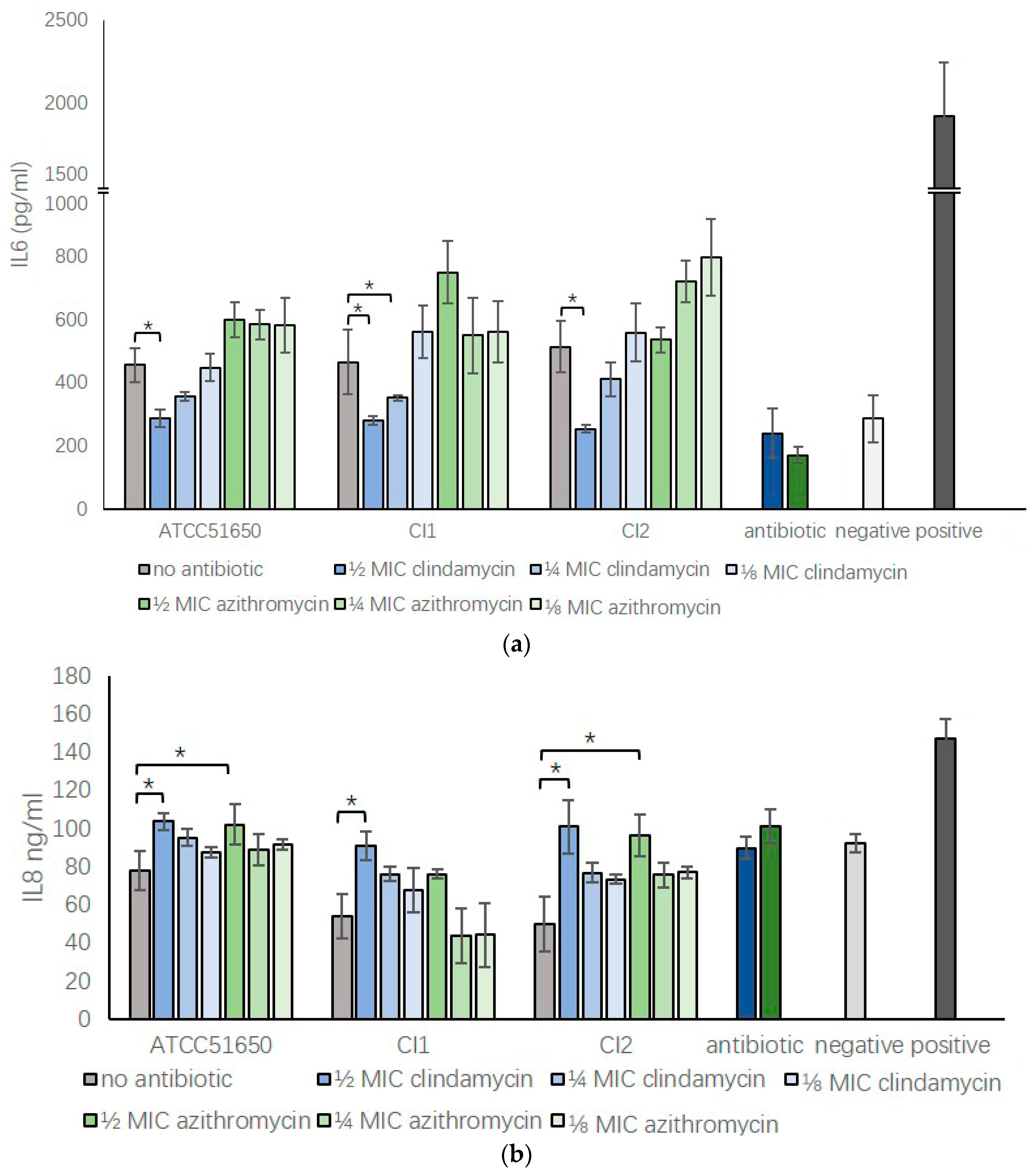
3.5. Sub-Inhibitory Clindamycin Reversed S. aureus Exoprotein Induced IL6 And IL8 Secretion by HNECs
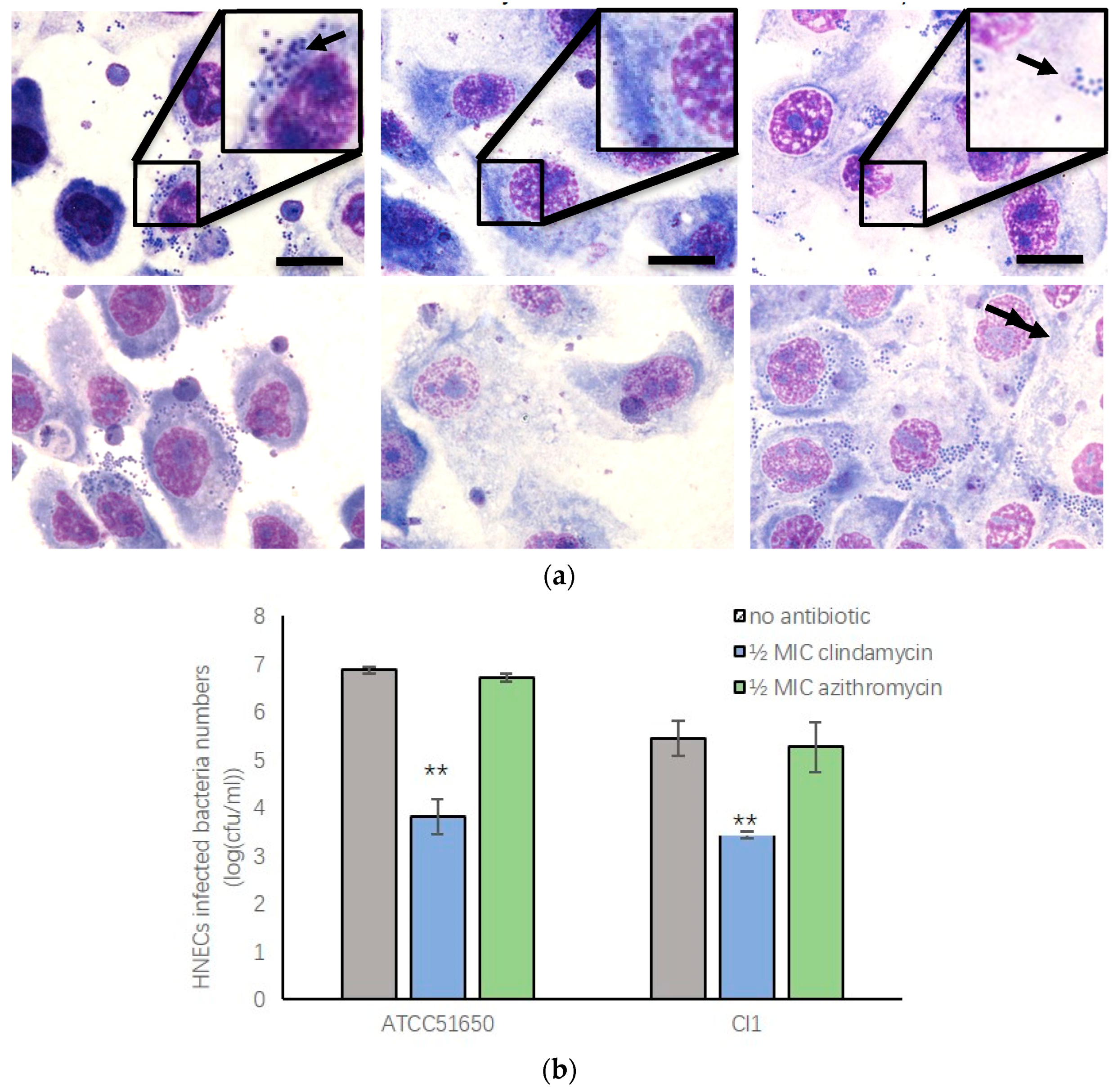
3.6. Sub-Inhibitory Clindamycin Treatment of S. aureus Reduced Intracellular Infection Rate of HNECs
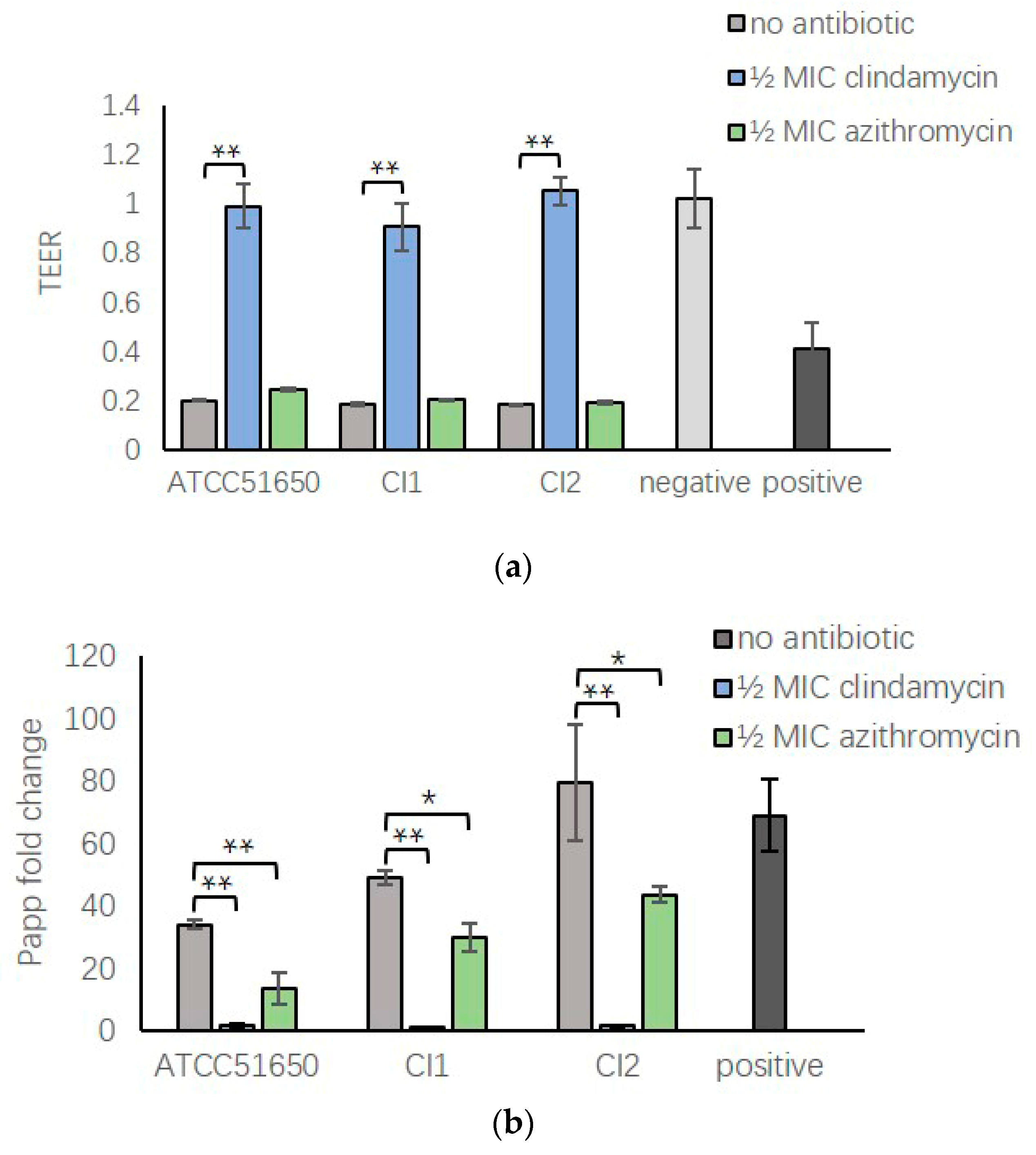
3.7. ½ MIC Clindamycin Reversed Bacterial Exoprotein Induced Mucosal Barrier Disruption of HNEC-ALI
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Smith, S.S.; Evans, C.T.; Tan, B.K.; Chandra, R.K.; Smith, S.B.; Kern, R.C. National burden of antibiotic use for adult rhinosinusitis. J. Allergy Clin. Immunol. 2013, 132, 1230–1232. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stevens, W.W.; Lee, R.J.; Schleimer, R.P.; Cohen, N.A. Chronic rhinosinusitis pathogenesis. J. Allergy Clin. Immunol. 2015, 136, 1442–1453. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abreu, N.A.; Naqalinqam, N.A.; Song, Y.; Roediqer, F.C.; Pletcher, S.D.; Goldberq, A.N.; Lynch, S.V. Sinus microbiome diversity depletion and Corynebacterium tuberculostearicum enrichment mediates rhinosinusitis. Sci. Transl. Med. 2012, 4, 151ra124. [Google Scholar] [CrossRef] [PubMed]
- Psaltis, A.J.; Wormald, P.J. Therapy of Sinonasal Microbiome in CRS: A Critical Approach. Curr. Allergy Asthma Rep. 2017, 17, 59. [Google Scholar] [CrossRef] [PubMed]
- Brook, I. Microbiology of chronic rhinosinusitis. Eur. J. Clin. Microbiol. Infect. Dis. 2016, 35, 1059–1068. [Google Scholar] [CrossRef] [PubMed]
- Cleland, E.J.; Bassiouni, A.; Vreqde, S.; Wormald, P.J. The bacterial microbiome in chronic rhinosinusitis: Richness, diversity, postoperative changes, and patient outcomes. Am. J. Rhinol. Allergy 2016, 30, 37–43. [Google Scholar] [CrossRef]
- Foreman, A.; Jervis-Bardy, J.; Wormald, P.J. Do biofilms contribute to the initiation and recalcitrance of chronic rhinosinusitis? Laryngoscope 2011, 121, 1085–1091. [Google Scholar] [CrossRef] [PubMed]
- Ou, J.; Drilling, A.; Sinqhal, D.; Tan, N.C.; Wallis-Hill, D.; Vreuqde, S.; Psaltis, A.J.; Wormald, P.J. Association of intracellular Staphylococcus aureus with prognosis in chronic rhinosinusitis. Int. Forum Allergy Rhinol. 2016, 6, 792–799. [Google Scholar] [CrossRef] [PubMed]
- Tan, N.C.; Cooksiey, C.M.; Roscioli, E.; Douqlas, R.; Wormald, P.J.; Vreuqde, S. Small-colony variants and phenotype switching of intracellular Staphylococcus aureus in chronic rhinosinusitis. Allergy 2014, 69, 1364–1371. [Google Scholar] [CrossRef]
- Guven, M.; Karabay, O.; Akidil, O.; Yilmaz, M.S.; Yildirim, M. Detection of staphylococcal exotoxins in antrochoanal polyps and chronic rhinosinusitis with nasal polyps. Otolaryngol. Head Neck Surg. 2013, 148, 302–307. [Google Scholar] [CrossRef]
- Seiberling, K.A.; Conley, D.B.; Tripathi, A.; Grammer, L.C.; Shuh, H.; Haines, G.K.; Schleimer, R.; Kern, R.C. Superantigens and chronic rhinosinusitis: detection of staphylococcal exotoxins in nasal polyps. Laryngoscope 2005, 115, 1580–1585. [Google Scholar] [CrossRef] [PubMed]
- Patou, J.; Patou, J.; Gevaert, P.; Van Zele, T.; Holtappels, G.; van Cauwenberqe, P.; Bachert, C. Staphylococcus aureus enterotoxin B, protein A, and lipoteichoic acid stimulations in nasal polyps. J. Allergy Clin. Immunol. 2008, 121, 110–115. [Google Scholar] [CrossRef] [PubMed]
- Van Zele, T.; Gevaert, P.; Holtappels, G.; van Cauwenberqe, P.; Bachert, C. Local immunoglobulin production in nasal polyposis is modulated by superantigens. Clin. Exp. Allergy 2007, 37, 1840–1847. [Google Scholar] [CrossRef] [PubMed]
- Tomassen, P.; Tomassen, P.; Vandeplas, G.; Van Zele, T.; Cardell, L.O.; Arebro, J.; Olze, H.; Förster-Ruhrmann, U.; Kowalski, M.L.; Olszewska-Ziąber, A.; et al. Holtappels G Inflammatory endotypes of chronic rhinosinusitis based on cluster analysis of biomarkers. J. Allergy Clin. Immunol. 2016, 137, 1449–1456 e4. [Google Scholar] [PubMed]
- Fokkens, W.J.; Fokkens, W.J.; Lund, V.J.; Mullol, J.; Bachert, C.; Alobid, I.; Baroody, F.; Cohen, N.; Cervin, A.; Douglas, R.; et al. European Position Paper on Rhinosinusitis and Nasal Polyps 2012. Rhinol. Suppl. 2012, 23, 1–298. [Google Scholar] [PubMed]
- Kanoh, S.; Rubin, B.K. Rubin, Mechanisms of action and clinical application of macrolides as immunomodulatory medications. Clin. Microbiol. Rev. 2010, 23, 590–615. [Google Scholar] [CrossRef]
- Cervin, A.; Wallwork, B. Macrolide therapy of chronic rhinosinusitis. Rhinology 2007, 45, 259–267. [Google Scholar]
- Harvey, R.J.; Wallwork, B.D.; Lund, V.J. Anti-inflammatory effects of macrolides: applications in chronic rhinosinusitis. Immunol. Allergy Clin. N. Am. 2009, 29, 689–703. [Google Scholar] [CrossRef]
- Russell, N.E.; Pachorek, R.E. Clindamycin in the treatment of streptococcal and staphylococcal toxic shock syndromes. Ann. Pharm. 2000, 34, 936–939. [Google Scholar] [CrossRef]
- Herbert, S.; Barry, P.; Novick, R.P. Subinhibitory clindamycin differentially inhibits transcription of exoprotein genes in Staphylococcus aureus. Infect. Immun. 2001, 69, 2996–3003. [Google Scholar] [CrossRef]
- Wiegand, I.; Hilpert, K.; Hancock, R.E. Agar and broth dilution methods to determine the minimal inhibitory concentration (MIC) of antimicrobial substances. Nat. Protoc. 2008, 3, 163–175. [Google Scholar] [CrossRef] [PubMed]
- Ramezanpour, M.; Bolt, H.; Psaltis, A.; Wormald, P.J.; Vreugde, S. Inducing a mucosal barrier–sparing inflammatory response in laboratory-grown primary human nasal epithelial cells. Curr. Protocols Toxicol. 2019, 80, e69. [Google Scholar] [CrossRef] [PubMed]
- Ramezanpour, M.; Bolt, H.; Pasltis, A.J.; Wormald, P.-J.; Vreugde, S. Primary human nasal epithelial cells: a source of poly (I:C) LMW-induced IL-6 production. Sci. Rep. 2018, 8, 11325. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramezanpour, M.; Murphy, J.; Smith, J.L.P.; Vreugde, S.; Psaltis, A.J. In vitro safety evaluation of human nasal epithelial cell monolayers exposed to carrageenan sinus wash. Int. Forum Allergy Rhinol. 2017, 7, 1170–1177. [Google Scholar] [CrossRef] [PubMed]
- Richter, K.; Ramezanpour, M.; Thomas, N.; Prestidge, C.A.; Wormald, P.J.; Vreugde, S. Mind “De GaPP”: in vitro efficacy of deferiprone and gallium-protoporphyrin against Staphylococcus aureus biofilms. Int. Forum Allergy Rhinol. 2016, 6, 737–743. [Google Scholar] [CrossRef] [PubMed]
- Ou, J.; Bassiouni, A.; Drilling, A.; Psaltis, A.J.; Vreugde, S.; Wormald, P.J. The persistence of intracellular Staphylococcus aureus in the sinuses: A longitudinal study. Rhinology 2017, 55, 305–311. [Google Scholar] [CrossRef] [PubMed]
- Malik, Z.; Roscioli, E.; Murphy, J.; Ou, J.; Bassiouni, A.; Wormald, P.J.; Vreugde, S. Staphylococcus aureus impairs the airway epithelial barrier in vitro. Int. Forum Allergy Rhinol. 2015, 5, 551–556. [Google Scholar] [CrossRef]
- Murphy, J.; Ramezanpour, M.; Drilling, A.; Roscioli, E.; Psaltis, A.J.; Wormald, P.J.; Vreugde, S. In vitro characteristics of an airway barrier-disrupting factor secreted by Staphylococcus aureus. Int. Forum Allergy Rhinol. 2018, 80, e69. [Google Scholar] [CrossRef]
- Murphy, J.; Ramezanpour, M.; Stach, N.; Dubin, G.; Psaltis, A.J.; Wormald, P.J.; Vreugde, S. Staphylococcus aureus V8 protease disrupts the integrity of the airway epithelial barrier and impairs IL-6 production in vitro. Laryngoscope 2018, 128, E8–E15. [Google Scholar] [CrossRef]
- Parnham, M.J.; Erakovic Haber, V.; Giamarellos-Bourboulis, E.J.; Perletti, G.; Verleden, G.M.; Vos, R. Azithromycin: mechanisms of action and their relevance for clinical applications. Pharm. Ther. 2014, 143, 225–245. [Google Scholar] [CrossRef]
- Feazel, L.M.; Robertson, C.E.; Ramakrishnan, V.R.; Frank, D.N. Microbiome complexity and Staphylococcus aureus in chronic rhinosinusitis. Laryngoscope 2012, 122, 467–472. [Google Scholar] [CrossRef] [PubMed]
- Anderson, M.; Stokken, J.; Sanford, T.; Aurora, R.; Sindwani, R.A. systematic review of the sinonasal microbiome in chronic rhinosinusitis. Am. J. Rhinol. Allergy 2016, 30, 161–166. [Google Scholar] [CrossRef] [PubMed]
- Cleland, E.J.; Bassiouni, A.; Wormald, P.J. The bacteriology of chronic rhinosinusitis and the pre-eminence of Staphylococcus aureus in revision patients. Int. Forum Allergy Rhinol. 2013, 3, 642–646. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.C.; Anderson, M.J.; Kohler, P.L.; Strandberg, K.L.; Olson, M.E.; Horswill, A.R.; Schlievert, P.M.; Peterson, M.L. Proinflammatory exoprotein characterization of toxic shock syndrome Staphylococcus aureus. Biochemistry 2011, 50, 7157–7167. [Google Scholar] [CrossRef] [PubMed]
- Bhakdi, S.; Tranum-Jensen, J. Alpha-toxin of Staphylococcus aureus. Microbiol. Rev. 1991, 55, 733–751. [Google Scholar] [PubMed]
- Cantero, D.; Cooksley, C.; Bassiouni, A.; Tran, H.B.; Roscioli, E.; Wormald, P.J.; Vreugde, S. Staphylococcus aureus biofilms induce apoptosis and expression of interferon-gamma, interleukin-10, and interleukin-17A on human sinonasal explants. Am. J. Rhinol. Allergy 2015, 29, 23–28. [Google Scholar] [CrossRef] [PubMed]
- Tajima, A.; Iwase, T.; Shinji, H.; Seki, K.; Mizunoe, Y. Inhibition of endothelial interleukin-8 production and neutrophil transmigration by Staphylococcus aureus beta-hemolysin. Infect. Immun. 2009, 77, 327–334. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Antibiotic | ATCC51650 | CI1 | CI2 |
|---|---|---|---|
| Clindamycin (μg/mL) | 0.2 | 0.2 | 0.2 |
| Azithromycin (μg/mL) | 2 | 2 | 2 |
| Antibiotic | ATCC51650 | CI1 | CI2 |
|---|---|---|---|
| No antibiotic | 41.31 ± 2.36 | 37.91 ± 2.48 | 10.74 ± 2.60 |
| ½ MIC clindamycin | 12.22 ± 0.58 * | 9.18 ± 0.53 * | 1.51 ± 1.56 * |
| ¼ MIC clindamycin | 18.31 ± 0.90* | 11.79 ± 1.08 * | 3.96 ± 1.10 * |
| ⅛ MIC clindamycin | 21.45 ± 1.82 * | 16.99 ± 1.23 * | 8.51 ± 1.85 |
| ½ MIC azithromycin | 18.48 ± 0.70 * | 25.06 ± 1.89 * | 3.94 ± 2.28 * |
| ¼ MIC azithromycin | 23.82 ± 2.27 * | 31.03 ± 3.68 * | 9.46 ± 1.94 |
| ⅛ MIC azithromycin | 28.66 ± 0.65 * | 32.97 ± 2.14 | 10.46 ± 1.99 |
| Antibiotic | ATCC51650 | CI1 | CI2 |
|---|---|---|---|
| No antibiotic | 11.54 ± 1.85 | 38.85 ± 1.90 | 15.60 ± 1.04 |
| 1 MIC clindamycin | 3.80 ± 0.43 * | 1.57 ± 0.12 * | 1.79 ± 0.49 * |
| ½ MIC clindamycin | 10.27 ± 1.09 | 7.51 ± 0.68 * | 1.57 ± 0.88 * |
| 1 MIC azithromycin | 7.20 ± 0.49 * | 6.35 ± 0.68 * | 1.86 ± 0.58 * |
| ½ MIC azithromycin | 10.15 ± 2.09 | 7.43 ± 0.64 * | 7.22 ± 2.07 * |
| Antibiotic | ATCC51650 | CI1 | CI2 |
|---|---|---|---|
| No antibiotic | 49 ± 9.8% | 26.9 ± 3.3% | 4 ± 0.6% |
| ½ MIC Clindamycin | 6.3 ± 0.76% ** | 5.6 ± 3.8% ** | 2.9 ± 2.6% |
| ½ MIC Azithromycin | 42.8 ± 10.7% | 27.4 ± 19.7% | 3.6 ± 2.3% |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hu, H.; Ramezanpour, M.; Hayes, A.J.; Liu, S.; Psaltis, A.J.; Wormald, P.-J.; Vreugde, S. Sub-Inhibitory Clindamycin and Azithromycin reduce S. aureus Exoprotein Induced Toxicity, Inflammation, Barrier Disruption and Invasion. J. Clin. Med. 2019, 8, 1617. https://doi.org/10.3390/jcm8101617
Hu H, Ramezanpour M, Hayes AJ, Liu S, Psaltis AJ, Wormald P-J, Vreugde S. Sub-Inhibitory Clindamycin and Azithromycin reduce S. aureus Exoprotein Induced Toxicity, Inflammation, Barrier Disruption and Invasion. Journal of Clinical Medicine. 2019; 8(10):1617. https://doi.org/10.3390/jcm8101617
Chicago/Turabian StyleHu, Hua, Mahnaz Ramezanpour, Andrew J Hayes, Sha Liu, Alkis J Psaltis, Peter-John Wormald, and Sarah Vreugde. 2019. "Sub-Inhibitory Clindamycin and Azithromycin reduce S. aureus Exoprotein Induced Toxicity, Inflammation, Barrier Disruption and Invasion" Journal of Clinical Medicine 8, no. 10: 1617. https://doi.org/10.3390/jcm8101617
APA StyleHu, H., Ramezanpour, M., Hayes, A. J., Liu, S., Psaltis, A. J., Wormald, P. -J., & Vreugde, S. (2019). Sub-Inhibitory Clindamycin and Azithromycin reduce S. aureus Exoprotein Induced Toxicity, Inflammation, Barrier Disruption and Invasion. Journal of Clinical Medicine, 8(10), 1617. https://doi.org/10.3390/jcm8101617