Insights into the Regulation of Tumor Angiogenesis by Micro-RNAs

 ,
, 
 ,
,  ,
,  ,
,
Abstract
:1. Introduction
2. Tumor Angiogenesis
3. MicroRNAs
4. Tumor Angiogenesis and miRNAs
5. Pro-Angiogenic miRNAs
5.1. miR-155
5.2. miR-566
5.3. miR-210
5.4. miR-21
5.5. miR-182
5.6. miR-296
5.7. miR-17-92 Cluster
5.8. Lethal (Let)-7b and -7f
5.9. miR-378
5.10. miR-221 and miR-222
6. Anti-Angiogenic miRNAs
6.1. miR-221 and miR-222
6.2. miR-126
6.3. Let-7a
6.4. miR-328
6.5. miR-135a
6.6. miR-29b
6.7. miR-206
6.8. miR-140-5p
6.9. miR-497
6.10. miR-377
6.11. miR-218
6.12. miR-134
7. Clinical Implications
7.1. miRNAs as Biomarkers of the Anti-Angiogenic Therapy Response
7.2. miRNAs as Therapeutics Against Tumor Angiogenesis
8. Conclusions and Perspectives
Author Contributions
Funding
Conflicts of Interest
References
- Carmeliet, P.; Jain, R.K. Angiogenesis in Cancer and Other Diseases. Nature 2000, 407, 249–257. [Google Scholar] [CrossRef] [PubMed]
- Viallard, C.; Larrivee, B. Tumor Angiogenesis and Vascular Normalization: Alternative Therapeutic Targets. Angiogenesis 2017, 20, 409–426. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.L.C.; Rouhi, P.; Jensen, L.D.; Zhang, D.; Ji, H.; Hauptmann, G.; Ingham, P.W.; Cao, Y. Hypoxia-Induced Pathological Angiogenesis Mediates Tumor Cell Dissemination, Invasion, and Metastasis in a Zebrafish Tumor Model. Proc. Natl. Acad. Sci. USA 2009, 106, 19485–19490. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Dabrosin, C.; Yin, X.; Fuster, M.M.; Arreola, A.; Rathmell, W.K.; Generali, D.; Nagaraju, G.P.; El Rayes, B.; Ribatti, D.; et al. Broad Targeting of Angiogenesis for Cancer Prevention and Therapy. Semin. Cancer Biol. 2015, 35, S224–S243. [Google Scholar] [CrossRef] [PubMed]
- Lathia, J.D.; Heddleston, J.M.; Venere, M.; Rich, J.N. Deadly Teamwork: Neural Cancer Stem Cells and the Tumor Microenvironment. Cell Stem Cell 2011, 8, 482–485. [Google Scholar] [CrossRef]
- Goradel, N.H.; Mohammadi, N.; Haghi Aminjan, H.; Farhood, B.; Negahdari, B.; Sahebkar, A. Regulation of Tumor Angiogenesis by microRNAs: State of the Art. J. Cell Physiol. 2019, 234, 1099–1110. [Google Scholar] [CrossRef]
- Wang, Y.; Wang, L.; Chen, C.; Chu, X. New Insights into the Regulatory Role of microRNA in Tumor Angiogenesis and Clinical Implications. Mol. Cancer 2018, 17, 22. [Google Scholar] [CrossRef]
- Lujambio, A.; Lowe, S.W. The Microcosmos of Cancer. Nature 2012, 482, 347–355. [Google Scholar] [CrossRef]
- Lewis, B.P.; Burge, C.B.; Bartel, D.P. Conserved Seed Pairing, Often Flanked by Adenosines, Indicates that Thousands of Human Genes are MicroRNA Targets. Cell 2005, 120, 15–20. [Google Scholar] [CrossRef]
- Calin, G.A.; Croce, C.M. MicroRNA Signatures in Human Cancers. Nat. Rev. Cancer 2006, 6, 857–866. [Google Scholar] [CrossRef]
- Martello, G.; Rosato, A.; Ferrari, F.; Manfrin, A.; Cordenonsi, M.; Dupont, S.; Enzo, E.; Guzzardo, V.; Rondina, M.; Spruce, T.; et al. A MicroRNA Targeting Dicer for Metastasis Control. Cell 2010, 141, 1195–1207. [Google Scholar] [CrossRef] [PubMed]
- Xie, T.; Huang, M.; Wang, Y.; Wang, L.; Chen, C.; Chu, X. MicroRNAs as Regulators, Biomarkers and Therapeutic Targets in the Drug Resistance of Colorectal Cancer. Cell. Physiol. Biochem. 2016, 40, 62–76. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Xue, Y.; Wu, X.; Le, C.; Bhutkar, A.; Bell, E.L.; Zhang, F.; Langer, R.; Sharp, P.A. Global microRNA Depletion Suppresses Tumor Angiogenesis. Genes Dev. 2014, 28, 1054–1067. [Google Scholar] [CrossRef] [PubMed]
- Bentley, K.; Chakravartula, S. The Temporal Basis of Angiogenesis. Philos. Trans. R. Soc. B Boil. Sci. 2017, 372, 20150522. [Google Scholar] [CrossRef] [PubMed]
- Betz, C.; Lenard, A.; Belting, H.G.; Affolter, M. Cell Behaviors and Dynamics During Angiogenesis. Development 2016, 143, 2249–2260. [Google Scholar] [CrossRef]
- Li, J.; Sun, X.; Wang, Z.; Chen, L.; Li, D.; Zhou, J.; Liu, M. Regulation of Vascular Endothelial Cell Polarization and Migration by Hsp70/Hsp90-Organizing Protein. PLoS ONE 2012, 7, e36389. [Google Scholar] [CrossRef] [PubMed]
- Holmes, D.I.R.; Zachary, I. The Vascular Endothelial Growth Factor (VEGF) Family: Angiogenic Factors in Health and Disease. Genome Boil. 2005, 6, 209. [Google Scholar] [CrossRef]
- Kofler, N.M.; Shawber, C.J.; Kangsamaksin, T.; Reed, H.O.; Galatioto, J.; Kitajewski, J. Notch Signaling in Developmental and Tumor Angiogenesis. Genes Cancer 2011, 2, 1106–1116. [Google Scholar] [CrossRef] [PubMed]
- Saltarella, I.; Frassanito, M.A.; Lamanuzzi, A.; Brevi, A.; Leone, P.; Desantis, V.; Di Marzo, L.; Bellone, M.; Derudas, D.; Ribatti, D.; et al. Homotypic and Heterotypic Activation of the Notch Pathway in Multiple Myeloma-Enhanced Angiogenesis: A Novel Therapeutic Target? Neoplasia 2019, 21, 93–105. [Google Scholar] [CrossRef]
- Ferrara, N. Vascular Endothelial Growth Factor: Basic Science and Clinical Progress. Endocr. Rev. 2004, 25, 581–611. [Google Scholar] [CrossRef]
- Good, D.J.; Polverini, P.J.; Rastinejad, F.; Le Beau, M.M.; Lemons, R.S.; Frazier, W.A.; Bouck, N.P. A Tumor Suppressor-Dependent Inhibitor of Angiogenesis is Immunologically and Functionally Indistinguishable from a Fragment of Thrombospondin. Proc. Natl. Acad. Sci. USA 1990, 87, 6624–6628. [Google Scholar] [CrossRef] [PubMed]
- O’Reilly, M.S.; Boehm, T.; Shing, Y.; Fukai, N.; Vasios, G.; Lane, W.S.; Flynn, E.; Birkhead, J.R.; Olsen, B.R.; Folkman, J. Endostatin: An Endogenous Inhibitor of Angiogenesis and Tumor Growth. Cell 1997, 88, 277–285. [Google Scholar] [CrossRef]
- Maeshima, Y.; Sudhakar, A.; Lively, J.C.; Ueki, K.; Kharbanda, S.; Kahn, C.R.; Sonenberg, N.; Hynes, R.O.; Kalluri, R. Tumstatin, an Endothelial Cell-Specific Inhibitor of Protein Synthesis. Science 2002, 295, 140–143. [Google Scholar] [CrossRef] [PubMed]
- Magnon, C.; Galaup, A.; Mullan, B.; Rouffiac, V.; Bidart, J.M.; Griscelli, F.; Opolon, P.; Perricaudet, M. Canstatin Acts on Endothelial and Tumor Cells via Mitochondrial Damage Initiated through Interaction with v 3 and v 5 Integrins. Cancer Res. 2005, 65, 4353–4361. [Google Scholar] [CrossRef] [PubMed]
- Baeriswyl, V.; Christofori, G. The Angiogenic Switch in Carcinogenesis. Semin. Cancer Boil. 2009, 19, 329–337. [Google Scholar] [CrossRef]
- Repetto, O.; De Re, V. Coagulation and Fibrinolysis in Gastric Cancer. Ann. N. Y. Acad. Sci. 2017, 1404, 27–48. [Google Scholar] [CrossRef]
- Bergers, G.; Benjamin, L.E. Tumorigenesis and the Angiogenic Switch. Nat. Rev. Cancer 2003, 3, 401–410. [Google Scholar] [CrossRef]
- Kerbel, R.S. Tumor Angiogenesis: Past, Present and the Near Future. Carcinogenesis 2000, 21, 505–515. [Google Scholar] [CrossRef]
- Bielenberg, D.R.; Zetter, B.R. The Contribution of Angiogenesis to the Process of Metastasis. Cancer J. 2015, 21, 267–273. [Google Scholar] [CrossRef]
- Morikawa, S.; Baluk, P.; Kaidoh, T.; Haskell, A.; Jain, R.K.; McDonald, D.M. Abnormalities in Pericytes on Blood Vessels and Endothelial Sprouts in Tumors. Am. J. Pathol. 2002, 160, 985–1000. [Google Scholar] [CrossRef]
- Seaman, S.; Stevens, J.; Yang, M.Y.; Logsdon, D.; Graff Cherry, C.; Croix, B.S. Genes that Distinguish Physiological and Pathological Angiogenesis. Cancer Cell 2007, 11, 539–554. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ge, H.; Luo, H. Overview of Advances in Vasculogenic Mimicry–A Potential Target for Tumor Therapy. Cancer Manag. Res. 2018, 10, 2429–2437. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Angara, K.; Borin, T.F.; Arbab, A.S. Vascular Mimicry: A Novel Neovascularization Mechanism Driving Anti-Angiogenic Therapy (AAT) Resistance in Glioblastoma. Transl. Oncol. 2017, 10, 650–660. [Google Scholar] [CrossRef] [PubMed]
- Chang, Y.S.; Di Tomaso, E.; McDonald, D.M.; Jones, R.; Jain, R.K.; Munn, L.L. Mosaic Blood Vessels in Tumors: Frequency of Cancer Cells in Contact with Flowing Blood. Proc. Natl. Acad. Sci. USA 2000, 97, 14608–14613. [Google Scholar] [CrossRef] [Green Version]
- Yang, J.P.; Liao, Y.D.; Mai, D.M.; Xie, P.; Qiang, Y.Y.; Zheng, L.S.; Wang, M.Y.; Mei, Y.; Meng, D.F.; Xu, L.; et al. Tumor Vasculogenic Mimicry Predicts Poor Prognosis in Cancer Patients: A Meta-Analysis. Angiogenesis 2016, 19, 191–200. [Google Scholar] [CrossRef]
- Dayan, F.; Mazure, N.M.; Brahimi Horn, M.C.; Pouyssegur, J. A Dialogue between the Hypoxia-Inducible Factor and the Tumor Microenvironment. Cancer Microenviron. 2008, 1, 53–68. [Google Scholar] [CrossRef] [Green Version]
- Zimna, A.; Kurpisz, M. Hypoxia-Inducible Factor-1 in Physiological and Pathophysiological Angiogenesis: Applications and Therapies. BioMed Res. Int. 2015, 2015, 549412. [Google Scholar] [CrossRef] [Green Version]
- Bartel, D.P. MicroRNAs: Target Recognition and Regulatory Functions. Cell 2009, 136, 215–233. [Google Scholar] [CrossRef] [Green Version]
- Lee, Y.; Jeon, K.; Lee, J.; Kim, S.; Kim, V.N. MicroRNA Maturation: Stepwise Processing and Subcellular Localization. EMBO J. 2002, 21, 4663–4670. [Google Scholar] [CrossRef] [Green Version]
- Lund, E.; Guttinger, S.; Calado, A.; Dahlberg, J.E.; Kutay, U. Nuclear Export of MicroRNA Precursors. Science 2004, 303, 95–98. [Google Scholar] [CrossRef] [Green Version]
- Carthew, R.W.; Sontheimer, E.J. Origins and Mechanisms of miRNAs and siRNAs. Cell 2009, 136, 642–655. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garzon, R.; Marcucci, G.; Croce, C.M. Targeting microRNAs in Cancer: Rationale, Strategies and Challenges. Nat. Rev. Drug Discov. 2010, 9, 775–789. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, S.; Olson, E.N. AngiomiRs--Key Regulators of Angiogenesis. Curr. Opin. Genet. Dev. 2009, 19, 205–211. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kong, W.; He, L.; Richards, E.J.; Challa, S.; Xu, C.X.; Permuth Wey, J.; Lancaster, J.M.; Coppola, D.; Sellers, T.A.; Djeu, J.Y.; et al. Upregulation of miRNA-155 Promotes Tumour Angiogenesis by Targeting VHL and is Associated with Poor Prognosis and Triple-Negative Breast Cancer. Oncogene 2014, 33, 679–689. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Petrovic, N.; Kolakovic, A.; Stankovic, A.; Lukic, S.; Rami, A.; Ivkovic, M.; Mandusic, V. miR-155 Expression Level Changes Might be Associated with Initial Phases of Breast Cancer Pathogenesis and Lymph-Node Metastasis. Cancer Biomark. 2016, 16, 385–394. [Google Scholar] [CrossRef] [PubMed]
- White, N.M.; Bao, T.T.; Grigull, J.; Youssef, Y.M.; Girgis, A.; Diamandis, M.; Fatoohi, E.; Metias, M.; Honey, R.J.; Stewart, R.; et al. miRNA Profiling for Clear Cell Renal Cell Carcinoma: Biomarker Discovery and Identification of Potential Controls and Consequences of miRNA Dysregulation. J. Urol. 2011, 186, 1077–1083. [Google Scholar] [CrossRef]
- Zhou, X.; Yan, T.; Huang, C.; Xu, Z.; Wang, L.; Jiang, E.; Wang, H.; Chen, Y.; Liu, K.; Shao, Z.; et al. Melanoma Cell-Secreted Exosomal miR-155-5p Induce Proangiogenic Switch of Cancer-Associated Fibroblasts Via SOCS1/JAK2/STAT3 Signaling Pathway. J. Exp. Clin. Cancer Res. 2018, 37, 242. [Google Scholar] [CrossRef] [Green Version]
- Xiao, B.; Zhou, X.; Ye, M.; Lv, S.; Wu, M.; Liao, C.; Han, L.; Kang, C.; Zhu, X. MicroRNA566 Modulates Vascular Endothelial Growth Factor by Targeting Von HippelLandau in Human Glioblastoma in Vitro and in Vivo. Mol. Med. Rep. 2016, 13, 379–385. [Google Scholar] [CrossRef]
- Zhang, K.L.; Zhou, X.; Han, L.; Chen, L.Y.; Chen, L.C.; Shi, Z.D.; Yang, M.; Ren, Y.; Yang, J.X.; Frank, T.S.; et al. MicroRNA-566 Activates EGFR Signaling and its Inhibition Sensitizes Glioblastoma Cells to Nimotuzumab. Mol. Cancer 2014, 13, 63. [Google Scholar] [CrossRef] [Green Version]
- Camps, C.; Buffa, F.M.; Colella, S.; Moore, J.; Sotiriou, C.; Sheldon, H.; Harris, A.L.; Gleadle, J.M.; Ragoussis, J. Hsa-Mir-210 Is Induced by Hypoxia and Is an Independent Prognostic Factor in Breast Cancer. Clin. Cancer Res. 2008, 14, 1340–1348. [Google Scholar] [CrossRef] [Green Version]
- Giannakakis, A.; Sandaltzopoulos, R.; Greshock, J.; Liang, S.; Huang, J.; Hasegawa, K.; Li, C.; O’Brien Jenkins, A.; Katsaros, D.; Weber, B.L.; et al. miR-210 Links Hypoxia with Cell Cycle Regulation and is Deleted in Human Epithelial Ovarian Cancer. Cancer Biol. Ther. 2008, 7, 255–264. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lou, Y.L.; Guo, F.; Liu, F.; Gao, F.L.; Zhang, P.Q.; Niu, X.; Guo, S.C.; Yin, J.H.; Wang, Y.; Deng, Z.F. miR-210 Activates Notch Signaling Pathway in Angiogenesis Induced by Cerebral Ischemia. Mol. Cell. Biochem. 2012, 370, 45–51. [Google Scholar] [CrossRef] [PubMed]
- Fasanaro, P.; D’Alessandra, Y.; Di Stefano, V.; Melchionna, R.; Romani, S.; Pompilio, G.; Capogrossi, M.C.; Martelli, F. MicroRNA-210 Modulates Endothelial Cell Response to Hypoxia and Inhibits the Receptor Tyrosine Kinase Ligand Ephrin-A3. J. Boil. Chem. 2008, 283, 15878–15883. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, Y.; Zhang, J.; Xia, T.; Li, G.; Tian, T.; Wang, M.; Wang, R.; Zhao, L.; Yang, Y.; Lan, K.; et al. MicroRNA-210 Promotes Cancer Angiogenesis by Targeting Fibroblast Growth Factor Receptor-Like 1 in Hepatocellular Carcinoma. Oncol. Rep. 2016, 36, 2553–2562. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.Z.; Li, C.; Chen, Q.; Jing, Y.; Carpenter, R.; Jiang, Y.; Kung, H.F.; Lai, L.; Jiang, B.H. MiR-21 Induced Angiogenesis Through AKT and ERK Activation and HIF-1α Expression. PLoS ONE 2011, 6, e19139. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Siegal, T.; Charbit, H.; Paldor, I.; Zelikovitch, B.; Canello, T.; Benis, A.; Wong, M.L.; Morokoff, A.P.; Kaye, A.H.; Lavon, I. Dynamics of Circulating Hypoxia-Mediated miRNAs and Tumor Response in Patients with High-Grade Glioma Treated with Bevacizumab. J. Neurosurg. 2016, 125, 1008–1015. [Google Scholar] [CrossRef] [Green Version]
- Bao, B.; Ali, S.; Ahmad, A.; Azmi, A.S.; Li, Y.; Banerjee, S.; Kong, D.; Sethi, S.; Aboukameel, A.; Padhye, S.B.; et al. Hypoxia-Induced Aggressiveness of Pancreatic Cancer Cells Is Due to Increased Expression of VEGF, IL-6 and miR-21, Which Can Be Attenuated by CDF Treatment. PLoS ONE 2012, 7, e50165. [Google Scholar] [CrossRef] [Green Version]
- Qian, B.; Katsaros, D.; Lu, L.; Preti, M.; Durando, A.; Arisio, R.; Mu, L.; Yu, H. High miR-21 Expression in Breast Cancer Associated with Poor Disease-Free Survival in Early Stage Disease and High TGF-Beta1. Breast Cancer Res. Treat. 2009, 117, 131–140. [Google Scholar] [CrossRef]
- Meng, F.; Henson, R.; Wehbe Janek, H.; Ghoshal, K.; Jacob, S.T.; Patel, T. MicroRNA-21 Regulates Expression of the PTEN Tumor Suppressor Gene in Human Hepatocellular Cancer. Gastroenterology 2007, 133, 647–658. [Google Scholar] [CrossRef] [Green Version]
- Asangani, I.A.; Rasheed, S.A.; Nikolova, D.A.; Leupold, J.H.; Colburn, N.H.; Post, S.; Allgayer, H. MicroRNA-21 (miR-21) Post-Transcriptionally Downregulates Tumor Suppressor Pdcd4 and Stimulates Invasion, Intravasation and Metastasis in Colorectal Cancer. Oncogene 2008, 27, 2128–2136. [Google Scholar] [CrossRef] [Green Version]
- Chiang, C.H.; Chu, P.Y.; Hou, M.F.; Hung, W.C. MiR-182 Promotes Proliferation and Invasion and Elevates the HIF-1alpha-VEGF-A Axis in Breast Cancer Cells by Targeting FBXW7. Am. J. Cancer Res. 2016, 6, 1785–1798. [Google Scholar] [PubMed]
- Li, Y.; Zhang, D.; Wang, X.; Yao, X.; Ye, C.; Zhang, S.; Wang, H.; Chang, C.; Xia, H.; Wang, Y.C.; et al. Hypoxia-Inducible miR-182 Enhances HIF1α Signaling Via Targeting PHD2 and FIH1 in Prostate Cancer. Sci. Rep. 2015, 5, 12495. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wurdinger, T.; Tannous, B.A.; Saydam, O.; Skog, J.; Grau, S.; Soutschek, J.; Weissleder, R.; Breakefield, X.O.; Krichevsky, A.M. miR-296 Regulates Growth Factor Receptor Overexpression in Angiogenic Endothelial Cells. Cancer Cell 2008, 14, 382–393. [Google Scholar] [CrossRef] [Green Version]
- Chamorro Jorganes, A.; Lee, M.Y.; Araldi, E.; Landskroner Eiger, S.; Fernandez Fuertes, M.; Sahraei, M.; Del Rey, M.Q.; Van Solingen, C.; Yu, J.; Fernandez Hernando, C.; et al. VEGF-Induced Expression of miR-17–92 Cluster in Endothelial Cells Is Mediated by ERK/ELK1 Activation and Regulates Angiogenesis. Circ. Res. 2016, 118, 38–47. [Google Scholar] [CrossRef] [PubMed]
- Taguchi, A.; Yanagisawa, K.; Tanaka, M.; Cao, K.; Matsuyama, Y.; Goto, H.; Takahashi, T. Identification of Hypoxia-Inducible Factor-1 as a Novel Target for miR-17-92 MicroRNA Cluster. Cancer Res. 2008, 68, 5540–5545. [Google Scholar] [CrossRef] [Green Version]
- Otsuka, M.; Zheng, M.; Hayashi, M.; Lee, J.D.; Yoshino, O.; Lin, S.; Han, J. Impaired microRNA Processing Causes Corpus Luteum Insufficiency and Infertility in Mice. J. Clin. Investig. 2008, 118, 1944–1954. [Google Scholar] [CrossRef] [Green Version]
- Tiwari, A.; Mukherjee, B.; Dixit, M. MicroRNA Key to Angiogenesis Regulation: MiRNA Biology and Therapy. Curr. Cancer Drug Targets 2018, 18, 266–277. [Google Scholar] [CrossRef]
- Wang, Z.; Xu, L.; Hu, Y.; Huang, Y.; Zhang, Y.; Zheng, X.; Wang, S.; Wang, Y.; Yu, Y.; Zhang, M.; et al. miRNA let-7b Modulates Macrophage Polarization and Enhances Tumor-Associated Macrophages to Promote Angiogenesis and Mobility in Prostate Cancer. Sci. Rep. 2016, 6, 25602. [Google Scholar] [CrossRef] [Green Version]
- Chan, J.K.; Kiet, T.K.; Blansit, K.; Ramasubbaiah, R.; Hilton, J.F.; Kapp, D.S.; Matei, D. MiR-378 as a Biomarker for Response to Anti-Angiogenic Treatment in Ovarian Cancer. Gynecol. Oncol. 2014, 133, 568–574. [Google Scholar] [CrossRef]
- Lee, D.Y.; Deng, Z.; Wang, C.H.; Yang, B.B. MicroRNA-378 Promotes Cell Survival, Tumor Growth, and Angiogenesis by Targeting SuFu and Fus-1 Expression. Proc. Natl. Acad. Sci. USA 2007, 104, 20350–20355. [Google Scholar] [CrossRef] [Green Version]
- Chen, L.T.; Xu, S.D.; Xu, H.; Zhang, J.F.; Ning, J.F.; Wang, S.F. MicroRNA-378 is Associated with Non-Small Cell Lung Cancer Brain Metastasis by Promoting Cell Migration, Invasion and Tumor Angiogenesis. Med. Oncol. 2012, 29, 1673–1680. [Google Scholar] [CrossRef] [PubMed]
- Zaravinos, A.; Radojicic, J.; Lambrou, G.I.; Volanis, D.; Delakas, D.; Stathopoulos, E.N.; Spandidos, D.A. Expression of miRNAs Involved in Angiogenesis, Tumor Cell Proliferation, Tumor Suppressor Inhibition, Epithelial-Mesenchymal Transition and Activation of Metastasis in Bladder Cancer. J. Urol. 2012, 188, 615–623. [Google Scholar] [CrossRef]
- Xu, C.; Liu, Y.; Xiao, L.; Chen, L.; Zheng, S.; Zeng, E.; Li, D.; Li, Y. Silencing microRNA-221/222 Cluster Suppresses Glioblastoma Angiogenesis by Suppressor of Cytokine Signaling-3-Dependent JAK/STAT Pathway. J. Cell. Physiol. 2019, 234, 22272–22284. [Google Scholar] [CrossRef] [PubMed]
- Yang, F.; Wang, W.; Zhou, C.; Xi, W.; Yuan, L.; Chen, X.; Li, Y.; Yang, A.; Zhang, J.; Wang, T. MiR-221/222 Promote Human Glioma Cell Invasion and Angiogenesis by Targeting TIMP2. Tumor Boil. 2015, 36, 3763–3773. [Google Scholar] [CrossRef] [PubMed]
- Poliseno, L.; Tuccoli, A.; Mariani, L.; Evangelista, M.; Citti, L.; Woods, K.; Mercatanti, A.; Hammond, S.; Rainaldi, G. MicroRNAs Modulate the Angiogenic Properties of HUVECs. Blood 2006, 108, 3068–3071. [Google Scholar] [CrossRef] [PubMed]
- Khella, H.W.Z.; Butz, H.; Ding, Q.; Rotondo, F.; Evans, K.R.; Kupchak, P.; Dharsee, M.; Latif, A.; Pasic, M.D.; Lianidou, E.; et al. miR-221/222 Are Involved in Response to Sunitinib Treatment in Metastatic Renal Cell Carcinoma. Mol. Ther. 2015, 23, 1748–1758. [Google Scholar] [CrossRef]
- Zhu, N.; Zhang, D.; Chen, S.; Liu, X.; Lin, L.; Huang, X.; Guo, Z.; Liu, J.; Wang, Y.; Yuan, W.; et al. Endothelial Enriched microRNAs Regulate Angiotensin II-Induced Endothelial Inflammation and Migration. Atherosclerosis 2011, 215, 286–293. [Google Scholar] [CrossRef]
- Nicoli, S.; Knyphausen, C.P.; Zhu, L.J.; Lakshmanan, A.; Lawson, N.D. miR-221 is Required for Endothelial Tip Cell Behaviors During Vascular Development. Dev. Cell 2012, 22, 418–429. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.; Banda, M.; Speyer, C.L.; Smith, J.S.; Rabson, A.B.; Gorski, D.H. Regulation of the Expression and Activity of the Antiangiogenic Homeobox Gene GAX/MEOX2 by ZEB2 and MicroRNA-221. Mol. Cell. Boil. 2010, 30, 3902–3913. [Google Scholar] [CrossRef] [Green Version]
- Gorski, D.H.; Leal, A.J. Inhibition of Endothelial Cell Activation by the Homeobox Gene Gax. J. Surg. Res. 2003, 111, 91–99. [Google Scholar] [CrossRef]
- Patel, S.; Leal, A.D.; Gorski, D.H. The Homeobox GeneGaxInhibits Angiogenesis Through Inhibition of Nuclear Factor-ΚB–Dependent Endothelial Cell Gene Expression. Cancer Res. 2005, 65, 1414–1424. [Google Scholar] [CrossRef] [Green Version]
- Kontomanolis, E.; Mitrakas, A.; Giatromanolaki, A.; Kareli, D.; Panteliadou, M.; Pouliliou, S.; Koukourakis, M.I. A Pilot Study on Plasma Levels of micro-RNAs Involved in Angiogenesis and Vascular Maturation in Patients with Breast Cancer. Med Oncol. 2017, 34, 20. [Google Scholar] [CrossRef] [PubMed]
- Dentelli, P.; Rosso, A.; Orso, F.; Olgasi, C.; Taverna, D.; Brizzi, M.F. microRNA-222 Controls Neovascularization by Regulating Signal Transducer and Activator of Transcription 5A Expression. Arter. Thromb. Vasc. Boil. 2010, 30, 1562–1568. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kong, R.; Ma, Y.; Feng, J.; Li, S.; Zhang, W.; Jiang, J.; Zhang, J.; Qiao, Z.; Yang, X.; Zhou, B. The Crucial Role of miR-126 on Suppressing Progression of Esophageal Cancer by Targeting VEGF-A. Cell. Mol. Biol. Lett. 2016, 21, 3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, N.; Zhang, D.; Xie, H.; Zhou, Z.; Chen, H.; Hu, T.; Bai, Y.; Shen, Y.; Yuan, W.; Jing, Q.; et al. Endothelial-Specific Intron-Derived miR-126 is Down-Regulated in Human Breast Cancer and Targets Both VEGFA and PIK3R2. Mol. Cell. Biochem. 2011, 351, 157–164. [Google Scholar] [CrossRef] [PubMed]
- Huang, T.H.; Chu, T.Y. Repression of miR-126 and Upregulation of Adrenomedullin in the Stromal Endothelium by Cancer-Stromal Cross Talks Confers Angiogenesis of Cervical Cancer. Oncogene 2014, 33, 3636–3647. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Zhou, H.; Wu, D.; Ni, H.; Chen, Z.; Chen, C.; Xiang, Y.; Dai, K.; Chen, X.; Li, X. MicroRNA let-7a Regulates Angiogenesis by Targeting TGFBR3 mRNA. J. Cell. Mol. Med. 2019, 23, 556–567. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sureban, S.M.; Madhoun, M.F.; May, R.; Qu, D.; Ali, N.; Fazili, J.; Weygant, N.; Chandrakesan, P.; Ding, K.; Lightfoot, S.A.; et al. Plasma DCLK1 is a Marker of Hepatocellular Carcinoma (HCC): Targeting DCLK1 Prevents HCC Tumor Xenograft Growth Via a microRNA-Dependent Mechanism. Oncotarget 2015, 6, 37200–37215. [Google Scholar] [CrossRef]
- Sureban, S.M.; May, R.; Qu, D.; Weygant, N.; Chandrakesan, P.; Ali, N.; Lightfoot, S.A.; Pantazis, P.; Rao, C.V.; Postier, R.G.; et al. DCLK1 Regulates Pluripotency and Angiogenic Factors Via microRNA-Dependent Mechanisms in Pancreatic Cancer. PLoS ONE 2013, 8, e73940. [Google Scholar] [CrossRef]
- Sureban, S.M.; May, R.; Weygant, N.; Qu, D.; Chandrakesan, P.; Bannerman Menson, E.; Ali, N.; Pantazis, P.; Westphalen, C.B.; Wang, T.C.; et al. XMD8-92 Inhibits Pancreatic Tumor Xenograft Growth Via a DCLK1-Dependent Mechanism. Cancer Lett. 2014, 351, 151–161. [Google Scholar] [CrossRef]
- Jeyapalan, Z.; Deng, Z.; Shatseva, T.; Fang, L.; He, C.; Yang, B.B. Expression of CD44 3’-Untranslated Region Regulates Endogenous microRNA Functions in Tumorigenesis and Angiogenesis. Nucleic Acids Res. 2011, 39, 3026–3041. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, Z.; Liu, F.; Zhang, H.; Li, X.; Li, Y.; Li, J.; Liu, F.; Cao, Y.; Cao, L.; Li, F. miR-135a Inhibits Tumor Metastasis and Angiogenesis by Targeting FAK Pathway. Oncotarget 2017, 8, 31153–31168. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, Y.; Li, S.; Li, J.; Wang, D.; Li, Q. Effect of microRNA-135a on Cell Proliferation, Migration, Invasion, Apoptosis and Tumor Angiogenesis Through the IGF-1/PI3K/Akt Signaling Pathway in Non-Small Cell Lung Cancer. Cell. Physiol. Biochem. 2017, 42, 1431–1446. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.X.; Xu, X.X.; Tan, B.Z.; Zhang, Z.; Zhou, X.D. MicroRNA-29b Inhibits Angiogenesis by Targeting VEGFA Through the MAPK/ERK and PI3K/Akt Signaling Pathways in Endometrial Carcinoma. Cell. Physiol. Biochem. 2017, 41, 933–946. [Google Scholar] [CrossRef] [PubMed]
- Fang, J.H.; Zhou, H.C.; Zeng, C.; Yang, J.; Liu, Y.; Huang, X.; Zhang, J.P.; Guan, X.Y.; Zhuang, S.M. MicroRNA-29b Suppresses Tumor Angiogenesis, Invasion, and Metastasis by Regulating Matrix Metalloproteinase 2 Expression. Hepatology 2011, 54, 1729–1740. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Cai, B.; Shen, L.; Dong, Y.; Lu, Q.; Sun, S.; Liu, S.; Ma, S.; Ma, P.X.; Chen, J. MiRNA-29b Suppresses Tumor Growth Through Simultaneously Inhibiting Angiogenesis and Tumorigenesis by Targeting Akt3. Cancer Lett. 2017, 397, 111–119. [Google Scholar] [CrossRef]
- Zhao, X.; Liu, Y.; Li, Z.; Zheng, S.; Wang, Z.; Li, W.; Bi, Z.; Li, L.; Jiang, Y.; Luo, Y.; et al. Linc00511 Acts as a Competing Endogenous RNA to Regulate VEGFA Expression Through Sponging Hsa-Mir-29b-3p in Pancreatic Ductal Adenocarcinoma. J. Cell. Mol. Med. 2018, 22, 655–667. [Google Scholar] [CrossRef]
- Zhang, T.; Liu, M.; Wang, C.; Lin, C.; Sun, Y.; Jin, D. Down-Regulation of MiR-206 Promotes Proliferation and Invasion of Laryngeal Cancer by Regulating VEGF Expression. Anticancer. Res. 2011, 31, 3859–3863. [Google Scholar]
- Liang, Z.; Bian, X.; Shim, H. Downregulation of microRNA-206 Promotes Invasion and Angiogenesis of Triple Negative Breast Cancer. Biochem. Biophys. Res. Commun. 2016, 477, 461–466. [Google Scholar] [CrossRef] [Green Version]
- Xue, D.; Yang, Y.; Liu, Y.; Wang, P.; Dai, Y.; Liu, Q.; Chen, L.; Shen, J.; Ju, H.; Li, Y.; et al. MicroRNA-206 Attenuates the Growth and Angiogenesis in Non-Small Cell Lung Cancer Cells by Blocking the 14-3-3ζ/STAT3/HIF-1α/VEGF Signaling. Oncotarget 2016, 7, 79805–79813. [Google Scholar] [CrossRef]
- Chen, Q.Y.; Jiao, D.M.; Wu, Y.Q.; Chen, J.; Wang, J.; Tang, X.L.; Mou, H.; Hu, H.Z.; Song, J.; Yan, J.; et al. MiR-206 Inhibits HGF-Induced Epithelial-Mesenchymal Transition and Angiogenesis in Non-Small Cell Lung Cancer Via c-Met/PI3k/Akt/mTOR Pathway. Oncotarget 2016, 7, 18247–18261. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, Y.; Li, Y.; Wu, C.; Zhou, L.; Han, X.; Wang, Q.; Xie, X.; Zhou, Y.; Du, Z. MicroRNA-140-5p inhibits Cell Proliferation and Invasion by Regulating VEGFA/MMP2 Signaling in Glioma. Tumor Boil. 2017, 39, 1010428317697558. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, W.; Zou, C.; Pan, L.; Xu, Y.; Qi, W.; Ma, G.; Hou, Y.; Jiang, P. MicroRNA-140-5p Inhibits the Progression of Colorectal Cancer by Targeting VEGFA. Cell. Physiol. Biochem. 2015, 37, 1123–1133. [Google Scholar] [CrossRef] [PubMed]
- Gong, J.; Zhu, S.; Zhang, Y.; Wang, J. Interplay of VEGFa and MMP2 Regulates Invasion of Glioblastoma. Tumor Boil. 2014, 35, 11879–11885. [Google Scholar] [CrossRef]
- Qiu, Y.; Yu, H.; Shi, X.; Xu, K.; Tang, Q.; Liang, B.; Hu, S.; Bao, Y.; Cai, J.; Peng, W.; et al. microRNA-497 Inhibits Invasion and Metastasis of Colorectal Cancer Cells by Targeting Vascular Endothelial Growth Factor-A. Cell Prolif. 2016, 49, 69–78. [Google Scholar] [CrossRef]
- Wu, Z.; Cai, X.; Huang, C.; Xu, J.; Liu, A. miR-497 Suppresses Angiogenesis in Breast Carcinoma by Targeting HIF-1alpha. Oncol. Rep. 2016, 35, 1696–1702. [Google Scholar] [CrossRef]
- Pengcheng, S.; Ziqi, W.; Luyao, Y.; Xiangwei, Z.; Liang, L.; Yuwei, L.; Lechen, L.; Wanhai, X. MicroRNA-497 Suppresses Renal Cell Carcinoma by Targeting VEGFR-2 in ACHN cells. Biosci. Rep. 2017, 37, 37. [Google Scholar] [CrossRef]
- Li, B.; Xu, W.W.; Han, L.; Chan, K.T.; Tsao, S.W.; Lee, N.P.Y.; Law, S.; Xu, L.Y.; Li, E.M.; Chan, K.W.; et al. MicroRNA-377 Suppresses Initiation and Progression of Esophageal Cancer by Inhibiting CD133 and VEGF. Oncogene 2017, 36, 3986–4000. [Google Scholar] [CrossRef]
- Zhang, R.; Luo, H.; Wang, S.; Chen, W.; Chen, Z.; Wang, H.W.; Chen, Y.; Yang, J.; Zhang, X.; Wu, W.; et al. MicroRNA-377 Inhibited Proliferation and Invasion of Human Glioblastoma Cells by Directly Targeting Specificity Protein 1. Neuro Oncol. 2014, 16, 1510–1522. [Google Scholar] [CrossRef] [Green Version]
- Guan, B.; Wu, K.; Zeng, J.; Xu, S.; Mu, L.; Gao, Y.; Wang, K.; Ma, Z.; Tian, J.; Shi, Q.; et al. Tumor-Suppressive microRNA-218 Inhibits Tumor Angiogenesis Via Targeting the mTOR Component RICTOR in Prostate Cancer. Oncotarget 2017, 8, 8162–8172. [Google Scholar] [CrossRef] [Green Version]
- Tang, S.; Wang, D.; Zhang, Q.; Li, L. miR-218 Suppresses Gastric Cancer Cell Proliferation and Invasion Via Regulation of Angiopoietin-2. Exp. Ther. Med. 2016, 12, 3837–3842. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, X.; Dong, J.; He, Y.; Zhao, M.; Liu, Z.; Wang, N.; Jiang, M.; Zhang, Z.; Liu, G.; Liu, H.; et al. miR-218 Inhibited Tumor Angiogenesis by Targeting ROBO1 in Gastric Cancer. Gene 2017, 615, 42–49. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Lv, Z.; Xu, J.; Chen, C.; Ge, Q.; Li, P.; Wei, D.; Wu, Z.; Sun, X. Micro RNA-134 Inhibits Osteosarcoma Angiogenesis and Proliferation by Targeting the VEGFA/VEGFR 1 Pathway. FEBS J. 2018, 285, 1359–1371. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ivan, M.; Kondo, K.; Yang, H.; Kim, W.; Valiando, J.; Ohh, M.; Salic, A.; Asara, J.M.; Lane, W.S.; Kaelin, W.G. HIFalpha Targeted for VHL-Mediated Destruction by Proline Hydroxylation: Implications for O2 Sensing. Science 2001, 292, 464–468. [Google Scholar] [CrossRef] [PubMed]
- Kurban, G.; Sundberg, T.B.; Ney, G.M.; Subramanian, C.; Opipari, A.W.; Glick, G.D. Characterization of a Von Hippel Lindau Pathway Involved in Extracellular Matrix Remodeling, Cell Invasion, and Angiogenesis. Cancer Res. 2006, 66, 1313–1319. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matsuura, Y.; Wada, H.; Eguchi, H.; Gotoh, K.; Kobayashi, S.; Kinoshita, M.; Kubo, M.; Hayashi, K.; Iwagami, Y.; Yamada, D.; et al. Exosomal miR-155 Derived from Hepatocellular Carcinoma Cells Under Hypoxia Promotes Angiogenesis in Endothelial Cells. Dig. Dis. Sci. 2019, 64, 792–802. [Google Scholar] [CrossRef]
- Corn, P.G. Hypoxic Regulation of miR-210: Shrinking Targets Expand HIF-1’s Influence. Cancer Boil. Ther. 2008, 7, 265–267. [Google Scholar] [CrossRef] [Green Version]
- Lu, Y.; Thomson, J.M.; Wong, H.Y.F.; Hammond, S.M.; Hogan, B.L. Transgenic Over-Expression of the microRNA miR-17-92 Cluster Promotes Proliferation and Inhibits Differentiation of Lung Epithelial Progenitor Cells. Dev. Boil. 2007, 310, 442–453. [Google Scholar] [CrossRef] [Green Version]
- Dews, M.; Homayouni, A.; Yu, D.; Murphy, D.; Sevignani, C.; Wentzel, E.; E Furth, E.; Lee, W.M.; Enders, G.H.; Mendell, J.T.; et al. Augmentation of Tumor Angiogenesis by a Myc-Activated microRNA Cluster. Nat. Genet. 2006, 38, 1060–1065. [Google Scholar] [CrossRef] [Green Version]
- O’Donnell, K.A.; Wentzel, E.A.; Zeller, K.I.; Dang, C.V.; Mendell, J.T. c-Myc-Regulated microRNAs Modulate E2F1 Expression. Nature 2005, 435, 839–843. [Google Scholar] [CrossRef]
- Amini, S.; Abak, A.; Sakhinia, E.; Abhari, A. MicroRNA-221 and MicroRNA-222 in Common Human Cancers: Expression, Function, and Triggering of Tumor Progression as a Key Modulator. Lab. Med. 2019, 50, 334–347. [Google Scholar] [CrossRef]
- Fish, J.E.; Santoro, M.M.; Morton, S.U.; Yu, S.; Yeh, R.F.; Wythe, J.D.; Ivey, K.N.; Bruneau, B.G.; Stainier, D.Y.R.; Srivastava, D. miR-126 Regulates Angiogenic Signaling and Vascular Integrity. Dev. Cell 2008, 15, 272–284. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, S.; Aurora, A.B.; Johnson, B.A.; Qi, X.; McAnally, J.; Hill, J.A.; Richardson, J.A.; Bassel Duby, R.; Olson, E.N. The Endothelial-Specific microRNA miR-126 Governs Vascular Integrity and Angiogenesis. Dev. Cell 2008, 15, 261–271. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fox, S.B.; Fawcett, J.; Jackson, D.G.; Collins, I.; Gatter, K.C.; Harris, A.L.; Gearing, A.; Simmons, D.L. Normal Human Tissues, in Addition to Some Tumors, Express Multiple Different CD44 Isoforms. Cancer Res. 1994, 54, 4539–4546. [Google Scholar] [PubMed]
- Taher, T.E.I.; Van Der Voort, R.; Smit, L.; Keehnen, R.M.J.; Schilder Tol, E.J.M.; Spaargaren, M.; Pals, S.T. Cross-Talk Between CD44 and c-Met in B Cells. Curr. Top. Microbiol. Immunol. 1999, 246, 31–38. [Google Scholar]
- Weber, G.F.; Ashkar, S.; Glimcher, M.J.; Cantor, H. Receptor-Ligand Interaction Between CD44 and Osteopontin (Eta-1). Science 1996, 271, 509–512. [Google Scholar] [CrossRef]
- Ferrucci, A.; Moschetta, M.; Frassanito, M.A.; Berardi, S.; Catacchio, I.; Ria, R.; Racanelli, V.; Caivano, A.; Solimando, A.G.; Vergara, D.; et al. A HGF/cMET Autocrine Loop Is Operative in Multiple Myeloma Bone Marrow Endothelial Cells and May Represent a Novel Therapeutic Target. Clin. Cancer Res. 2014, 20, 5796–5807. [Google Scholar] [CrossRef]
- Shi, Q.; Le, X.; Abbruzzese, J.L.; Peng, Z.; Qian, C.N.; Tang, H.; Xiong, Q.; Wang, B.; Li, X.C.; Xie, K. Constitutive Sp1 Activity is Essential for Differential Constitutive Expression of Vascular Endothelial Growth Factor in Human Pancreatic Adenocarcinoma. Cancer Res. 2001, 61, 4143–4154. [Google Scholar]
- Yao, J.C.; Nordal, R.A.; Nagy, A.; Pintilie, M.; Wong, C.S. Association Between Expression of Transcription Factor Sp1 and Increased Vascular Endothelial Growth Factor Expression, Advanced Stage, and Poor Survival in Patients with Resected Gastric Cancer. Clin. Cancer Res. 2004, 10, 4109–4117. [Google Scholar] [CrossRef] [Green Version]
- Lou, Z.; O’Reilly, S.; Liang, H.; Maher, V.M.; Sleight, S.D.; McCormick, J.J. Down-Regulation of Overexpressed sp1 Protein in Human Fibrosarcoma Cell Lines Inhibits Tumor Formation. Cancer Res. 2005, 65, 1007–1017. [Google Scholar]
- Casanovas, O. Cancer: Limitations of Therapies Exposed. Nature 2012, 484, 44–46. [Google Scholar] [CrossRef] [PubMed]
- Jaszai, J.; Schmidt, M.H. Trends and Challenges in Tumor Anti-Angiogenic Therapies. Cells 2019, 8, 1102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Romano, G.; Kwong, L.N. Diagnostic and Therapeutic Applications of miRNA-Based Strategies to Cancer Immunotherapy. Cancer Metastasis Rev. 2018, 37, 45–53. [Google Scholar] [CrossRef] [PubMed]
- Kelderman, S.; Schumacher, T.N.; Haanen, J.B. Acquired and Intrinsic Resistance in Cancer Immunotherapy. Mol. Oncol. 2014, 8, 1132–1139. [Google Scholar] [CrossRef]
- O’Donnell, J.S.; Smyth, M.J.; Teng, M.W.L. Acquired Resistance to anti-PD1 Therapy: Checkmate to Checkpoint Blockade? Genome Med. 2016, 8, 111. [Google Scholar] [CrossRef] [Green Version]
- Sharma, P.; Hu Lieskovan, S.; Wargo, J.A.; Ribas, A. Primary, Adaptive, and Acquired Resistance to Cancer Immunotherapy. Cell 2017, 168, 707–723. [Google Scholar] [CrossRef] [Green Version]
- Hansen, T.F.; Carlsen, A.L.; Heegaard, N.H.H.; Sorensen, F.B.; Jakobsen, A. Changes in Circulating microRNA-126 During Treatment with Chemotherapy and Bevacizumab Predicts Treatment Response in Patients with Metastatic Colorectal Cancer. Br. J. Cancer 2015, 112, 624–629. [Google Scholar] [CrossRef]
- Merhautova, J.; Hezova, R.; Poprach, A.; Kovarikova, A.; Radova, L.; Svoboda, M.; Vyzula, R.; Demlova, R.; Slaby, O. miR-155 and miR-484 Are Associated with Time to Progression in Metastatic Renal Cell Carcinoma Treated with Sunitinib. BioMed Res. Int. 2015, 2015, 941980. [Google Scholar] [CrossRef] [Green Version]
- Anand, S.; Majeti, B.K.; Acevedo, L.M.; Acevedo, M.E.; Mukthavaram, R.; Scheppke, L.; Huang, M.; Shields, D.J.; Lindquist, J.N.; Lapinski, P.E.; et al. MicroRNA-132-Mediated Loss of p120RasGAP Activates the Endothelium to Facilitate Pathological Angiogenesis. Nat. Med. 2010, 16, 909–914. [Google Scholar] [CrossRef]
- Babae, N.; Bourajjaj, M.; Liu, Y.; Van Beijnum, J.R.; Cerisoli, F.; Scaria, P.V.; Verheul, M.; Van Berkel, M.P.; Pieters, E.H.E.; Van Haastert, R.J.; et al. Systemic miRNA-7 Delivery Inhibits Tumor Angiogenesis and Growth in Murine Xenograft Glioblastoma. Oncotarget 2014, 5, 6687–6700. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.Q.; Song, W.J.; Sun, T.M.; Zhang, P.Z.; Wang, J. Targeted Delivery of Antisense Inhibitor of miRNA for Antiangiogenesis Therapy Using cRGD-Functionalized Nanoparticles. Mol. Pharm. 2011, 8, 250–259. [Google Scholar] [CrossRef] [PubMed]
- Muramatsu, F.; Kidoya, H.; Naito, H.; Sakimoto, S.; Takakura, N. microRNA-125b Inhibits Tube Formation of Blood Vessels Through Translational Suppression of VE-Cadherin. Oncogene 2013, 32, 414–421. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, H.; Bai, M.; Deng, T.; Liu, R.; Wang, X.; Qu, Y.; Duan, J.; Zhang, L.; Ning, T.; Ge, S.; et al. Cell-Derived Microvesicles Mediate the Delivery of miR-29a/c to Suppress Angiogenesis in Gastric Carcinoma. Cancer Lett. 2016, 375, 331–339. [Google Scholar] [CrossRef] [PubMed]


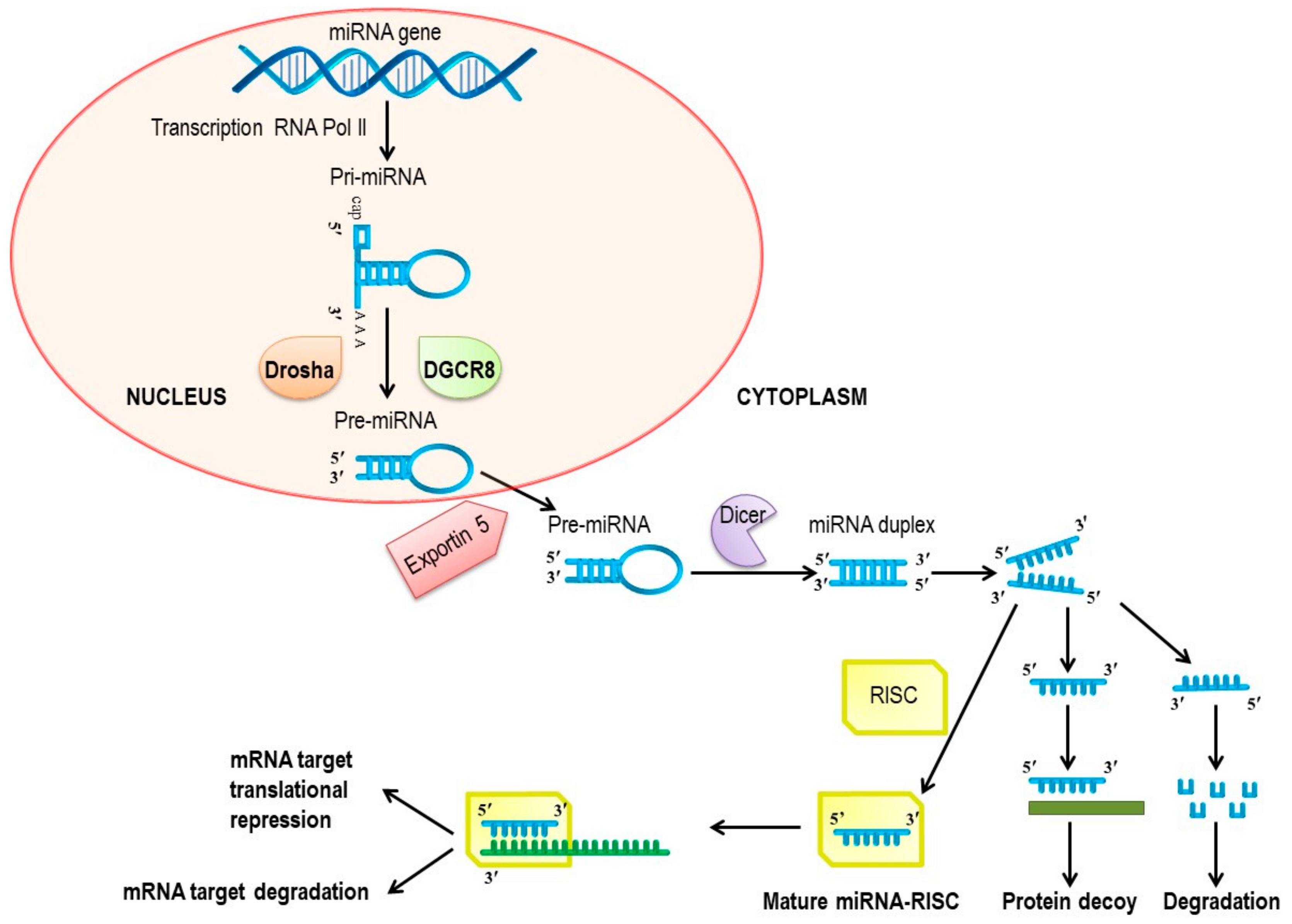{kind=link}
| miRNAs | Targets | Tumor | Function | References |
|---|---|---|---|---|
| miR-155 | VHL | Triple negative breast cancer, renal cell carcinoma, melanoma cell lines | ↑ proliferation, invasion, migration and tube formation ↑ VEGF-A, FGF-2 and MMP9 | [44,45,46,47] |
| miR-566 | VHL | Glioblastoma, glioma cell line | ↑ EGFR pathway, invasion and migration ↑ VEGF, HIF-1α | [48,49] |
| miR-210 | EFNA3 FGFRL1 | Breast cancer, ovarian cancer, HBV-related hepatocellular carcinoma | ↑ Notch pathway ↑ endothelial cell migration and capillary-like structure formation | [50,51,52,53,54] |
| miR-21 | PTEN Pdcd4 | Prostate cancer cell lines, glioma, pancreatic cancer cell lines, breast cancer, hepatocellular carcinoma, colorectal cancer | ↑ AKT and ERK1/2 signaling pathways ↑ VEGF, HIF-1α | [55,56,57,58,59,60] |
| miR-182 | FBXW7 PHD FIH1 | Breast cancer, prostate cancer | ↑ VEGF, HIF-1α | [61,62] |
| miR-296 | HGS | Glioma | ↑ VEGFR-2, PDGFRβ | [63] |
| miR-17-92 | Tsp1 CTGF PTEN HIF-1α | Lung cancer cell lines | Endothelial cell activation | [64,65] |
| let-7b/-7f | TIMP Tsp1/2 | Prostate cancer | ↑ endothelial cell migration | [66,67,68] |
| miR-378 | ALCAM EHD1 TLK1 ELK3 Sufu Fus-1 | Ovarian cancer, non-small cell lung cancer, glioblastoma cell lines, bladder cancer | endothelial cell function regulation ↑ VEGF activity | [69,70,71,72] |
| miR-221/222 | SOCS3 TIMP2 | Glioblastoma, glioma | ↑ MMP2, MMP9, VEGF ↑ tube formation ↑ JAK/STAT pathway | [73,74] |
| miRNAs | Targets | Tumor | Function | References |
|---|---|---|---|---|
| miR-221/222 | c-Kit VEGFR-2 ETS-1 PIK3R1 CDKN1B ZEB2 STAT5A | Metastatic renal cell carcinoma, breast cancer | ↓ endothelial cell migration, proliferation, and ability to form new capillaries ↓ VEGFR-2 | [75,76,77,78,79,80,81,82,83] |
| miR-126 | VEGF-A PIK3R2 ADM | Esophageal cancer, breast cancer, cervical cancer | ↓ VEGF/PI3K/AKT signaling ↓ Angiogenesis and microvessel density | [84,85,86] |
| let-7a | TGFBR3 | Pancreatic cancer, hepatocellular carcinoma | ↓ tube formation and migration of endothelial cells | [87,88,89,90] |
| miR-328 | CD44 | Breast cancer MT1cell line | ↓ endothelial cell activity, tubulogenesis and blood vessel formation | [91] |
| miR-135a | FAK IGF-1/PI3K/Akt pathway | Gastric cancer, non-small cell lung cancer | ↓ VEGF signaling | [92,93] |
| miR-29b | VEGF ERK Akt MMP-2 | endometrial carcinoma, breast cancer, pancreatic ductal adenocarcinoma and hepatocellular carcinoma | ↓ VEGF expression ↓ MAPK/ERK and PI3K/Akt signaling pathways ↓ MMP-2 expression and VEGFR-2 signaling | [94,95,96,97] |
| miR-206 | VEGF MAPK3 SOX914-3-3ζ/STAT3/HIF-1α/VEGF signaling c-Met/PI3k/Akt/mTOR signaling | laryngeal squamous cell carcinoma, triple negative breast cancer, non-small cell lung cancer | ↓ VEGF expression ↓ endothelial cell migration and capillary tube formation | [98,99,100,101] |
| miR-140-5p | VEGF-A | colorectal cancer, glioma, glioblastoma | ↓ VEGF-A/MMP2 signaling | [102,103,104] |
| miR-497 | VEGF-A HIF-1α VEGFR-2 | colorectal cancer, breast cancer; renal carcinoma | ↓ VEGF-A/ERK/MMP-9 signaling ↓ tube formation and branch points ↓ MEK/ERK and p38 MAPK pathways | [105,106,107] |
| miR-377 | VEGF CD133 Sp1 | esophageal squamous cell carcinoma, glioblastoma | ↓ VEGF | [108,109] |
| miR-218 | RICTOR Angiopoietin-2 ROBO1 | Prostate cancer, gastric cancer | ↓ RICTOR/mTOR/HIF-1/VEGF signaling pathway ↓ endothelial cell sprouting, motility and tube formation | [110,111,112] |
| miR-134 | VEGF-A VEGFR-1 | osteosarcoma | ↓ VEGF-A/VEGFR-1-AKT signaling | [113] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Leone, P.; Buonavoglia, A.; Fasano, R.; Solimando, A.G.; De Re, V.; Cicco, S.; Vacca, A.; Racanelli, V. Insights into the Regulation of Tumor Angiogenesis by Micro-RNAs. J. Clin. Med. 2019, 8, 2030. https://doi.org/10.3390/jcm8122030
Leone P, Buonavoglia A, Fasano R, Solimando AG, De Re V, Cicco S, Vacca A, Racanelli V. Insights into the Regulation of Tumor Angiogenesis by Micro-RNAs. Journal of Clinical Medicine. 2019; 8(12):2030. https://doi.org/10.3390/jcm8122030
Chicago/Turabian StyleLeone, Patrizia, Alessio Buonavoglia, Rossella Fasano, Antonio Giovanni Solimando, Valli De Re, Sebastiano Cicco, Angelo Vacca, and Vito Racanelli. 2019. "Insights into the Regulation of Tumor Angiogenesis by Micro-RNAs" Journal of Clinical Medicine 8, no. 12: 2030. https://doi.org/10.3390/jcm8122030
APA StyleLeone, P., Buonavoglia, A., Fasano, R., Solimando, A. G., De Re, V., Cicco, S., Vacca, A., & Racanelli, V. (2019). Insights into the Regulation of Tumor Angiogenesis by Micro-RNAs. Journal of Clinical Medicine, 8(12), 2030. https://doi.org/10.3390/jcm8122030