Human Colorectal Cancer Infrastructure Constructed by the Glycocalyx

 ,
,  ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Methods
2.1. Human Specimens and Mice
2.2. Tissue Preparation
2.3. Immunohistochemistry for CD31 and Periodic Acid-Schiff (PAS) Double Staining
2.4. Azan Staining
2.5. VM Quantification
2.6. Immunofluorescence (IF) and Lectin Staining
2.7. Electron Microscopy
2.8. Quantitative Assessment of the GCX
2.9. Statistical Analysis
3. Results
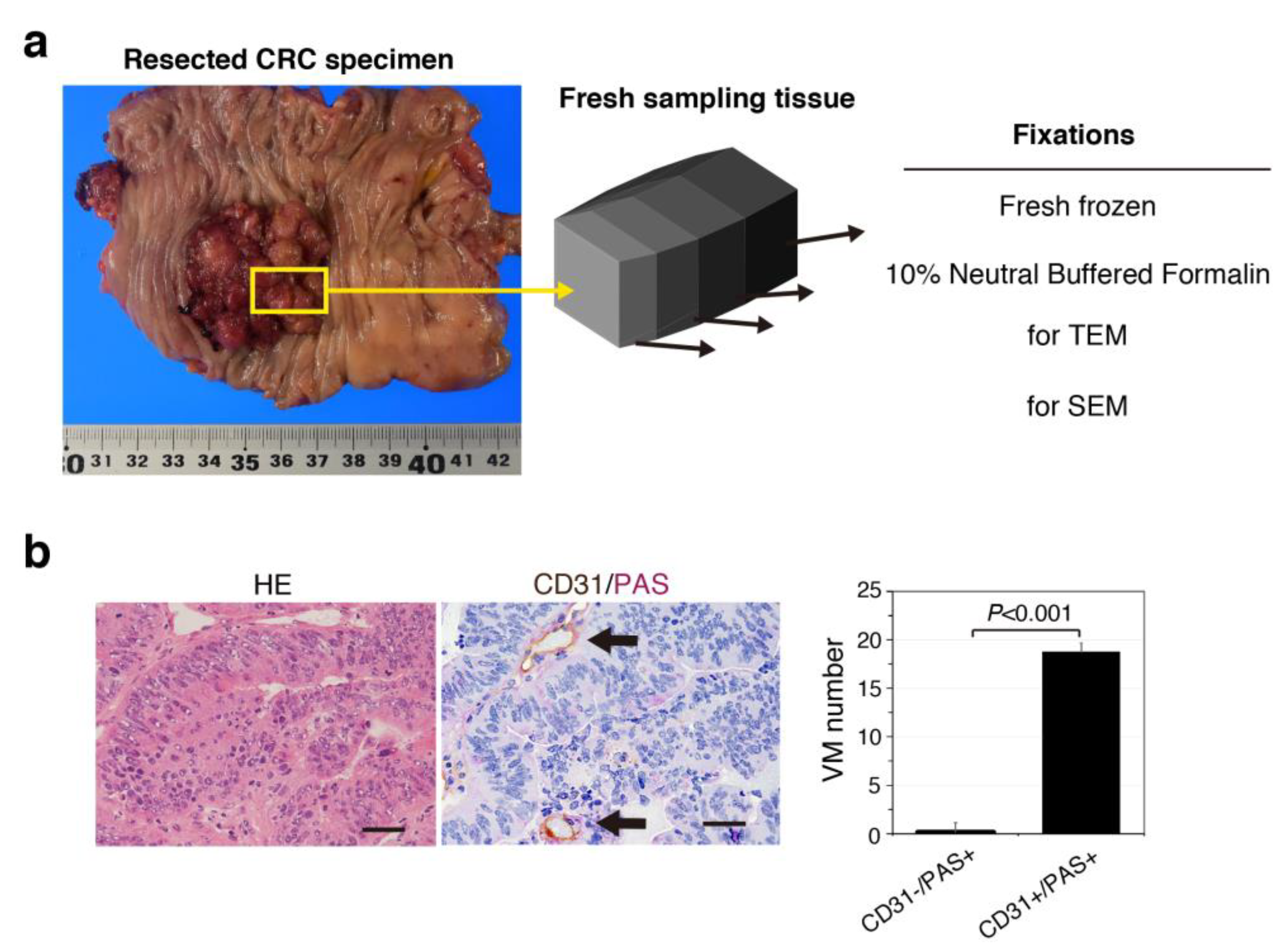
3.1. VM Formation Defined by CD31/PAS Staining Is Uncommon in Patient-Derived CRC Tissues
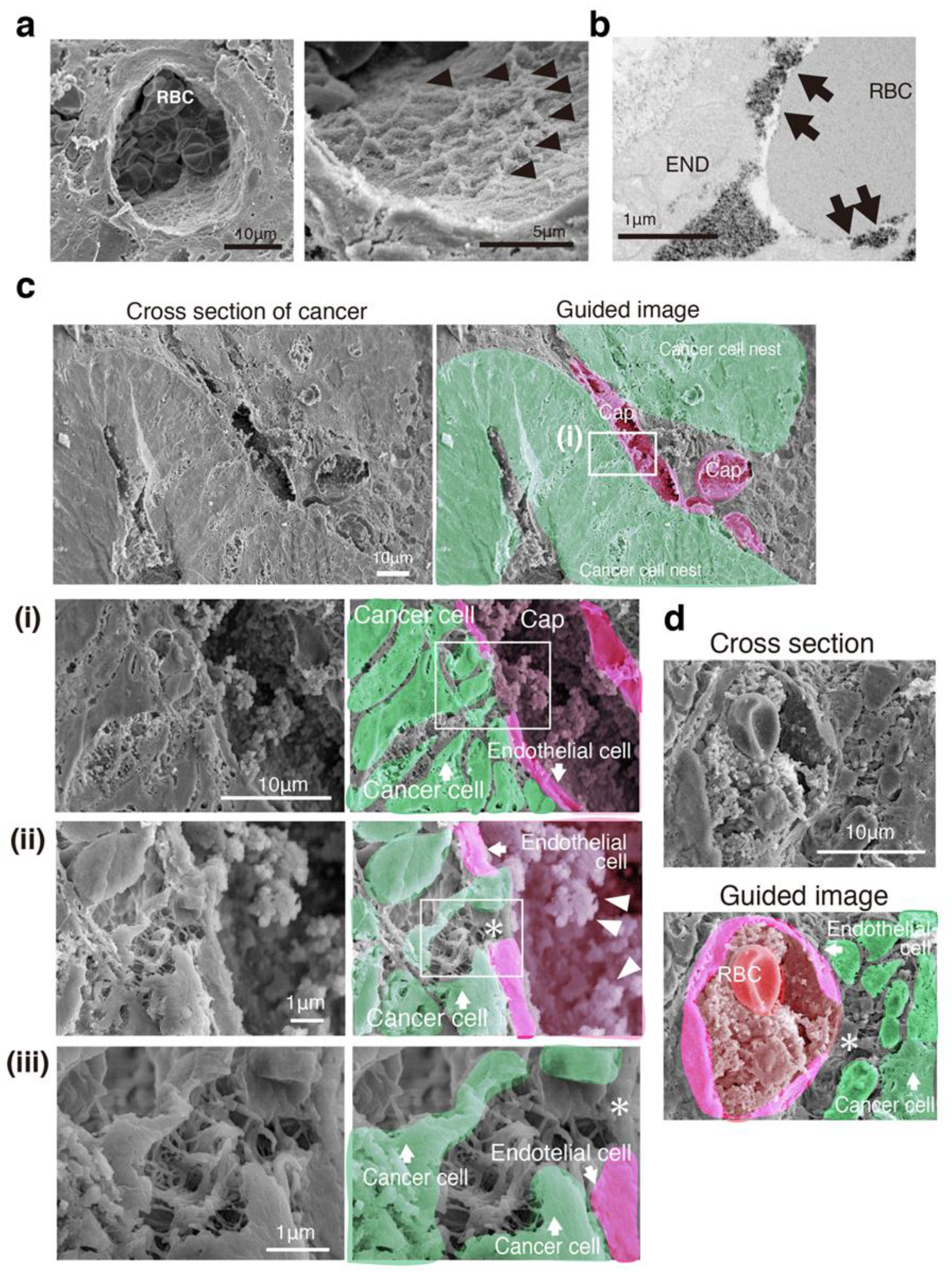
3.2. SEM Imaging Visualizes Cancer Intercellular Tunnels Connected to Small Capillaries
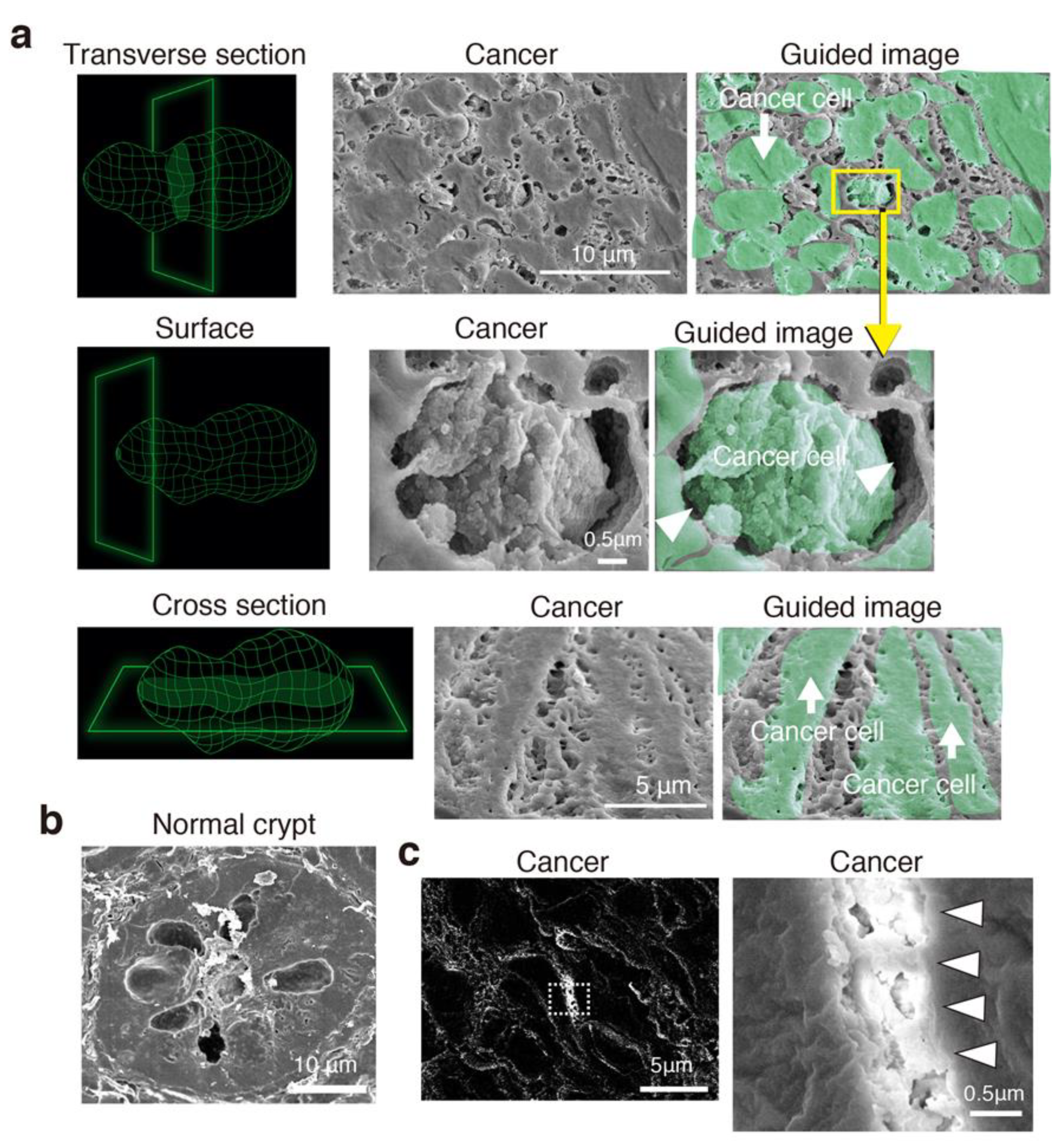
3.3. Cancer Intercellular Tunnels Are Wider than Those of Normal Epithelium
3.4. CRC Has Intercellular Tunnels with GCX Even after Considering Sample Preparation Artifacts
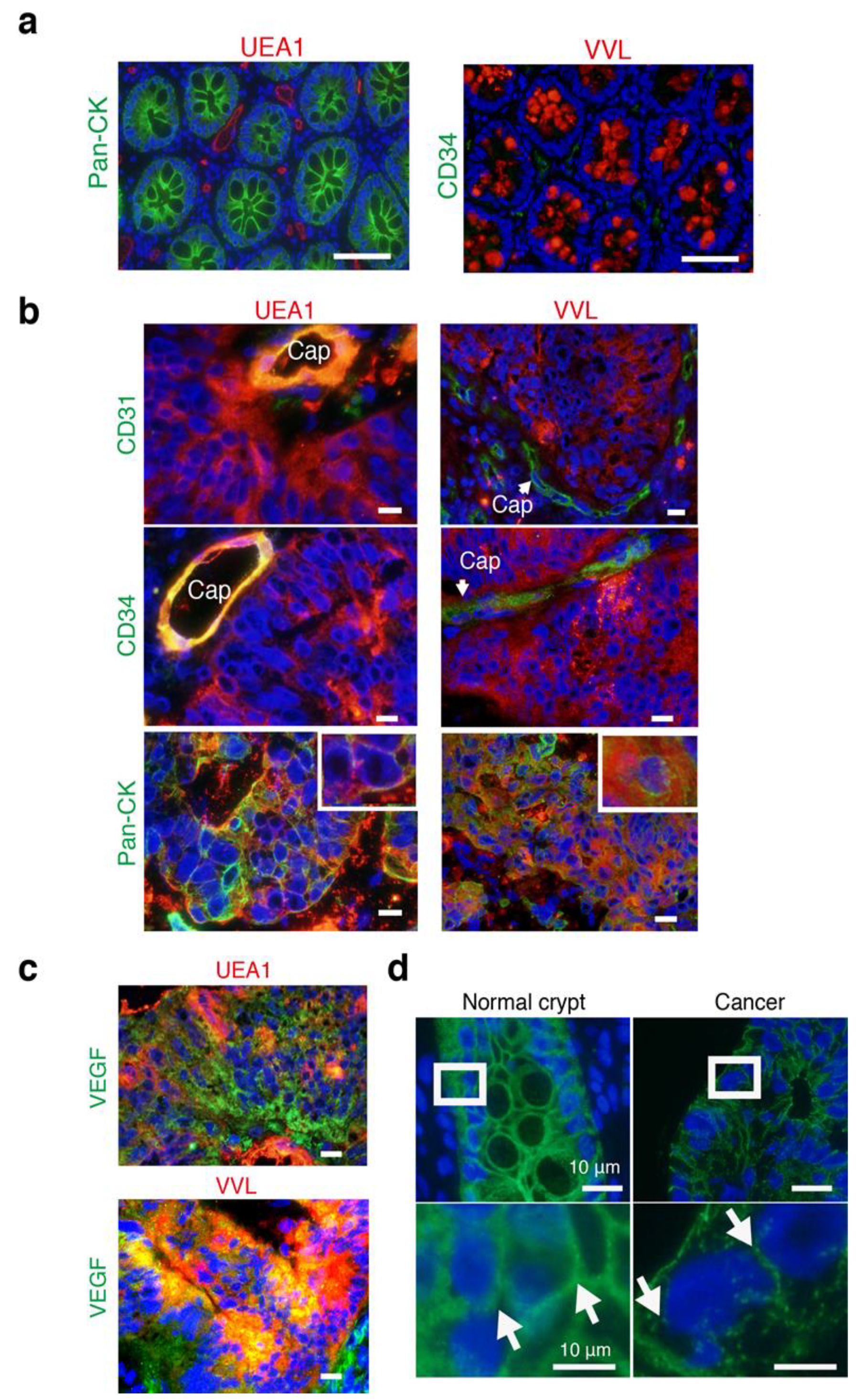
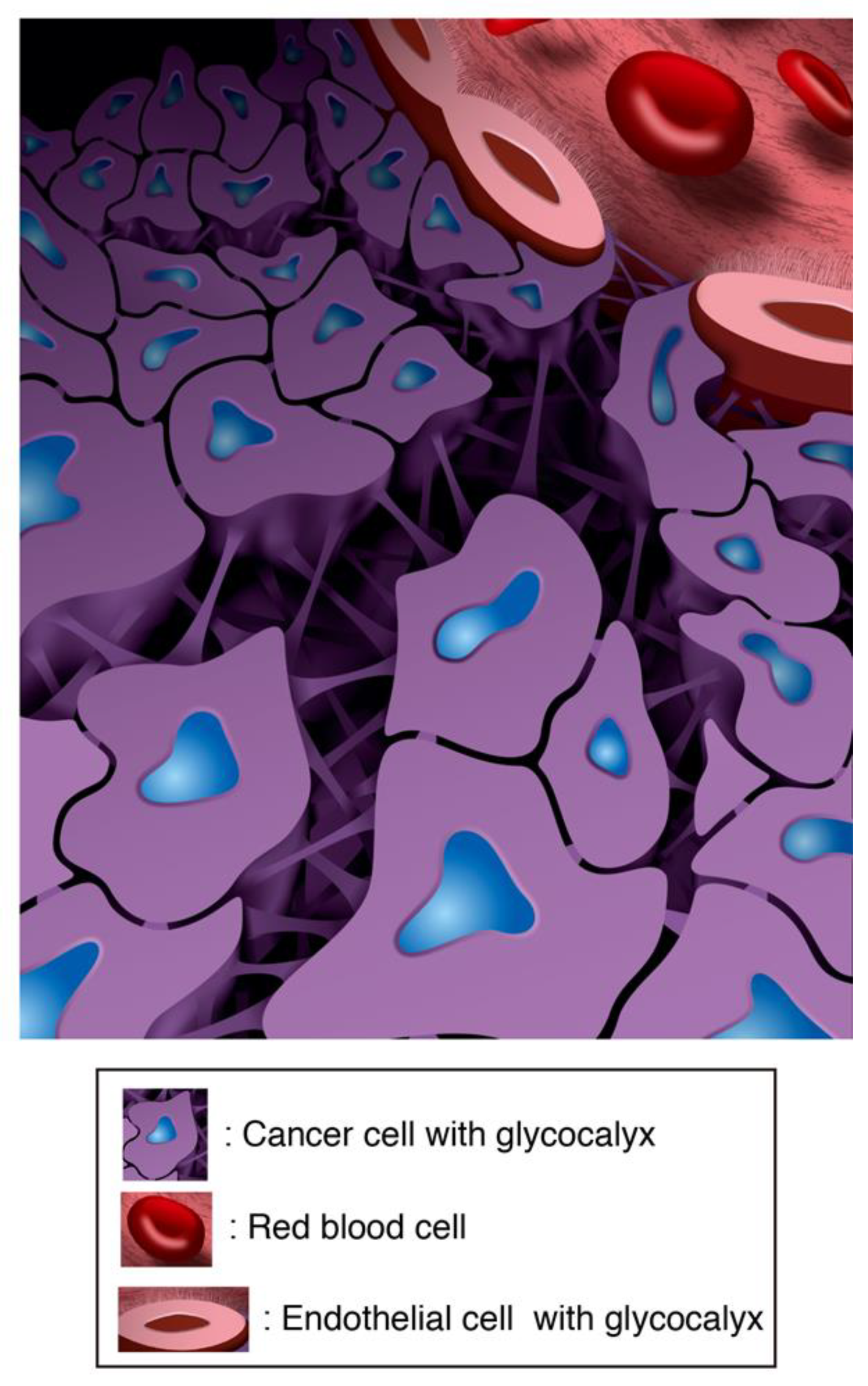
3.5. The Intercellular Tunnels of Cancer Tissue Contain Rich Sugar, the Component of Cancer-Specific GCX
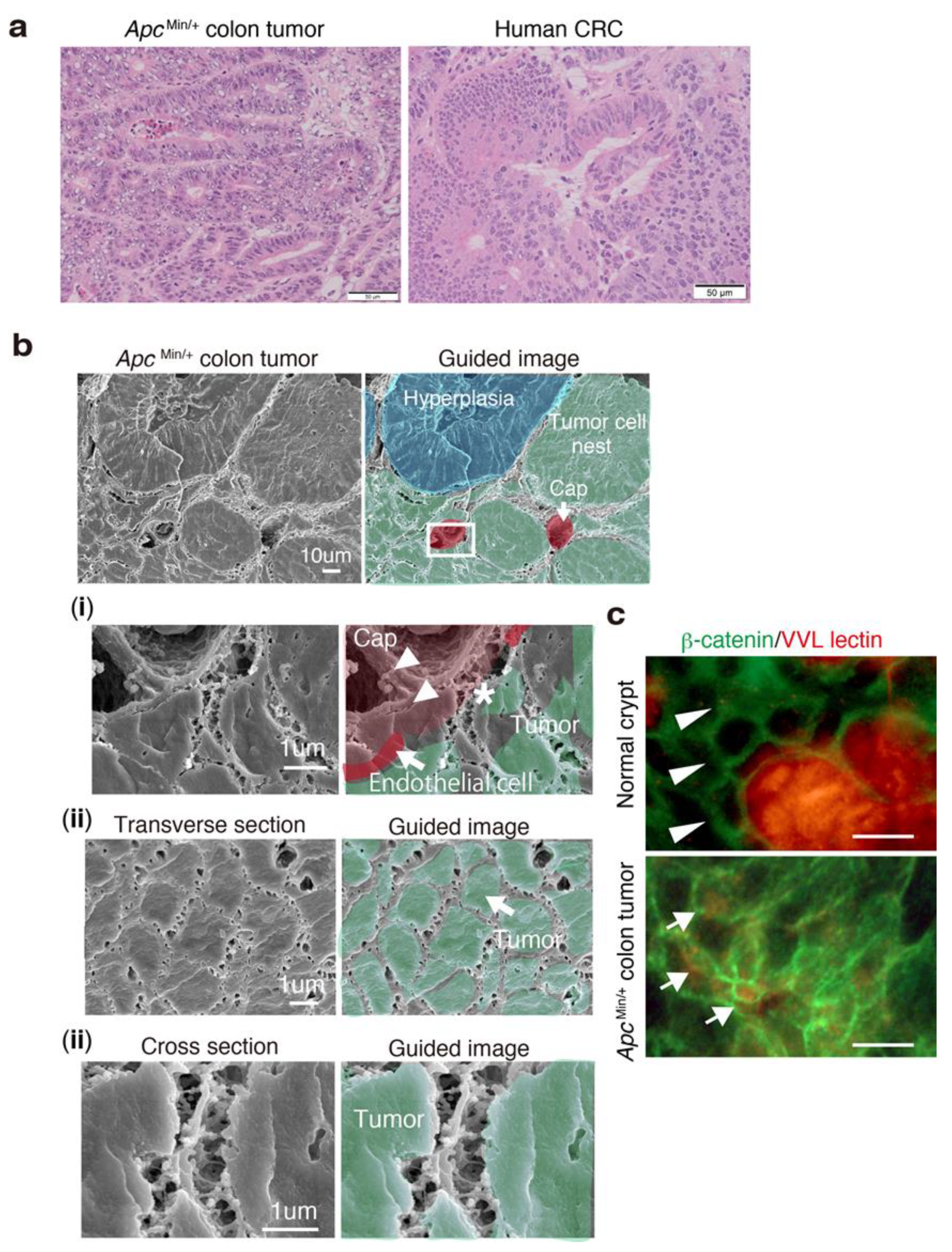
3.6. 3D Ultrastructure of Mouse Colon Cancer Phenocopies Human CRC
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Carmeliet, P.; Jain, R.K. Molecular mechanisms and clinical applications of angiogenesis. Nature 2011, 473, 298–307. [Google Scholar] [CrossRef] [Green Version]
- Folberg, R.; Maniotis, A.J. Vasculogenic mimicry. APMIS 2004, 112, 508–525. [Google Scholar] [CrossRef]
- Hendrix, M.J.; Seftor, E.A.; Seftor, R.E.; Chao, J.T.; Chien, D.S.; Chu, Y.W. Tumor cell vascular mimicry: Novel targeting opportunity in melanoma. Pharmacol. Ther. 2016, 159, 83–92. [Google Scholar] [CrossRef] [Green Version]
- Maniotis, A.J.; Folberg, R.; Hess, A.; Seftor, E.A.; Gardner, L.M.; Pe'er, J.; Trent, J.M.; Meltzer, P.S.; Hendrix, M.J. Vascular channel formation by human melanoma cells in vivo and in vitro: Vasculogenic mimicry. Am. J. Pathol. 1999, 155, 739–752. [Google Scholar] [CrossRef]
- Leslie, M. Tumors’ do-it-yourself blood vessels. Science 2016, 352, 1381–1383. [Google Scholar] [CrossRef]
- Chelazzi, C.; Villa, G.; Mancinelli, P.; De Gaudio, A.R.; Adembri, C. Glycocalyx and sepsis-induced alterations in vascular permeability. Crit. Care 2015, 19, 26. [Google Scholar] [CrossRef]
- Tarbell, J.M.; Cancel, L.M. The glycocalyx and its significance in human medicine. J. Intern. Med. 2016, 280, 97–113. [Google Scholar] [CrossRef] [Green Version]
- Kang, H.; Wu, Q.; Sun, A.; Liu, X.; Fan, Y.; Deng, X. Cancer Cell Glycocalyx and Its Significance in Cancer Progression. Int. J. Mol. Sci. 2018, 19, 2484. [Google Scholar] [CrossRef]
- Paszek, M.J.; DuFort, C.C.; Rossier, O.; Bainer, R.; Mouw, J.K.; Godula, K.; Hudak, J.E.; Lakins, J.N.; Wijekoon, A.C.; Cassereau, L.; et al. The cancer glycocalyx mechanically primes integrin-mediated growth and survival. Nature 2014, 511, 319–325. [Google Scholar] [CrossRef] [Green Version]
- Barnes, J.M.; Kaushik, S.; Bainer, R.O.; Sa, J.K.; Woods, E.C.; Kai, F.; Przybyla, L.; Lee, M.; Lee, H.W.; Tung, J.C.; et al. Tension-mediated glycocalyx-integrin feedback loop promotes mesenchymal-like glioblastoma. Nat. Cell Biol. 2018, 20, 1203–1214. [Google Scholar] [CrossRef]
- Qazi, H.; Palomino, R.; Shi, Z.D.; Munn, L.L.; Tarbell, J.M. Cancer cell glycocalyx mediates mechanotransduction and flow-regulated invasion. Integr. Biol. (Camb) 2013, 5, 1334–1343. [Google Scholar] [CrossRef] [Green Version]
- Okada, H.; Takemura, G.; Suzuki, K.; Oda, K.; Takada, C.; Hotta, Y.; Miyazaki, N.; Tsujimoto, A.; Muraki, I.; Ando, Y.; et al. Three-dimensional ultrastructure of capillary endothelial glycocalyx under normal and experimental endotoxemic conditions. Crit. Care 2017, 21, 261. [Google Scholar] [CrossRef]
- Ando, Y.; Okada, H.; Takemura, G.; Suzuki, K.; Takada, C.; Tomita, H.; Zaikokuji, R.; Hotta, Y.; Miyazaki, N.; Yano, H.; et al. Brain-specific ultrastrucuture of capillsry endothelial glycocalyx and its possible contribution for blood brain barrier. Sci. Rep. 2018, in press. [Google Scholar] [CrossRef]
- Inagawa, R.; Okada, H.; Takemura, G.; Suzuki, K.; Takada, C.; Yano, H.; Ando, Y.; Usui, T.; Hotta, Y.; Miyazaki, N.; et al. Ultrastructural Alteration of Pulmonary Capillary Endothelial Glycocalyx During Endotoxemia. Chest 2018, 154, 317–325. [Google Scholar] [CrossRef] [Green Version]
- MacAdam, A. The effect of gastro-intestinal mucus on drug absorption. Adv. Drug Deliv. Rev. 1993, 11, 201–220. [Google Scholar] [CrossRef]
- Ramaker, K.; Bade, S.; Rockendorf, N.; Meckelein, B.; Vollmer, E.; Schultz, H.; Froschle, G.W.; Frey, A. Absence of the Epithelial Glycocalyx As Potential Tumor Marker for the Early Detection of Colorectal Cancer. PLoS ONE 2016, 11, e0168801. [Google Scholar] [CrossRef]
- Hagerbaumer, P.; Vieth, M.; Anders, M.; Schumacher, U. Lectin Histochemistry Shows WGA, PHA-L and HPA Binding Increases During Progression of Human Colorectal Cancer. Anticancer Res. 2015, 35, 5333–5339. [Google Scholar]
- Benias, P.C.; Wells, R.G.; Sackey-Aboagye, B.; Klavan, H.; Reidy, J.; Buonocore, D.; Miranda, M.; Kornacki, S.; Wayne, M.; Carr-Locke, D.L.; et al. Structure and Distribution of an Unrecognized Interstitium in Human Tissues. Sci. Rep. 2018, 8, 4947. [Google Scholar] [CrossRef]
- Rastogi, V.; Puri, N.; Arora, S.; Kaur, G.; Yadav, L.; Sharma, R. Artefacts: A diagnostic dilemma—A review. J. Clin. Diagn. Res. 2013, 7, 2408–2413. [Google Scholar] [CrossRef]
- Potter, R.F.; Groom, A.C. Capillary diameter and geometry in cardiac and skeletal muscle studied by means of corrosion casts. Microvasc. Res. 1983, 25, 68–84. [Google Scholar] [CrossRef]
- Chevalier, L.; Selim, J.; Genty, D.; Baste, J.M.; Piton, N.; Boukhalfa, I.; Hamzaoui, M.; Pareige, P.; Richard, V. Electron microscopy approach for the visualization of the epithelial and endothelial glycocalyx. Morphologie 2017, 101, 55–63. [Google Scholar] [CrossRef]
- McDonald, D.M.; Baluk, P. Significance of blood vessel leakiness in cancer. Cancer Res. 2002, 62, 5381–5385. [Google Scholar]
- Wisse, E.; Braet, F.; Duimel, H.; Vreuls, C.; Koek, G.; Damink, S.W.O.; van den Broek, M.A.; De Geest, B.; Dejong, C.H.; Tateno, C.; et al. Fixation methods for electron microscopy of human and other liver. World J. Gastroenterol. 2010, 16, 2851–2866. [Google Scholar] [CrossRef]
- Miettinen, M.; Holthofer, H.; Lehto, V.P.; Miettinen, A.; Virtanen, I. Ulex europaeus I lectin as a marker for tumors derived from endothelial cells. Am. J. Clin. Pathol. 1983, 79, 32–36. [Google Scholar] [CrossRef]
- Hollingsworth, M.A.; Swanson, B.J. Mucins in cancer: Protection and control of the cell surface. Nat. Rev. Cancer 2004, 4, 45–60. [Google Scholar] [CrossRef]
- Bendris, N.; Stearns, C.J.; Reis, C.R.; Rodriguez-Canales, J.; Liu, H.; Witkiewicz, A.W.; Schmid, S.L. Sorting nexin 9 negatively regulates invadopodia formation and function in cancer cells. J. Cell Sci. 2016, 129, 2804–2816. [Google Scholar] [CrossRef] [Green Version]
- Bendris, N.; Williams, K.C.; Reis, C.R.; Welf, E.S.; Chen, P.H.; Lemmers, B.; Hahne, M.; Leong, H.S.; Schmid, S.L. SNX9 promotes metastasis by enhancing cancer cell invasion via differential regulation of RhoGTPases. Mol. Biol. Cell 2016, 27, 1409–1419. [Google Scholar] [CrossRef]
- Tanigawa, K.; Maekawa, M.; Kiyoi, T.; Nakayama, J.; Kitazawa, R.; Kitazawa, S.; Semba, K.; Taguchi, T.; Akita, S.; Yoshida, M.; et al. SNX9 determines the surface levels of integrin beta1 in vascular endothelial cells: Implication in poor prognosis of human colorectal cancers overexpressing SNX9. J. Cell. Physiol. 2019, 234, 17280–17294. [Google Scholar] [CrossRef]
- Moser, A.R.; Luongo, C.; Gould, K.A.; McNeley, M.K.; Shoemaker, A.R.; Dove, W.F. ApcMin: A mouse model for intestinal and mammary tumorigenesis. Eur. J. Cancer 1995, 31, 1061–1064. [Google Scholar] [CrossRef]
- Wagner, M.; Wiig, H. Tumor Interstitial Fluid Formation, Characterization, and Clinical Implications. Front. Oncol. 2015, 5, 115. [Google Scholar] [CrossRef] [Green Version]







© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tachi, M.; Okada, H.; Matsuhashi, N.; Takemura, G.; Suzuki, K.; Fukuda, H.; Niwa, A.; Tanaka, T.; Mori, H.; Hara, A.; et al. Human Colorectal Cancer Infrastructure Constructed by the Glycocalyx. J. Clin. Med. 2019, 8, 1270. https://doi.org/10.3390/jcm8091270
Tachi M, Okada H, Matsuhashi N, Takemura G, Suzuki K, Fukuda H, Niwa A, Tanaka T, Mori H, Hara A, et al. Human Colorectal Cancer Infrastructure Constructed by the Glycocalyx. Journal of Clinical Medicine. 2019; 8(9):1270. https://doi.org/10.3390/jcm8091270
Chicago/Turabian StyleTachi, Masahito, Hideshi Okada, Nobuhisa Matsuhashi, Genzou Takemura, Kodai Suzuki, Hirotsugu Fukuda, Ayumi Niwa, Takuji Tanaka, Hideki Mori, Akira Hara, and et al. 2019. "Human Colorectal Cancer Infrastructure Constructed by the Glycocalyx" Journal of Clinical Medicine 8, no. 9: 1270. https://doi.org/10.3390/jcm8091270
APA StyleTachi, M., Okada, H., Matsuhashi, N., Takemura, G., Suzuki, K., Fukuda, H., Niwa, A., Tanaka, T., Mori, H., Hara, A., Yoshida, K., Ogura, S., & Tomita, H. (2019). Human Colorectal Cancer Infrastructure Constructed by the Glycocalyx. Journal of Clinical Medicine, 8(9), 1270. https://doi.org/10.3390/jcm8091270