DJ-1 in Parkinson’s Disease: Clinical Insights and Therapeutic Perspectives

Abstract
:1. Introduction
- DJ-1 is highly expressed in cells with high energy demands, hence cells with higher levels of reactive oxygen species. Indeed, the majority of human cancers overexpress DJ-1 mRNA [28] and it is highly expressed in testicles, all of which are high-energy demand tissues characterized by a high glycolytic flux. Indeed, decreased expression of DJ-1 in sperm and epididymides correlates with male infertility [29,30]. DJ-1 protein levels also increase in human islets of Langerhans beta cells upon exposure to high glucose concentrations, indicating a protective role for DJ-1 in this cell type [31,32].
- DJ-1 is involved in protection from oxidative stress, although the molecular mechanisms underlying these effects are not entirely clear. DJ-1 overexpression blocks oxidative damage, while oxidative stress-induced cell death increases in the absence of DJ-1 in cell culture and animal models [33,34,35,36,37,38]. Unfortunately, the molecular mechanisms underlying DJ-1 function remain elusive and a key outstanding question is how DJ-1 function is affected by its oxidative modification. Several review articles summarize what is currently known regarding DJ-1 protection from oxidative stress [39,40,41].
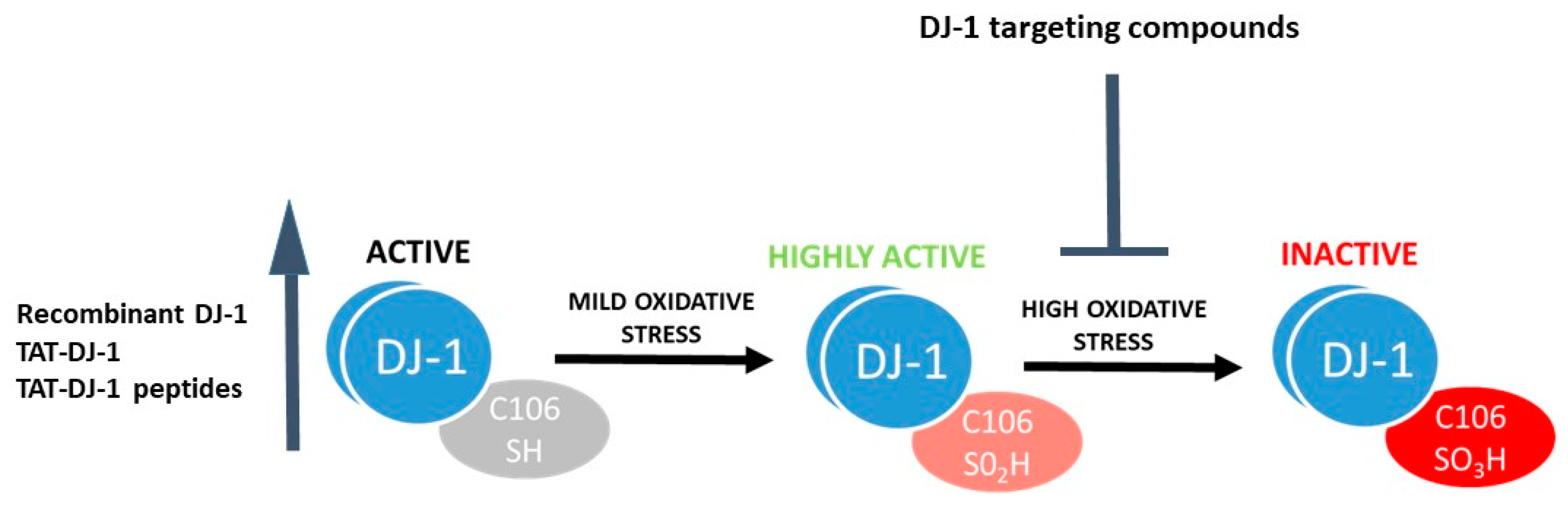
- DJ-1 can sense oxidative stress. The protein structure of DJ-1 has been studied in detail [42,43,44], with a particular emphasis on the relevance of the conserved Cysteine (Cys) residue at position 106 for biological function. Cys106 is the preferential target for oxidative protein modification and is required for DJ-1 mediated protection from oxidative stress [45,46,47]. The reduced form of DJ-1 (DJ-1 Cys106-SH) can be oxidized to a sulfinic acid form (DJ-1 Cys106-SO2H) and a sulfonic acid form (DJ-1 Cys106-SO3H) in the presence of moderate or high oxidative stress paradigms (Figure 1). While the reduced and sulfinic DJ-1 forms are stable, the sulfonic form of DJ-1 is unstable and prone to aggregate formation [47,48,49].
2. DJ-1 in the Pathological Human Brain
3. DJ-1 as a Biomarker
4. DJ-1 as a Target for Therapeutic Approaches
5. Conclusions and Future Directions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Bonifati, V.; Rizzu, P.; van Baren, M.J.; Schaap, O.; Breedveld, G.J.; Krieger, E.; Dekker, M.C.; Squitieri, F.; Ibanez, P.; Joosse, M.; et al. Mutations in the DJ-1 gene associated with autosomal recessive early-onset parkinsonism. Science 2003, 299, 256–259. [Google Scholar] [CrossRef] [PubMed]
- Abou-Sleiman, P.M.; Healy, D.G.; Quinn, N.; Lees, A.J.; Wood, N.W. The role of pathogenic DJ-1 mutations in Parkinson’s disease. Ann. Neurol. 2003, 54, 283–286. [Google Scholar] [CrossRef] [PubMed]
- Annesi, G.; Savettieri, G.; Pugliese, P.; D’Amelio, M.; Tarantino, P.; Ragonese, P.; La Bella, V.; Piccoli, T.; Civitelli, D.; Annesi, F.; et al. DJ-1 mutations and parkinsonism-dementia-amyotrophic lateral sclerosis complex. Ann. Neurol. 2005, 58, 803–807. [Google Scholar] [CrossRef] [PubMed]
- Tarantino, P.; Civitelli, D.; Annesi, F.; De Marco, E.V.; Rocca, F.E.; Pugliese, P.; Nicoletti, G.; Carrideo, S.; Provenzano, G.; Annesi, G.; et al. Compound heterozygosity in DJ-1 gene non-coding portion related to parkinsonism. Parkinsonism Relat. Disord. 2009, 15, 324–326. [Google Scholar] [CrossRef] [PubMed]
- Guo, J.F.; Xiao, B.; Liao, B.; Zhang, X.W.; Nie, L.L.; Zhang, Y.H.; Shen, L.; Jiang, H.; Xia, K.; Pan, Q.; et al. Mutation analysis of Parkin, PINK1, DJ-1 and ATP13A2 genes in Chinese patients with autosomal recessive early-onset Parkinsonism. Mov. Disord. 2008, 23, 2074–2079. [Google Scholar] [CrossRef] [PubMed]
- Hague, S.; Rogaeva, E.; Hernandez, D.; Gulick, C.; Singleton, A.; Hanson, M.; Johnson, J.; Weiser, R.; Gallardo, M.; Ravina, B.; et al. Early-onset Parkinson’s disease caused by a compound heterozygous DJ-1 mutation. Ann. Neurol. 2003, 54, 271–274. [Google Scholar] [CrossRef]
- Taghavi, S.; Chaouni, R.; Tafakhori, A.; Azcona, L.J.; Firouzabadi, S.G.; Omrani, M.D.; Jamshidi, J.; Emamalizadeh, B.; Shahidi, G.A.; Ahmadi, M.; et al. A Clinical and Molecular Genetic Study of 50 Families with Autosomal Recessive Parkinsonism Revealed Known and Novel Gene Mutations. Mol. Neurobiol. 2018, 55, 3477–3489. [Google Scholar] [CrossRef]
- Stephenson, S.E.; Djaldetti, R.; Rafehi, H.; Wilson, G.R.; Gillies, G.; Bahlo, M.; Lockhart, P.J. Familial early onset Parkinson’s disease caused by a homozygous frameshift variant in PARK7: Clinical features and literature update. Parkinsonism Relat. Disord. 2019. [Google Scholar] [CrossRef]
- Ghazavi, F.; Fazlali, Z.; Banihosseini, S.S.; Hosseini, S.R.; Kazemi, M.H.; Shojaee, S.; Parsa, K.; Sadeghi, H.; Sina, F.; Rohani, M.; et al. PRKN, DJ-1, and PINK1 screening identifies novel splice site mutation in PRKN and two novel DJ-1 mutations. Mov. Disord. 2011, 26, 80–89. [Google Scholar] [CrossRef]
- Hanagasi, H.A.; Giri, A.; Kartal, E.; Guven, G.; Bilgic, B.; Hauser, A.K.; Emre, M.; Heutink, P.; Basak, N.; Gasser, T.; et al. A novel homozygous DJ1 mutation causes parkinsonism and ALS in a Turkish family. Parkinsonism Relat. Disord. 2016, 29, 117–120. [Google Scholar] [CrossRef]
- Hering, R.; Strauss, K.M.; Tao, X.; Bauer, A.; Woitalla, D.; Mietz, E.M.; Petrovic, S.; Bauer, P.; Schaible, W.; Muller, T.; et al. Novel homozygous p.E64D mutation in DJ1 in early onset Parkinson disease (PARK7). Hum. Mutat. 2004, 24, 321–329. [Google Scholar] [CrossRef] [PubMed]
- Darvish, H.; Movafagh, A.; Omrani, M.D.; Firouzabadi, S.G.; Azargashb, E.; Jamshidi, J.; Khaligh, A.; Haghnejad, L.; Naeini, N.S.; Talebi, A.; et al. Detection of copy number changes in genes associated with Parkinson’s disease in Iranian patients. Neurosci. Lett. 2013, 551, 75–78. [Google Scholar] [CrossRef] [PubMed]
- Abbas, M.M.; Govindappa, S.T.; Sudhaman, S.; Thelma, B.K.; Juyal, R.C.; Behari, M.; Muthane, U.B. Early Onset Parkinson’s disease due to DJ1 mutations: An Indian study. Parkinsonism Relat. Disord. 2016, 32, 20–24. [Google Scholar] [CrossRef] [PubMed]
- Erer, S.; Egeli, U.; Zarifoglu, M.; Tezcan, G.; Cecener, G.; Tunca, B.; Ak, S.; Demirdogen, E.; Kenangil, G.; Kaleagasi, H.; et al. Mutation analysis of the PARKIN, PINK1, DJ1, and SNCA genes in Turkish early-onset Parkinson’s patients and genotype-phenotype correlations. Clin. Neurol. Neurosurg. 2016, 148, 147–153. [Google Scholar] [CrossRef] [PubMed]
- Di Nottia, M.; Masciullo, M.; Verrigni, D.; Petrillo, S.; Modoni, A.; Rizzo, V.; Di Giuda, D.; Rizza, T.; Niceta, M.; Torraco, A.; et al. DJ-1 modulates mitochondrial response to oxidative stress: Clues from a novel diagnosis of PARK7. Clin. Genet. 2017, 92, 18–25. [Google Scholar] [CrossRef] [PubMed]
- Macedo, M.G.; Verbaan, D.; Fang, Y.; van Rooden, S.M.; Visser, M.; Anar, B.; Uras, A.; Groen, J.L.; Rizzu, P.; van Hilten, J.J.; et al. Genotypic and phenotypic characteristics of Dutch patients with early onset Parkinson’s disease. Mov. Disord. 2009, 24, 196–203. [Google Scholar] [CrossRef]
- Taipa, R.; Pereira, C.; Reis, I.; Alonso, I.; Bastos-Lima, A.; Melo-Pires, M.; Magalhaes, M. DJ-1 linked parkinsonism (PARK7) is associated with Lewy body pathology. Brain 2016, 139, 1680–1687. [Google Scholar] [CrossRef]
- Wilson, M.A.; Collins, J.L.; Hod, Y.; Ringe, D.; Petsko, G.A. The 1.1-A resolution crystal structure of DJ-1, the protein mutated in autosomal recessive early onset Parkinson’s disease. Proc. Natl. Acad. Sci. USA 2003, 100, 9256–9261. [Google Scholar] [CrossRef]
- Gorner, K.; Holtorf, E.; Waak, J.; Pham, T.T.; Vogt-Weisenhorn, D.M.; Wurst, W.; Haass, C.; Kahle, P.J. Structural determinants of the C-terminal helix-kink-helix motif essential for protein stability and survival promoting activity of DJ-1. J. Biol. Chem. 2007, 282, 13680–13691. [Google Scholar] [CrossRef]
- Ramsey, C.P.; Giasson, B.I. L10p and P158DEL DJ-1 mutations cause protein instability, aggregation, and dimerization impairments. J. Neurosci. Res. 2010, 88, 3111–3124. [Google Scholar] [CrossRef] [Green Version]
- Malgieri, G.; Eliezer, D. Structural effects of Parkinson’s disease linked DJ-1 mutations. Protein Sci. 2008, 17, 855–868. [Google Scholar] [CrossRef] [PubMed]
- Kasten, M.; Hartmann, C.; Hampf, J.; Schaake, S.; Westenberger, A.; Vollstedt, E.J.; Balck, A.; Domingo, A.; Vulinovic, F.; Dulovic, M.; et al. Genotype-Phenotype Relations for the Parkinson’s Disease Genes Parkin, PINK1, DJ1: MDSGene Systematic Review. Mov. Disord. 2018, 33, 730–741. [Google Scholar] [CrossRef] [PubMed]
- Goldberg, M.S.; Pisani, A.; Haburcak, M.; Vortherms, T.A.; Kitada, T.; Costa, C.; Tong, Y.; Martella, G.; Tscherter, A.; Martins, A.; et al. Nigrostriatal dopaminergic deficits and hypokinesia caused by inactivation of the familial Parkinsonism-linked gene DJ-1. Neuron 2005, 45, 489–496. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Chandran, J.S.; Cai, H.; Mattson, M.P. DJ-1 is essential for long-term depression at hippocampal CA1 synapses. Neuromolecular Med. 2008, 10, 40–45. [Google Scholar] [CrossRef] [PubMed]
- Mihoub, M.; Abdallah, J.; Richarme, G. Protein Repair from Glycation by Glyoxals by the DJ-1 Family Maillard Deglycases. Adv. Exp. Med. Biol. 2017, 1037, 133–147. [Google Scholar] [CrossRef] [PubMed]
- Raninga, P.V.; Di Trapani, G.; Tonissen, K.F. The Multifaceted Roles of DJ-1 as an Antioxidant. Adv. Exp. Med. Biol. 2017, 1037, 67–87. [Google Scholar] [CrossRef] [PubMed]
- Takahashi-Niki, K.; Niki, T.; Iguchi-Ariga, S.M.M.; Ariga, H. Transcriptional Regulation of DJ-1. Adv. Exp. Med. Biol. 2017, 1037, 89–95. [Google Scholar] [CrossRef]
- Kawate, T.; Tsuchiya, B.; Iwaya, K. Expression of DJ-1 in Cancer Cells: Its Correlation with Clinical Significance. Adv. Exp. Med. Biol. 2017, 1037, 45–59. [Google Scholar] [CrossRef]
- Chunna, A.; Pu, X.P. Role of DJ-1 in Fertilization. Adv. Exp. Med. Biol. 2017, 1037, 61–66. [Google Scholar] [CrossRef]
- Wagenfeld, A.; Yeung, C.H.; Strupat, K.; Cooper, T.G. Shedding of a rat epididymal sperm protein associated with infertility induced by ornidazole and alpha-chlorohydrin. Biol. Reprod. 1998, 58, 1257–1265. [Google Scholar] [CrossRef]
- Eberhard, D.; Lammert, E. The Role of the Antioxidant Protein DJ-1 in Type 2 Diabetes Mellitus. Adv. Exp. Med. Biol. 2017, 1037, 173–186. [Google Scholar] [CrossRef] [PubMed]
- Jain, D.; Jain, R.; Eberhard, D.; Eglinger, J.; Bugliani, M.; Piemonti, L.; Marchetti, P.; Lammert, E. Age- and diet-dependent requirement of DJ-1 for glucose homeostasis in mice with implications for human type 2 diabetes. J. Mol. Cell. Biol. 2012, 4, 221–230. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Batelli, S.; Invernizzi, R.W.; Negro, A.; Calcagno, E.; Rodilossi, S.; Forloni, G.; Albani, D. The Parkinson’s disease-related protein DJ-1 protects dopaminergic neurons in vivo and cultured cells from alpha-synuclein and 6-hydroxydopamine toxicity. Neurodegener. Dis. 2015, 15, 13–23. [Google Scholar] [CrossRef] [PubMed]
- Kim, R.H.; Smith, P.D.; Aleyasin, H.; Hayley, S.; Mount, M.P.; Pownall, S.; Wakeham, A.; You-Ten, A.J.; Kalia, S.K.; Horne, P.; et al. Hypersensitivity of DJ-1-deficient mice to 1-methyl-4-phenyl-1,2,3,6-tetrahydropyrindine (MPTP) and oxidative stress. Proc. Natl. Acad. Sci. USA 2005, 102, 5215–5220. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meulener, M.; Whitworth, A.J.; Armstrong-Gold, C.E.; Rizzu, P.; Heutink, P.; Wes, P.D.; Pallanck, L.J.; Bonini, N.M. Drosophila DJ-1 mutants are selectively sensitive to environmental toxins associated with Parkinson’s disease. Curr. Biol. 2005, 15, 1572–1577. [Google Scholar] [CrossRef] [PubMed]
- Ottolini, D.; Cali, T.; Negro, A.; Brini, M. The Parkinson disease-related protein DJ-1 counteracts mitochondrial impairment induced by the tumour suppressor protein p53 by enhancing endoplasmic reticulum-mitochondria tethering. Hum. Mol. Genet. 2013, 22, 2152–2168. [Google Scholar] [CrossRef] [PubMed]
- Taira, T.; Saito, Y.; Niki, T.; Iguchi-Ariga, S.M.; Takahashi, K.; Ariga, H. DJ-1 has a role in antioxidative stress to prevent cell death. EMBO Rep. 2004, 5, 213–218. [Google Scholar] [CrossRef] [PubMed]
- Thomas, K.J.; McCoy, M.K.; Blackinton, J.; Beilina, A.; van der Brug, M.; Sandebring, A.; Miller, D.; Maric, D.; Cedazo-Minguez, A.; Cookson, M.R. DJ-1 acts in parallel to the PINK1/parkin pathway to control mitochondrial function and autophagy. Hum. Mol. Genet. 2011, 20, 40–50. [Google Scholar] [CrossRef] [PubMed]
- Ariga, H.; Takahashi-Niki, K.; Kato, I.; Maita, H.; Niki, T.; Iguchi-Ariga, S.M. Neuroprotective function of DJ-1 in Parkinson’s disease. Oxid. Med. Cell Longev. 2013, 2013, 683920. [Google Scholar] [CrossRef]
- Biosa, A.; Sandrelli, F.; Beltramini, M.; Greggio, E.; Bubacco, L.; Bisaglia, M. Recent findings on the physiological function of DJ-1: Beyond Parkinson’s disease. Neurobiol. Dis. 2017, 108, 65–72. [Google Scholar] [CrossRef]
- Cookson, M.R. Unravelling the role of defective genes. Prog. Brain Res. 2010, 183, 43–57. [Google Scholar] [CrossRef] [PubMed]
- Blackinton, J.; Lakshminarasimhan, M.; Thomas, K.J.; Ahmad, R.; Greggio, E.; Raza, A.S.; Cookson, M.R.; Wilson, M.A. Formation of a stabilized cysteine sulfinic acid is critical for the mitochondrial function of the parkinsonism protein DJ-1. J. Biol. Chem. 2009, 284, 6476–6485. [Google Scholar] [CrossRef] [PubMed]
- Canet-Aviles, R.M.; Wilson, M.A.; Miller, D.W.; Ahmad, R.; McLendon, C.; Bandyopadhyay, S.; Baptista, M.J.; Ringe, D.; Petsko, G.A.; Cookson, M.R. The Parkinson’s disease protein DJ-1 is neuroprotective due to cysteine-sulfinic acid-driven mitochondrial localization. Proc. Natl. Acad. Sci. USA 2004, 101, 9103–9108. [Google Scholar] [CrossRef] [PubMed]
- Wilson, M.A.; St Amour, C.V.; Collins, J.L.; Ringe, D.; Petsko, G.A. The 1.8-A resolution crystal structure of YDR533Cp from Saccharomyces cerevisiae: A member of the DJ-1/ThiJ/PfpI superfamily. Proc. Natl. Acad. Sci. USA 2004, 101, 1531–1536. [Google Scholar] [CrossRef] [PubMed]
- Kinumi, T.; Kimata, J.; Taira, T.; Ariga, H.; Niki, E. Cysteine-106 of DJ-1 is the most sensitive cysteine residue to hydrogen peroxide-mediated oxidation in vivo in human umbilical vein endothelial cells. Biochem. Biophys. Res. Commun. 2004, 317, 722–728. [Google Scholar] [CrossRef] [PubMed]
- Smith, N.; Wilson, M.A. Structural Biology of the DJ-1 Superfamily. Adv. Exp. Med. Biol. 2017, 1037, 5–24. [Google Scholar] [CrossRef] [PubMed]
- Wilson, M.A. The role of cysteine oxidation in DJ-1 function and dysfunction. Antioxid. Redox Signal. 2011, 15, 111–122. [Google Scholar] [CrossRef] [PubMed]
- Catazaro, J.; Andrews, T.; Milkovic, N.M.; Lin, J.; Lowe, A.J.; Wilson, M.A.; Powers, R. (15)N CEST data and traditional model-free analysis capture fast internal dynamics of DJ-1. Anal. Biochem. 2018, 542, 24–28. [Google Scholar] [CrossRef] [PubMed]
- Kiss, R.; Zhu, M.; Jojart, B.; Czajlik, A.; Solti, K.; Forizs, B.; Nagy, E.; Zsila, F.; Beke-Somfai, T.; Toth, G. Structural features of human DJ-1 in distinct Cys106 oxidative states and their relevance to its loss of function in disease. Biochim. Biophys. Acta Gen. Subj. 2017, 1861, 2619–2629. [Google Scholar] [CrossRef] [Green Version]
- Bandopadhyay, R.; Kingsbury, A.E.; Cookson, M.R.; Reid, A.R.; Evans, I.M.; Hope, A.D.; Pittman, A.M.; Lashley, T.; Canet-Aviles, R.; Miller, D.W.; et al. The expression of DJ-1 (PARK7) in normal human CNS and idiopathic Parkinson’s disease. Brain 2004, 127, 420–430. [Google Scholar] [CrossRef]
- Kumaran, R.; Vandrovcova, J.; Luk, C.; Sharma, S.; Renton, A.; Wood, N.W.; Hardy, J.A.; Lees, A.J.; Bandopadhyay, R. Differential DJ-1 gene expression in Parkinson’s disease. Neurobiol. Dis. 2009, 36, 393–400. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.; Sullards, M.C.; Olzmann, J.A.; Rees, H.D.; Weintraub, S.T.; Bostwick, D.E.; Gearing, M.; Levey, A.I.; Chin, L.S.; Li, L. Oxidative damage of DJ-1 is linked to sporadic Parkinson and Alzheimer diseases. J. Biol. Chem. 2006, 281, 10816–10824. [Google Scholar] [CrossRef] [PubMed]
- Mullett, S.J.; Hamilton, R.L.; Hinkle, D.A. DJ-1 immunoreactivity in human brain astrocytes is dependent on infarct presence and infarct age. Neuropathology 2009, 29, 125–131. [Google Scholar] [CrossRef] [PubMed]
- Saito, Y. Oxidized DJ-1 as a possible biomarker of Parkinson’s disease. J. Clin. Biochem. Nutr. 2014, 54, 138–144. [Google Scholar] [CrossRef] [PubMed]
- Saito, Y.; Hamakubo, T.; Yoshida, Y.; Ogawa, Y.; Hara, Y.; Fujimura, H.; Imai, Y.; Iwanari, H.; Mochizuki, Y.; Shichiri, M.; et al. Preparation and application of monoclonal antibodies against oxidized DJ-1. Significant elevation of oxidized DJ-1 in erythrocytes of early-stage Parkinson disease patients. Neurosci. Lett. 2009, 465, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Rizzu, P.; Hinkle, D.A.; Zhukareva, V.; Bonifati, V.; Severijnen, L.A.; Martinez, D.; Ravid, R.; Kamphorst, W.; Eberwine, J.H.; Lee, V.M.; et al. DJ-1 colocalizes with tau inclusions: A link between parkinsonism and dementia. Ann. Neurol. 2004, 55, 113–118. [Google Scholar] [CrossRef] [PubMed]
- Neumann, M.; Muller, V.; Gorner, K.; Kretzschmar, H.A.; Haass, C.; Kahle, P.J. Pathological properties of the Parkinson’s disease-associated protein DJ-1 in alpha-synucleinopathies and tauopathies: Relevance for multiple system atrophy and Pick’s disease. Acta Neuropathol. 2004, 107, 489–496. [Google Scholar] [CrossRef]
- Batelli, S.; Albani, D.; Rametta, R.; Polito, L.; Prato, F.; Pesaresi, M.; Negro, A.; Forloni, G. DJ-1 modulates alpha-synuclein aggregation state in a cellular model of oxidative stress: Relevance for Parkinson’s disease and involvement of HSP70. PLoS ONE 2008, 3, e1884. [Google Scholar] [CrossRef]
- Zondler, L.; Miller-Fleming, L.; Repici, M.; Goncalves, S.; Tenreiro, S.; Rosado-Ramos, R.; Betzer, C.; Straatman, K.R.; Jensen, P.H.; Giorgini, F.; et al. DJ-1 interactions with alpha-synuclein attenuate aggregation and cellular toxicity in models of Parkinson’s disease. Cell Death Dis. 2014, 5, e1350. [Google Scholar] [CrossRef]
- Sajjad, M.U.; Green, E.W.; Miller-Fleming, L.; Hands, S.; Herrera, F.; Campesan, S.; Khoshnan, A.; Outeiro, T.F.; Giorgini, F.; Wyttenbach, A. DJ-1 modulates aggregation and pathogenesis in models of Huntington’s disease. Hum. Mol. Genet. 2014, 23, 755–766. [Google Scholar] [CrossRef]
- Wang, Z.; Zhang, Y.; Zhang, S.; Guo, Q.; Tan, Y.; Wang, X.; Xiong, R.; Ding, J.; Chen, S. DJ-1 can inhibit microtubule associated protein 1 B formed aggregates. Mol. Neurodegener. 2011, 6, 38. [Google Scholar] [CrossRef] [PubMed]
- Dos Santos, M.C.T.; Scheller, D.; Schulte, C.; Mesa, I.R.; Colman, P.; Bujac, S.R.; Bell, R.; Berteau, C.; Perez, L.T.; Lachmann, I.; et al. Evaluation of cerebrospinal fluid proteins as potential biomarkers for early stage Parkinson’s disease diagnosis. PLoS ONE 2018, 13, e0206536. [Google Scholar] [CrossRef] [PubMed]
- Farotti, L.; Paciotti, S.; Tasegian, A.; Eusebi, P.; Parnetti, L. Discovery, validation and optimization of cerebrospinal fluid biomarkers for use in Parkinson’s disease. Expert Rev. Mol. Diagn. 2017, 17, 771–780. [Google Scholar] [CrossRef] [PubMed]
- Hong, Z.; Shi, M.; Chung, K.A.; Quinn, J.F.; Peskind, E.R.; Galasko, D.; Jankovic, J.; Zabetian, C.P.; Leverenz, J.B.; Baird, G.; et al. DJ-1 and alpha-synuclein in human cerebrospinal fluid as biomarkers of Parkinson’s disease. Brain 2010, 133, 713–726. [Google Scholar] [CrossRef] [PubMed]
- Salvesen, L.; Bech, S.; Lokkegaard, A.; Hjermind, L.E.; Nielsen, J.E.; Pakkenberg, B.; Tanassi, J.T.; Heegaard, N.H.; Winge, K. The DJ-1 concentration in cerebrospinal fluid does not differentiate among Parkinsonian syndromes. Parkinsonism Relat. Disord. 2012, 18, 899–901. [Google Scholar] [CrossRef] [PubMed]
- Shi, M.; Zabetian, C.P.; Hancock, A.M.; Ginghina, C.; Hong, Z.; Yearout, D.; Chung, K.A.; Quinn, J.F.; Peskind, E.R.; Galasko, D.; et al. Significance and confounders of peripheral DJ-1 and alpha-synuclein in Parkinson’s disease. Neurosci. Lett. 2010, 480, 78–82. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Z.H.; Chen, Z.T.; Zhou, R.L.; Zhang, X.; Ye, Q.Y.; Wang, Y.Z. Increased DJ-1 and alpha-Synuclein in Plasma Neural-Derived Exosomes as Potential Markers for Parkinson’s Disease. Front. Aging Neurosci. 2018, 10, 438. [Google Scholar] [CrossRef] [PubMed]
- Shi, M.; Kovac, A.; Korff, A.; Cook, T.J.; Ginghina, C.; Bullock, K.M.; Yang, L.; Stewart, T.; Zheng, D.; Aro, P.; et al. CNS tau efflux via exosomes is likely increased in Parkinson’s disease but not in Alzheimer’s disease. Alzheimers Dement. 2016, 12, 1125–1131. [Google Scholar] [CrossRef] [PubMed]
- Yamagishi, Y.; Saigoh, K.; Saito, Y.; Ogawa, I.; Mitsui, Y.; Hamada, Y.; Samukawa, M.; Suzuki, H.; Kuwahara, M.; Hirano, M.; et al. Diagnosis of Parkinson’s disease and the level of oxidized DJ-1 protein. Neurosci. Res. 2018, 128, 58–62. [Google Scholar] [CrossRef] [PubMed]
- Jang, J.; Jeong, S.; Lee, S.I.; Seol, W.; Seo, H.; Son, I.; Ho, D.H. Oxidized DJ-1 Levels in Urine Samples as a Putative Biomarker for Parkinson’s Disease. Parkinsons Dis. 2018, 2018, 1241757. [Google Scholar] [CrossRef] [PubMed]
- Drechsel, J.; Mandl, F.A.; Sieber, S.A. Chemical Probe To Monitor the Parkinsonism-Associated Protein DJ-1 in Live Cells. ACS Chem. Biol. 2018, 13, 2016–2019. [Google Scholar] [CrossRef] [PubMed]
- Gao, H.; Yang, W.; Qi, Z.; Lu, L.; Duan, C.; Zhao, C.; Yang, H. DJ-1 protects dopaminergic neurons against rotenone-induced apoptosis by enhancing ERK-dependent mitophagy. J. Mol. Biol. 2012, 423, 232–248. [Google Scholar] [CrossRef] [PubMed]
- Inden, M.; Taira, T.; Kitamura, Y.; Yanagida, T.; Tsuchiya, D.; Takata, K.; Yanagisawa, D.; Nishimura, K.; Taniguchi, T.; Kiso, Y.; et al. PARK7 DJ-1 protects against degeneration of nigral dopaminergic neurons in Parkinson’s disease rat model. Neurobiol. Dis. 2006, 24, 144–158. [Google Scholar] [CrossRef] [PubMed]
- Sun, S.Y.; An, C.N.; Pu, X.P. DJ-1 protein protects dopaminergic neurons against 6-OHDA/MG-132-induced neurotoxicity in rats. Brain Res. Bull. 2012, 88, 609–616. [Google Scholar] [CrossRef] [PubMed]
- Antoniou, X.; Borsello, T. Cell Permeable Peptides: A Promising Tool to Deliver Neuroprotective Agents in the Brain. Pharmaceuticals 2010, 3, 379–392. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lev, N.; Barhum, Y.; Ben-Zur, T.; Aharony, I.; Trifonov, L.; Regev, N.; Melamed, E.; Gruzman, A.; Offen, D. A DJ-1 Based Peptide Attenuates Dopaminergic Degeneration in Mice Models of Parkinson’s Disease via Enhancing Nrf2. PLoS ONE 2015, 10, e0127549. [Google Scholar] [CrossRef]
- Glat, M.J.; Ben-Zur, T.; Barhum, Y.; Offen, D. Neuroprotective Effect of a DJ-1 Based Peptide in a Toxin Induced Mouse Model of Multiple System Atrophy. PLoS ONE 2016, 11, e0148170. [Google Scholar] [CrossRef] [PubMed]
- Molcho, L.; Ben-Zur, T.; Barhum, Y.; Offen, D. DJ-1 based peptide, ND-13, promote functional recovery in mouse model of focal ischemic injury. PLoS ONE 2018, 13, e0192954. [Google Scholar] [CrossRef]
- Kim, W.; Kim, D.W.; Jeong, H.J.; Yoo, D.Y.; Jung, H.Y.; Nam, S.M.; Kim, J.H.; Choi, J.H.; Won, M.H.; Yoon, Y.S.; et al. Tat-DJ-1 protects neurons from ischemic damage in the ventral horn of rabbit spinal cord via increasing antioxidant levels. Neurochem. Res. 2014, 39, 187–193. [Google Scholar] [CrossRef]
- Lin, J.; Prahlad, J.; Wilson, M.A. Conservation of oxidative protein stabilization in an insect homologue of parkinsonism-associated protein DJ-1. Biochemistry 2012, 51, 3799–3807. [Google Scholar] [CrossRef]
- Miyazaki, S.; Yanagida, T.; Nunome, K.; Ishikawa, S.; Inden, M.; Kitamura, Y.; Nakagawa, S.; Taira, T.; Hirota, K.; Niwa, M.; et al. DJ-1-binding compounds prevent oxidative stress-induced cell death and movement defect in Parkinson’s disease model rats. J. Neurochem. 2008, 105, 2418–2434. [Google Scholar] [CrossRef] [PubMed]
- Inden, M.; Kitamura, Y.; Takahashi, K.; Takata, K.; Ito, N.; Niwa, R.; Funayama, R.; Nishimura, K.; Taniguchi, T.; Honda, T.; et al. Protection against dopaminergic neurodegeneration in Parkinson’s disease-model animals by a modulator of the oxidized form of DJ-1, a wild-type of familial Parkinson’s disease-linked PARK7. J. Pharmacol. Sci. 2011, 117, 189–203. [Google Scholar] [CrossRef] [PubMed]
- Kitamura, Y.; Watanabe, S.; Taguchi, M.; Takagi, K.; Kawata, T.; Takahashi-Niki, K.; Yasui, H.; Maita, H.; Iguchi-Ariga, S.M.; Ariga, H. Neuroprotective effect of a new DJ-1-binding compound against neurodegeneration in Parkinson’s disease and stroke model rats. Mol. Neurodegener. 2011, 6, 48. [Google Scholar] [CrossRef] [PubMed]
- Yamane, K.; Kitamura, Y.; Yanagida, T.; Takata, K.; Yanagisawa, D.; Taniguchi, T.; Taira, T.; Ariga, H. Oxidative neurodegeneration is prevented by UCP0045037, an allosteric modulator for the reduced form of DJ-1, a wild-type of familial Parkinson’s disease-linked PARK7. Int. J. Mol. Sci. 2009, 10, 4789–4804. [Google Scholar] [CrossRef] [PubMed]
- Yanagida, T.; Kitamura, Y.; Yamane, K.; Takahashi, K.; Takata, K.; Yanagisawa, D.; Yasui, H.; Taniguchi, T.; Taira, T.; Honda, T.; et al. Protection against oxidative stress-induced neurodegeneration by a modulator for DJ-1, the wild-type of familial Parkinson’s disease-linked PARK7. J. Pharmacol. Sci. 2009, 109, 463–468. [Google Scholar] [CrossRef] [PubMed]
- Takahashi-Niki, K.; Inafune, A.; Michitani, N.; Hatakeyama, Y.; Suzuki, K.; Sasaki, M.; Kitamura, Y.; Niki, T.; Iguchi-Ariga, S.M.; Ariga, H. DJ-1-dependent protective activity of DJ-1-binding compound no. 23 against neuronal cell death in MPTP-treated mouse model of Parkinson’s disease. J. Pharmacol. Sci. 2015, 127, 305–310. [Google Scholar] [CrossRef] [PubMed]
- Kitamura, Y.; Inden, M.; Kimoto, Y.; Takata, K.; Yanagisawa, D.; Hijioka, M.; Ashihara, E.; Tooyama, I.; Shimohama, S.; Ariga, H. Effects of a DJ-1-Binding Compound on Spatial Learning and Memory Impairment in a Mouse Model of Alzheimer’s Disease. J. Alzheimers Dis. 2017, 55, 67–72. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.; Oh, S.T.; Jeong, H.J.; Pak, S.C.; Park, H.-J.; Kim, J.; Cho, H.-S.; Jeon, S. MPTP-induced vulnerability of dopamine neurons in A53T a-synuclein overexpressed mice with the potential involvement of DJ-1 downregulation. Korean J. Physiol. Pharmacol. 2017, 21, 625–632. [Google Scholar] [CrossRef]


{kind=link}
| PARK7 Mutation | Location | References |
|---|---|---|
| g.159C > G + IVS4 + 3insA: Compound het | 5′ UTR; Intron 4 | [4] |
| Ex1-5del (g.4443-18524del): Hom | Exons 1–5 | [1] |
| Leu10Pro: Hom | Exon 2 | [5] |
| Thr19Lysfs*5 + IVS6-1G→C: Compound het | Exon 2; Intron 6 | [6] |
| Asp24Metfs*3: Hom | Exon 2 | [7] |
| Met26Ile: Hom | Exon 2 | [2] |
| Ile31Aspfs*2: Hom | Exon 2 | [8] |
| g.11032A>G: Hom | Intron 2 | [9] |
| Gln45*: Hom | Exon 3 | [10] |
| Glu64Asp: Hom | Exon 3 | [11] |
| Ex5del: Hom (breakpoints not detailed) | Exon 5 | [12] |
| Ile105Phe: Hom | Exon 5 | [13] |
| Ala107Pro: Hom | Exon 5 | [9] |
| g.16677 A > C: Hom | Intron 5 | [14] |
| Thr154Lys: Hom | Exon 7 | [15] |
| Pro158del: Hom | Exon 7 | [16] |
| Glu163Lys: Hom | Exon 7 | [3] |
| Leu166Pro: Hom | Exon 7 | [1] |
| Leu172Gln: Hom | Exon 7 | [17] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Repici, M.; Giorgini, F. DJ-1 in Parkinson’s Disease: Clinical Insights and Therapeutic Perspectives. J. Clin. Med. 2019, 8, 1377. https://doi.org/10.3390/jcm8091377
Repici M, Giorgini F. DJ-1 in Parkinson’s Disease: Clinical Insights and Therapeutic Perspectives. Journal of Clinical Medicine. 2019; 8(9):1377. https://doi.org/10.3390/jcm8091377
Chicago/Turabian StyleRepici, Mariaelena, and Flaviano Giorgini. 2019. "DJ-1 in Parkinson’s Disease: Clinical Insights and Therapeutic Perspectives" Journal of Clinical Medicine 8, no. 9: 1377. https://doi.org/10.3390/jcm8091377
APA StyleRepici, M., & Giorgini, F. (2019). DJ-1 in Parkinson’s Disease: Clinical Insights and Therapeutic Perspectives. Journal of Clinical Medicine, 8(9), 1377. https://doi.org/10.3390/jcm8091377