Whole Genome Sequencing Results Associated with Minimum Inhibitory Concentrations of 14 Anti-Tuberculosis Drugs among Rifampicin-Resistant Isolates of Mycobacterium Tuberculosis from Iran

 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Design and Mycobacterial Strains
2.2. Broth Microdilution
2.3. Whole Genome Sequencing and Bioinformatic Analysis
2.4. Statistical Analysis
3. Results
3.1. Mycobacterial Strains
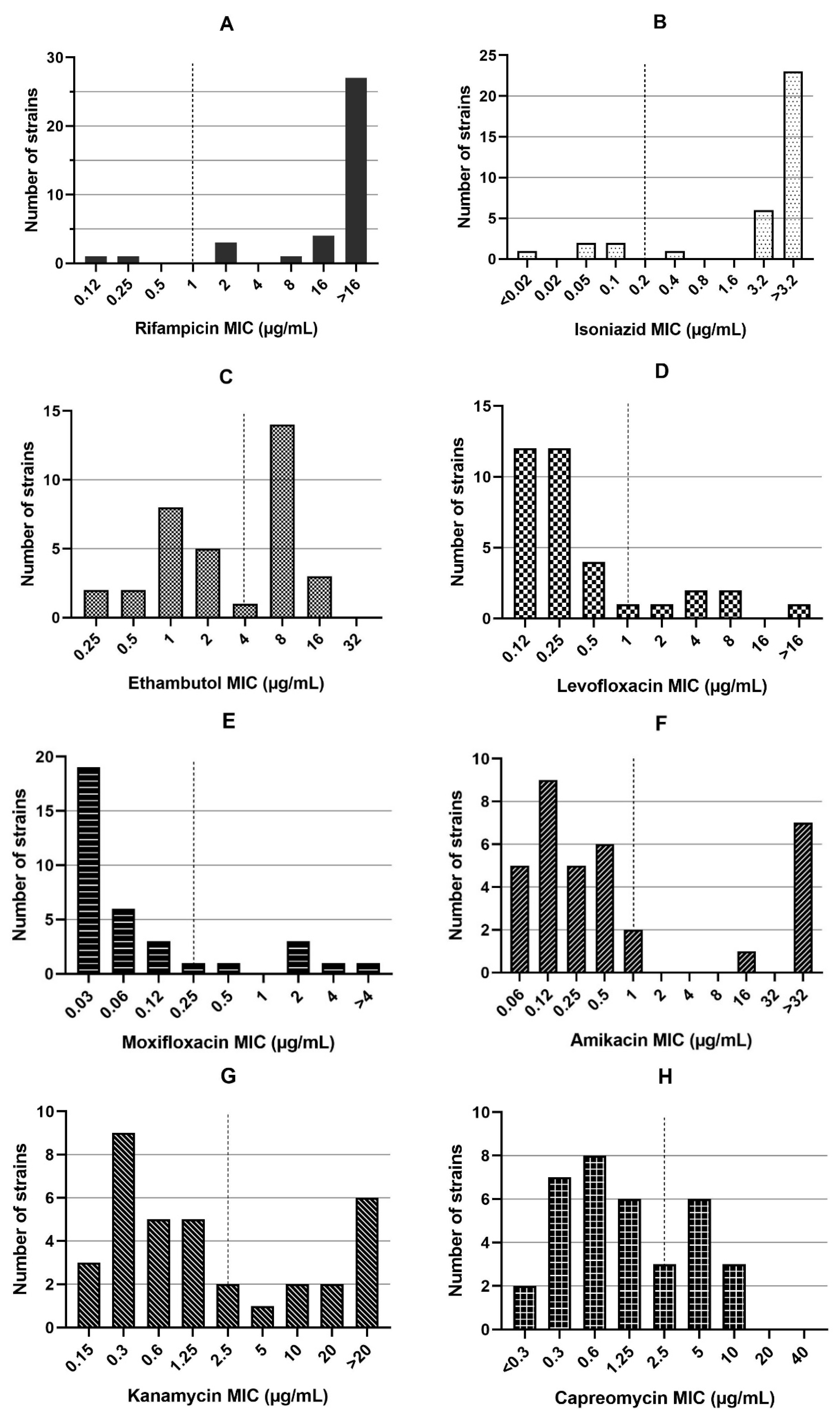
3.2. Minimum Inhibitory Concentrations
3.3. Resistance-Conferring Mutations Using WGS
3.3.1. Genotypic Resistance to the First-Line Drugs
3.3.2. Genotypic Resistance to Fluoroquinolones
3.3.3. Genotypic Resistance to Second-Line Injectable Drugs
3.3.4. Genotypic Resistance to Other Second-Line Drugs (WHO Group C)
3.3.5. Genotypic Resistance to Add-On Agents
3.4. Sensitivity, Specificity, and Accuracy of WGS
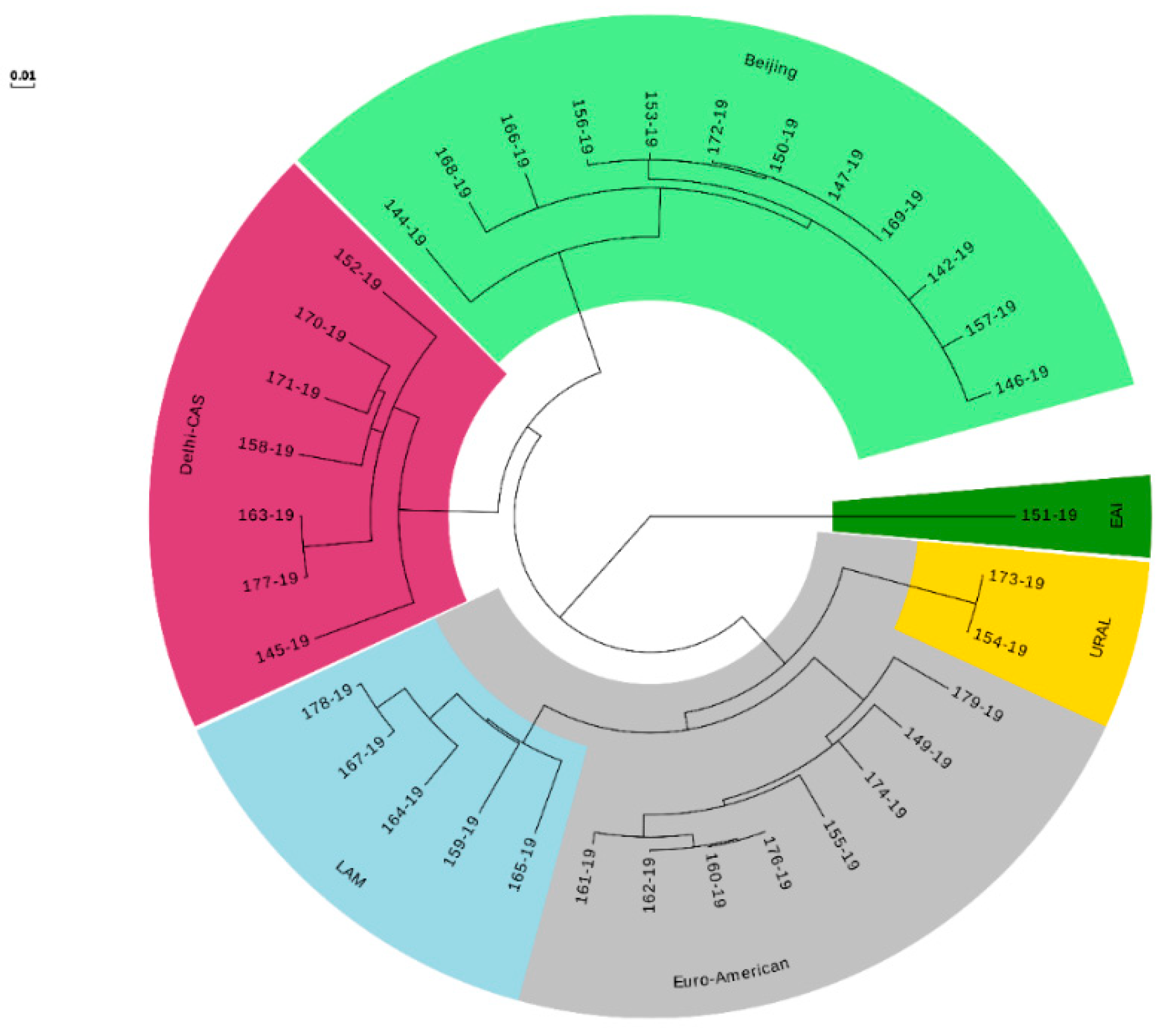
3.5. WGS-Based MTB Genotyping
4. Discussion
5. Conclusions
Author Contributions
Acknowledgments
Conflicts of Interest
References
- WHO. Global Tuberculosis Report 2017; World Health Organization: Geneva, Switzerland, 2017. [Google Scholar]
- WHO. Global Tuberculosis Report 2018; World Health Organization: Geneva, Switzerland, 2018. [Google Scholar]
- Coll, F.; Phelan, J.; Hill-Cawthorne, G.A.; Nair, M.B.; Mallard, K.; Ali, S.; Abdallah, A.M.; Alghamdi, S.; Alsomali, M.; Ahmed, A.; et al. Genome-wide analysis of multi-and extensively drug-resistant Mycobacterium tuberculosis. Nat. Genet. 2018, 50, 307. [Google Scholar] [CrossRef] [Green Version]
- Kaniga, K.; Cirillo, D.M.; Hoffner, S.; Ismail, N.A.; Kaur, D.; Lounis, N.; Metchock, B.; Pfyffer, G.E.; Venter, A. A multilaboratory, multicountry study to determine MIC quality control ranges for phenotypic drug susceptibility testing of selected first-line antituberculosis drugs, second-line injectables, fluoroquinolones, clofazimine, and linezolid. J. Clin. Microbiol. 2016, 54, 2963–2968. [Google Scholar] [CrossRef] [Green Version]
- Black, P.A.; Warren, R.M.; Louw, G.E.; van Helden, P.D.; Victor, T.C.; Kana, B.D. Energy metabolism and drug efflux in Mycobacterium tuberculosis. Antimicrob. Agents Chemother. 2014, 58, 2491–2503. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rastogi, N.; Goh, K.; David, H. Drug susceptibility testing in tuberculosis: A comparison of the proportion methods using Lowenstein-Jensen, Middlebrook 7H10 and 7H11 agar media and a radiometric method. Res. Microbiol. 1989, 140, 405–417. [Google Scholar] [CrossRef]
- Victor, T.C.; Warren, R.; Butt, J.L.; Jordaan, A.M.; Felix, J.V.; Venter, A.; Sirgel, F.A.; Schaaf, H.S.; Donald, P.R.; Richardson, M.; et al. Genome and MIC stability in Mycobacterium tuberculosis and indications for continuation of use of isoniazid in multidrug-resistant tuberculosis. J. Med. Microbiol. 1997, 46, 847–857. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coll, F.; McNerney, R.; Preston, M.D.; Guerra-Assunção, J.A.; Warry, A.; Hill-Cawthorne, G.; Mallard, K.; Nair, M.; Miranda, A.; Alves, A.; et al. Rapid determination of anti-tuberculosis drug resistance from whole-genome sequences. Genome Med. 2015, 7, 51. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deggim-Messmer, V.; Bloemberg, G.V.; Ritter, C.; Voit, A.; Hömke, R.; Keller, P.M.; Böttger, E.C. Diagnostic molecular mycobacteriology in regions with low tuberculosis endemicity: Combining real-time PCR assays for detection of multiple mycobacterial pathogens with line probe assays for identification of resistance mutations. EBioMedicine 2016, 9, 228–237. [Google Scholar] [CrossRef] [Green Version]
- Engström, A. Fighting an old disease with modern tools: Characteristics and molecular detection methods of drug-resistant Mycobacterium tuberculosis. Infect. Dis. 2016, 48, 1–17. [Google Scholar] [CrossRef]
- Gygli, S.M.; Keller, P.M.; Ballif, M.; Blöchliger, N.; Hömke, R.; Reinhard, M.; Loiseau, C.; Ritter, C.; Sander, P.; Borrell, S.; et al. Whole-genome sequencing for drug resistance profile prediction in Mycobacterium tuberculosis. Antimicrob. Agents Chemother. 2019, 63, e02175-18. [Google Scholar] [CrossRef] [Green Version]
- Dookie, N.; Rambaran, S.; Padayatchi, N.; Mahomed, S.; Naidoo, K. Evolution of drug resistance in Mycobacterium tuberculosis: A review on the molecular determinants of resistance and implications for personalized care. J. Antimicrob. Chemother. 2018, 73, 1138–1151. [Google Scholar] [CrossRef] [Green Version]
- Leao, S.C.; Martin, A.; Mejia, G.I.; Palomino, J.C.; Robledo, J.; da Silva Telles, M.A.; Portaels, F. Practical Handbook for the Phenotypic and Genotypic Identification of Mycobacteria; Vanden Broele: Brugges, Belgium, 2004. [Google Scholar]
- Canetti, G.; Froman, S.; Grosset, J.; Hauduroy, P.; Langerová, M.; Mahler, H.T.; Meissner, G.; Mitchison, D.A.; Šula, L. Mycobacteria: Laboratory methods for testing drug sensitivity and resistance. Bull. World Health Organ. 1963, 29, 565. [Google Scholar] [PubMed]
- Rancoita, P.M.V.; Cugnata, F.; Gibertoni Cruz, A.L.; Borroni, E.; Hoosdally, S.J.; Walker, T.M.; Grazian, C.; Davies, T.J.; Peto, T.E.A.; Crook, D.W.; et al. Validating a 14-drug microtiter plate containing bedaquiline and delamanid for large-scale research susceptibility testing of Mycobacterium tuberculosis. Antimicrob. Agents Chemother. 2018, 62, e00344-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- WHO. Technical Report on Critical Concentrations for Drug Susceptibility Testing of Medicines Used in the Treatment of Drug-Resistant Tuberculosis; World Health Organization: Geneva, Switzerland, 2018. [Google Scholar]
- Rationale Documents for EUCAST Clinical Breakpoints. 2019. Available online: http://www.eucast.org/documents/rd/ (accessed on 17 October 2019).
- Kohl, T.A.; Utpatel, C.; Schleusener, V.; De Filippo, M.R.; Beckert, P.; Cirillo, D.M.; Niemann, S. MTBseq: A comprehensive pipeline for whole genome sequence analysis of Mycobacterium tuberculosis complex isolates. PeerJ 2018, 6, e5895. [Google Scholar] [CrossRef] [Green Version]
- Comas, I.; Chakravartti, J.; Small, P.M.; Galagan, J.; Niemann, S.; Kremer, K.; Ernst, J.D.; Gagneux, S. Human T cell epitopes of Mycobacterium tuberculosis are evolutionarily hyperconserved. Nat. Genet. 2010, 42, 498. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walker, T.M.; Ip, C.L.; Harrell, R.H.; Evans, J.T.; Kapatai, G.; Dedicoat, M.J.; Eyre, D.W.; Wilson, D.J.; Hawkey, P.M.; Crook, D.W.; et al. Whole-genome sequencing to delineate Mycobacterium tuberculosis outbreaks: A retrospective observational study. Lancet Infect. Dis. 2013, 13, 137–146. [Google Scholar] [CrossRef] [Green Version]
- Feuerriegel, S.; Köser, C.U.; Niemann, S. Phylogenetic polymorphisms in antibiotic resistance genes of the Mycobacterium tuberculosis complex. J. Antimicrob. Chemother. 2014, 69, 1205–1210. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coll, F.; McNerney, R.; Guerra-Assunção, J.A.; Glynn, J.R.; Perdigão, J.; Viveiros, M.; Portugal, I.; Pain, A.; Martin, N.; Clark, T.G. A robust SNP barcode for typing Mycobacterium tuberculosis complex strains. Nat. Commun. 2014, 5, 4812. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Homolka, S.; Projahn, M.; Feuerriegel, S.; Ubben, T.; Diel, R.; Nübel, U.; Niemann, S. High resolution discrimination of clinical Mycobacterium tuberculosis complex strains based on single nucleotide polymorphisms. PLoS ONE 2012, 7, e39855. [Google Scholar] [CrossRef] [Green Version]
- Merker, M.; Blin, C.; Mona, S.; Duforet-Frebourg, N.; Lecher, S.; Willery, E.; Blum, M.G.; Rüsch-Gerdes, S.; Mokrousov, I.; Aleksic, E.; et al. Evolutionary history and global spread of the Mycobacterium tuberculosis Beijing lineage. Nat. Genet. 2015, 47, 242. [Google Scholar] [CrossRef]
- Stamatakis, A. RAxML version 8: A tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef]
- Walker, T.M.; Kohl, T.A.; Omar, S.V.; Hedge, J.; Del Ojo Elias, C.; Bradley, P.; Iqbal, Z.; Feuerriegel, S.; Niehaus, K.E.; Wilson, D.J.; et al. Whole-genome sequencing for prediction of Mycobacterium tuberculosis drug susceptibility and resistance: A retrospective cohort study. Lancet Infect. Dis. 2015, 15, 1193–1202. [Google Scholar] [CrossRef] [Green Version]
- Miotto, P.; Tessema, B.; Tagliani, E.; Chindelevitch, L.; Starks, A.M.; Emerson, C.; Hanna, D.; Kim, P.S.; Liwski, R.; Zignol, M.; et al. A standardised method for interpreting the association between mutations and phenotypic drug resistance in Mycobacterium tuberculosis. Eur. Respir. J. 2017, 50, 1701354. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- CRyPTIC Consortium and the 100,000 Genome Project. Prediction of susceptibility to first-line tuberculosis drugs by DNA sequencing. N. Engl. J. Med. 2018, 379, 1403–1415. [Google Scholar] [CrossRef] [PubMed]
- De Vos, M. Identification of Mechanisms Regulating the Intra Cellular Concentration of Rifampicin in Mycobacterium tuberculosis; Stellenbosch University: Stellenbosch, South Africa, 2013. [Google Scholar]
- Campbell, P.J.; Morlock, G.P.; Sikes, R.D.; Dalton, T.L.; Metchock, B.; Starks, A.M.; Hooks, D.P.; Cowan, L.S.; Plikaytis, B.B.; Posey, J.E. Molecular detection of mutations associated with first-and second-line drug resistance compared with conventional drug susceptibility testing of Mycobacterium tuberculosis. Antimicrob. Agents Chemother. 2011, 55, 2032–2041. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Desjardins, C.A.; Cohen, K.A.; Munsamy, V.; Abeel, T.; Maharaj, K.; Walker, B.J.; Shea, T.P.; Almeida, D.V.; Manson, A.L.; Salazar, A.; et al. Genomic and functional analyses of Mycobacterium tuberculosis strains implicate ald in D-cycloserine resistance. Nat. Genet. 2016, 48, 544. [Google Scholar] [CrossRef] [Green Version]
- Ismail, N.; Omar, S.V.; Ismail, N.A.; Petersa, R.P.H. Collated data of mutation frequencies and associated genetic variants of bedaquiline, clofazimine and linezolid resistance in Mycobacterium tuberculosis. Data Brief 2018, 20, 1975. [Google Scholar] [CrossRef]
- Pang, Y.; Zong, Z.; Huo, F.; Jing, W.; Ma, Y.; Dong, L.; Li, Y.; Zhao, L.; Fu, Y.; Huang, H. In Vitro drug susceptibility of bedaquiline, delamanid, linezolid, clofazimine, moxifloxacin, and gatifloxacin against extensively drug-resistant tuberculosis in Beijing, China. Antimicrob. Agents Chemother. 2017, 61, e00900-17. [Google Scholar] [CrossRef] [Green Version]
- EUCAST. Rationale for Defining a Reference Method for the Determination of Minimum Inhibitory Concentrations (MIC) of Anti-Tuberculous Agents for Mycobacterium tuberculosis Complex; EUCAST: Basel, Switzerland, 2019. [Google Scholar]
- Kardan-Yamchi, J.; Kazemian, H.; Haeili, M.; Harati, A.A.; Amini, S.; Feizabadi, M.M. Expression analysis of 10 efflux pump genes in multidrug-resistant and extensively drug-resistant Mycobacterium tuberculosis clinical isolates. J. Glob. Antimicrob. Resist. 2019, 17, 201–208. [Google Scholar] [CrossRef]
- Jamieson, F.B.; Guthrie, J.L.; Neemuchwala, A.; Lastovetska, O.; Melano, R.G.; Mehaffy, C. Profiling of rpoB mutations and MICs for rifampin and rifabutin in Mycobacterium tuberculosis. J. Clin. Microbiol. 2014, 52, 2157–2162. [Google Scholar] [CrossRef] [Green Version]
- Ma, X.; Wang, H.; Deng, Y.; Liu, Z.; Xu, Y.; Pan, X.; Musser, J.M.; Graviss, E.A. rpoB gene mutations and molecular characterization of rifampin-resistant Mycobacterium tuberculosis isolates from Shandong Province, China. J. Clin. Microbiol. 2006, 44, 3409–3412. [Google Scholar] [CrossRef] [Green Version]
- Mvelase, N.R.; Pillay, M.; Sibanda, W.; Ngozo, J.N.; Brust, J.C.M.; Mlisana, K.P. rpoB Mutations Causing Discordant Rifampicin Susceptibility in Mycobacterium tuberculosis: Retrospective Analysis of Prevalence, Phenotypic, Genotypic, and Treatment Outcomes. Open Forum Infect. Dis. 2019, 16, ofz065. [Google Scholar] [CrossRef] [PubMed]
- Mohajeri, P.; Sadri, H.; Farahani, A.; Norozi, B.; Atashi, S. Frequency of mutations associated with rifampicin resistance in Mycobacterium tuberculosis strains isolated from patients in West of Iran. Microb. Drug Resist. 2015, 21, 315–319. [Google Scholar] [CrossRef]
- Lavender, C.; Globan, M.; Sievers, A.; Billman-Jacobe, H.; Fyfe, J. Molecular characterization of isoniazid-resistant Mycobacterium tuberculosis isolates collected in Australia. Antimicrob. Agents Chemother. 2005, 49, 4068–4074. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huyen, M.N.; Cobelens, F.G.; Buu, T.N.; Lan, N.T.; Dung, N.H.; Kremer, K.; Tiemersma, E.W.; van Soolingen, D. Epidemiology of isoniazid resistance mutations and their effect on tuberculosis treatment outcomes. Antimicrob. Agents Chemother. 2013, 57, 3620–3627. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seifert, M.; Catanzaro, D.; Catanzaro, A.; Rodwell, T.C. Genetic mutations associated with isoniazid resistance in Mycobacterium tuberculosis: A systematic review. PLoS ONE 2015, 10, e0119628. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jagielski, T.; Bakuła, Z.; Roeske, K.; Kamiński, M.; Napiórkowska, A.; Augustynowicz-Kopeć, E.; Zwolska, Z.; Bielecki, J. Detection of mutations associated with isoniazid resistance in multidrug-resistant Mycobacterium tuberculosis clinical isolates. J. Antimicrob. Chemother. 2014, 69, 2369–2375. [Google Scholar] [CrossRef] [Green Version]
- Ghodousi, A.; Tagliani, E.; Karunaratne, E.; Niemann, S.; Perera, J.; Köser, C.U.; Cirillo, D.M. Isoniazid Resistance in Mycobacterium tuberculosis Is a Heterogeneous Phenotype Composed of Overlapping MIC Distributions with Different Underlying Resistance Mechanisms. Antimicrob. Agents Chemother. 2019, 63, e00092-19. [Google Scholar] [CrossRef] [Green Version]
- Ando, H.; Miyoshi-Akiyama, T.; Watanabe, S.; Kirikae, T. A silent mutation in mabA confers isoniazid resistance on Mycobacterium tuberculosis. Mol. Microbiol. 2014, 91, 538–547. [Google Scholar] [CrossRef]
- Zhang, M.; Yue, J.; Yang, Y.P.; Zhang, H.M.; Lei, J.Q.; Jin, R.L.; Zhang, X.L.; Wang, H.H. Detection of mutations associated with isoniazid resistance in Mycobacterium tuberculosis isolates from China. J. Clin. Microbiol. 2005, 43, 5477–5482. [Google Scholar] [CrossRef] [Green Version]
- Khosravi, A.D.; Sirous, M.; Abdi, M.; Ahmadkhosravi, N. Characterization of the most common embCAB gene mutations associated with ethambutol resistance in Mycobacterium tuberculosis isolates from Iran. Infect. Drug Resist. 2019, 12, 579. [Google Scholar] [CrossRef] [Green Version]
- Sun, Q.; Xiao, T.Y.; Liu, H.C.; Zhao, X.Q.; Liu, Z.G.; Li, Y.N.; Zeng, H.; Zhao, L.L.; Wan, K.L. Mutations within embCAB are associated with variable level of ethambutol resistance in Mycobacterium tuberculosis isolates from China. Antimicrob. Agents Chemother. 2018, 62, e01279-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, J.; Gao, X.; Luo, T.; Wu, J.; Sun, G.; Liu, Q.; Jiang, Y.; Zhang, Y.; Mei, J.; Gao, Q. Association of gyrA/B mutations and resistance levels to fluoroquinolones in clinical isolates of Mycobacterium tuberculosis. Emerg. Microbes Infect. 2014, 3, 1–5. [Google Scholar]
- Da Silva, P.E.; Von Groll, A.; Martin, A.; Palomino, J.C. Efflux as a mechanism for drug resistance in Mycobacterium tuberculosis. FEMS Immunol. Med. Microbiol. 2011, 63, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brossier, F.; Pham, A.; Bernard, C.; Aubry, A.; Jarlier, V.; Veziris, N.; Sougakoff, W. Molecular investigation of resistance to second-line injectable drugs in multidrug-resistant clinical isolates of Mycobacterium tuberculosis in France. Antimicrob. Agents Chemother. 2017, 61, e01299-16. [Google Scholar] [PubMed] [Green Version]
- Gikalo, M.B.; Nosova, E.Y.; Krylova, L.Y.; Moroz, A.M. The role of eis mutations in the development of kanamycin resistance in Mycobacterium tuberculosis isolates from the Moscow region. J. Antimicrob. Chemother. 2012, 67, 2107–2109. [Google Scholar] [CrossRef] [PubMed]
- Georghiou, S.B.; Magana, M.; Garfein, R.S.; Catanzaro, D.G.; Catanzaro, A.; Rodwell, T.C. Evaluation of genetic mutations associated with Mycobacterium tuberculosis resistance to amikacin, kanamycin and capreomycin: A systematic review. PLoS ONE 2012, 7, e33275. [Google Scholar] [CrossRef]
- Pholwat, S.; Stroup, S.; Heysell, S.; Ogarkov, O.; Zhdanova, S.; Ramakrishnan, G.; Houpt, E. eis promoter C14G and C15G mutations do not confer kanamycin resistance in Mycobacterium tuberculosis. Antimicrob. Agents Chemother. 2016, 60, 7522–7523. [Google Scholar]
- Tan, Y.; Su, B.; Zheng, H.; Song, Y.; Wang, Y.; Pang, Y. Molecular characterization of prothionamide-resistant Mycobacterium tuberculosis isolates in southern China. Front. Microbiol. 2017, 8, 2358. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.; He, G.; Wang, S.; Lin, S.; Chen, J.; Zhang, W. Evaluation of whole-genome sequence method to diagnose resistance of 13 anti-tuberculosis drugs and characterize resistance genes in clinical multi-drug resistance Mycobacterium tuberculosis isolates from China. Front. Microbiol. 2019, 10, 1741. [Google Scholar] [CrossRef] [Green Version]
- Leung, K.L.; Yip, C.W.; Yeung, Y.L.; Wong, K.L.; Chan, W.Y.; Chan, M.Y.; Kam, K.M. Usefulness of resistant gene markers for predicting treatment outcome on second-line anti-tuberculosis drugs. J. Appl. Microbiol. 2010, 109, 2087–2094. [Google Scholar] [CrossRef]
- Chen, J.M.; Uplekar, S.; Gordon, S.V.; Cole, S.T. A point mutation in cycA partially contributes to the D-cycloserine resistance trait of Mycobacterium bovis BCG vaccine strains. PLoS ONE 2012, 7, e43467. [Google Scholar] [CrossRef]
- Chen, J.; Zhang, S.; Cui, P.; Shi, W.; Zhang, W.; Zhang, Y. Identification of novel mutations associated with cycloserine resistance in Mycobacterium tuberculosis. J. Antimicrob. Chemother. 2017, 72, 3272–3276. [Google Scholar] [CrossRef] [Green Version]
- Ismail, N.A.; Omar, S.V.; Joseph, L.; Govender, N.; Blows, L.; Ismail, F.; Koornhof, H.; Dreyer, A.W.; Kaniga, K.; Ndjeka, N. Defining bedaquiline susceptibility, resistance, cross-resistance and associated genetic determinants: A retrospective cohort study. EBioMedicine 2018, 28, 136–142. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, S.; Chen, J.; Cui, P.; Shi, W.; Zhang, W.; Zhang, Y. Identification of novel mutations associated with clofazimine resistance in Mycobacterium tuberculosis. J. Antimicrob. Chemother. 2015, 70, 2507–2510. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kazemian, H.; Haeili, M.; Kardan Yamchi, J.; Rezaei, F.; Gizaw Feyisa, S.; Zahednamazi, F.; Mohajeri, P.; Zaker Bostanabd, S.; Hashemi Shahraki, A.; Imani Fooladi, A.A.; et al. Antimycobacterial activity of linezolid against multidrug-resistant and extensively drug-resistant strains of Mycobacterium tuberculosis in Iran. Int. J. Antimicrob. Agents 2015, 45, 668. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.S.; Kim, K.J.; Choi, H.; Lee, S.H. Delamanid, bedaquiline, and linezolid minimum inhibitory concentration distributions and resistance-related gene mutations in multidrug-resistant and extensively drug-resistant tuberculosis in Korea. Ann. Lab. Med. 2018, 38, 563–568. [Google Scholar] [CrossRef] [PubMed]
- Ghodousi, A.; Rizvi, A.H.; Baloch, A.Q.; Ghafoor, A.; Khanzada, F.M.; Qadir, M.; Borroni, E.; Trovato, A.; Tahseen, S.; Cirillo, D.M. Acquisition of cross-resistance to bedaquiline and clofazimine following treatment for tuberculosis in Pakistan. Antimicrob. Agents Chemother. 2019, 63, e00915-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| No. of Strains | LFX | MFX | AMK | KAN | CAP | |
|---|---|---|---|---|---|---|
| Pre-XDR | 3 | R | R | S | S | S |
| 4 | S | S | R | R | R | |
| 2 | S | S | S | R | S | |
| 1 | S | S | S | R | R | |
| XDR | 3 | R | R | R | R | R |
| Drug | Resistance Phenotype (No. of Strains) | Resistance Genotype | MIC (µg/mL) | Total | ||
|---|---|---|---|---|---|---|
| Gene | Mutations | No. of Strains | ||||
| RIF | Resistant (35) | rpoB | Ser531Leu | 19 | >16 | 34/35 |
| Ser531Leu/Arg862Trp (freq: 19.9%) | 1 | >16 | ||||
| Ser531Leu/Val643Ala (freq: 38.5%) | 1 | >16 | ||||
| Ser531leu/Pro564Ser (freq: 12.6%) | 1 | >16 | ||||
| His526Tyr | 2 | >16 | ||||
| 2 | 16 | |||||
| His526Leu | 2 | 16 | ||||
| His526Asp/Ser512Thr (freq: 5.6%) | 1 | >16 | ||||
| Leu533Pro | 2 | 4 | ||||
| Gln513Leu | 1 | >16 | ||||
| Asp516Val | 1 | >16 | ||||
| Ile572Phe | 1 | 8 | ||||
| WT | 1 | 2 | ||||
| INH | Resistant (30) | katG | Ser315Thr | 11 | >3.2 | 29/30 |
| 2 | 3.2 | |||||
| 1353 to 1476-del | 1 | >3.2 | ||||
| Ser315Thr/Leu216Pro | 1 | 3.2 | ||||
| katG/ahpC-ups | Asn138His/T-76A | 1 | >3.2 | |||
| katG/ahpC-ups | Ser315Thr/T-77G | 1 | >3.2 | |||
| katG/ahpC-ups/fabG1-ups/inhA | Trp149Arg/G-51A/ −27 to −38 del/Ser94Ala | 1 | >3.2 | |||
| katG/ahpC-ups/inhA | Trp149Arg/G-51A/Ser94Ala | 1 | >3.2 | |||
| katG/fabG1-inhA | Met624Ile/C-15T | 1 | >3.2 | |||
| katG/fabG1-inhA | Ser315Thr/C-15T | 3 | >3.2 | |||
| katG/fabG1-inhA | Leu587Pro/C-15T | 1 | >3.2 | |||
| katG/fabG1-inhA | Trp191Arg, Ala706Thr/C-15T | 1 | >3.2 | |||
| inhA/fabG1-inhA | Ala239Val/C-15T | 1 | >3.2 | |||
| fabG1/fabG1-inhA | Leu203leu (freq: 57.5%)/C-15T (freq: 50.9%) | 1 | 3.2 | |||
| fabG1-inhA | C-15T | 1 | 3.2 | |||
| 1 | 0.4 | |||||
| katG, inhA, ahpC, fabG1, furA | WT | 1 | 3.2 | |||
| EMB | Resistant (17) | embB | Met306Val | 10 | 8 | 17/17 |
| 1 | 16 | |||||
| Gly406Ser | 1 | 16 | ||||
| Gln497Arg | 1 | 8 | ||||
| Met306Ile/Gly406Asp | 1 | 16 | ||||
| embB/embA-ups | Met306Ile/C-16A | 1 | 8 | |||
| embB/embA-ups | Met306Ile+Trp349Arg/C-16A | 1 | 8 | |||
| embB/embA-ups | Gln497Arg/C-12T | 1 | 8 | |||
| Susceptible (18) | embA-ups | C-16T | 1 | 2 | 3/18 | |
| embB | His1002Arg | 1 | 1 | |||
| Asp328Gly | 1 | 4 | ||||
| Drug | Resistance Phenotype (No. of Strains) | Resistance Genotype | MIC (µg/mL) | Total | ||
|---|---|---|---|---|---|---|
| Gene | Mutations | No. of Strains | ||||
| LFX | Resistant (6) | gyrA | Gly88Cys | 1 | >16 | 5/6 |
| 1 | 8 | |||||
| Asp94Gly | 2 | 4 | ||||
| Ser91Pro | 1 | 8 | ||||
| gyrA, gyrB | WT | 1 | 2 | |||
| Susceptible (29) | gyrA | Asp94Ala | 1 | 1 | 1/29 | |
| MFX | Resistant (6) | gyrA | Gly88Cys | 1 | >4 | 5/6 |
| 1 | 4 | |||||
| Asp94Gly | 2 | 2 | ||||
| Ser91Pro | 1 | 2 | ||||
| gyrA, gyrB | WT | 1 | 0.5 | |||
| Susceptible (29) | gyrA | Asp94Ala | 1 | 0.25 | 1/29 | |
| Drug | Resistance Genotype (No. of Strains) | Resistance Genotype | MIC (µg/mL) | Total | ||
|---|---|---|---|---|---|---|
| Gene | Mutations | No. of Strains | ||||
| AMK | Resistant (8) | rrs | A1401G | 6 | >32 | 8/8 |
| 1 | 16 | |||||
| G1484T | 1 | >32 | ||||
| KAN | Resistant (11) | rrs | A1401G | 5 | >20 | 11/11 |
| 2 | 20 | |||||
| G1484T | 1 | >20 | ||||
| eis-ups | G-10A | 1 | 10 | |||
| C-12T | 1 | 5 | ||||
| C-15G | 1 | 10 | ||||
| Susceptible (24) | eis/eis-ups | G334 ins/C-14T | 1 | 0.3 | 1/24 | |
| CAP | Resistant (9) | rrs | A1401G | 5 | 5 | 9/9 |
| 2 | 10 | |||||
| G1484T | 1 | 10 | ||||
| tlyA | C210 ins | 1 | 5 | |||
| Susceptible (26) | tlyA | Gly196Glu | 1 | 2.5 | 1/26 | |
| Drug | Resistance Phenotype (No. of Strains) | Resistance Genotype | MIC (µg/mL) | Total | ||
|---|---|---|---|---|---|---|
| Gene | Mutations | No. of Strains | ||||
| PTO | Resistant (19) | ethA | Thr314Ile | 1 | 5 | 19/19 |
| Ala341Glu | 1 | 5 | ||||
| Trp21 del (freq: 10.32%) | 1 | 5 | ||||
| T1361 del | 1 | 10 | ||||
| Gln254Pro | 1 | 10 | ||||
| 1 | 5 | |||||
| ethA/fabG1-ups/inhA | Trp116 del (freq: 59.63%)/−27 to −38 del/Ser94Ala (freq: 20.69%) | 1 | 20 | |||
| ethA-ups | T-11G | 1 | 10 | |||
| fabG1-inhA | C-15T | 2 | 20 | |||
| 3 | 10 | |||||
| 3 | 5 | |||||
| inhA/fabG1-inhA/ndh | Ala239Val/C-15T/399A ins | 1 | 20 | |||
| fabG1/fabG1-inhA | Leu203Leu (freq: 57.5%)/C-15T (freq: 50.9%) | 1 | 20 | |||
| inhA | Ser94Ala | 1 | 20 | |||
| Susceptible (16) | ethA | Gln254Pro | 2 | 0.6 | 2/16 | |
| DCS | Resistant (9) | cycA | Pro188Ala | 1 | 32 | 1/9 |
| ald, ddlA, alr, cycA | WT | 6 | 32 | |||
| WT | 2 | 64 | ||||
| Susceptible (26) | ald | GC432 ins | 2 | 16 | 10/26 | |
| Cyc333Trp (freq: 10.9%) | 1 | 16 | ||||
| Ala340Ser (freq: 5.0%) | 1 | 16 | ||||
| ddlA | 2A ins (freq: 7.3%) | 1 | 4 | |||
| Arg264Gly (freq: 6.5%)/Val260Gly (freq: 6.0%) | 1 | 16 | ||||
| Gln293Glu (freq: 5.0%) | 1 | 8 | ||||
| Thr322Ser (freq: 5.1%) | 1 | 2 | ||||
| alr | Leu113Arg | 1 | 16 | |||
| cycA | Leu21His (freq: 5.3%) | 1 | 8 | |||
| CFZ | Resistant (5) | Rv1979c | 1014A del | 1 | 2 | 1/5 |
| Rv0678, Rv1979c, pepQ | WT | 3 | 2 | |||
| WT | 1 | 4 | ||||
| Susceptible (30) | Rv0678 | Gly87Arg | 1 | 1 | 3/30 | |
| Rv1979c | Asn297Ser | 1 | 1 | |||
| pepQ | Ala87Gly (freq: 12.79%) | 1 | 1 | |||
| LZD | Susceptible (35) | rrl | C1537T | 1 | 0.25 | 4/35 |
| C1331T | 1 | 0.12 | ||||
| rplC | Ala72Thr | 2 | 0.5 | |||
| Drug | Sensitivity (%) | Specificity (%) | Accuracy (%) |
|---|---|---|---|
| Rifampicin | 97.1 | 100.0 | 97.3 |
| Isoniazid | 96.7 | 100.0 | 97.1 |
| Ethambutol | 100.0 | 83.3 | 91.4 |
| Levofloxacin | 83.3 | 96.5 | 94.3 |
| Moxifloxacin | 83.3 | 96.5 | 94.3 |
| Amikacin | 100.0 | 100.0 | 100.0 |
| Kanamycin | 100.0 | 95.8 | 97.1 |
| Capreomycin | 100.0 | 96.2 | 97.1 |
| Prothinamide | 100.0 | 87.5 | 94.3 |
| D-cycloserine | 11.1 | 61.5 | 48.6 |
| Clofazimine | 20.0 | 90.0 | 80.0 |
| Linezolid | - | 88.6 | 88.6 |
| Bedaquiline | 0.0 | 97.1 | 94.3 |
| Delamanid | 44.4 | 96.2 | 82.9 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kardan-Yamchi, J.; Kazemian, H.; Battaglia, S.; Abtahi, H.; Rahimi Foroushani, A.; Hamzelou, G.; Cirillo, D.M.; Ghodousi, A.; Feizabadi, M.M. Whole Genome Sequencing Results Associated with Minimum Inhibitory Concentrations of 14 Anti-Tuberculosis Drugs among Rifampicin-Resistant Isolates of Mycobacterium Tuberculosis from Iran. J. Clin. Med. 2020, 9, 465. https://doi.org/10.3390/jcm9020465
Kardan-Yamchi J, Kazemian H, Battaglia S, Abtahi H, Rahimi Foroushani A, Hamzelou G, Cirillo DM, Ghodousi A, Feizabadi MM. Whole Genome Sequencing Results Associated with Minimum Inhibitory Concentrations of 14 Anti-Tuberculosis Drugs among Rifampicin-Resistant Isolates of Mycobacterium Tuberculosis from Iran. Journal of Clinical Medicine. 2020; 9(2):465. https://doi.org/10.3390/jcm9020465
Chicago/Turabian StyleKardan-Yamchi, Jalil, Hossein Kazemian, Simone Battaglia, Hamidreza Abtahi, Abbas Rahimi Foroushani, Gholamreza Hamzelou, Daniela Maria Cirillo, Arash Ghodousi, and Mohammad Mehdi Feizabadi. 2020. "Whole Genome Sequencing Results Associated with Minimum Inhibitory Concentrations of 14 Anti-Tuberculosis Drugs among Rifampicin-Resistant Isolates of Mycobacterium Tuberculosis from Iran" Journal of Clinical Medicine 9, no. 2: 465. https://doi.org/10.3390/jcm9020465
APA StyleKardan-Yamchi, J., Kazemian, H., Battaglia, S., Abtahi, H., Rahimi Foroushani, A., Hamzelou, G., Cirillo, D. M., Ghodousi, A., & Feizabadi, M. M. (2020). Whole Genome Sequencing Results Associated with Minimum Inhibitory Concentrations of 14 Anti-Tuberculosis Drugs among Rifampicin-Resistant Isolates of Mycobacterium Tuberculosis from Iran. Journal of Clinical Medicine, 9(2), 465. https://doi.org/10.3390/jcm9020465