Human Wharton’s Jelly—Cellular Specificity, Stemness Potency, Animal Models, and Current Application in Human Clinical Trials

 ,
,  ,
,  ,
, 

Abstract
:1. Introduction
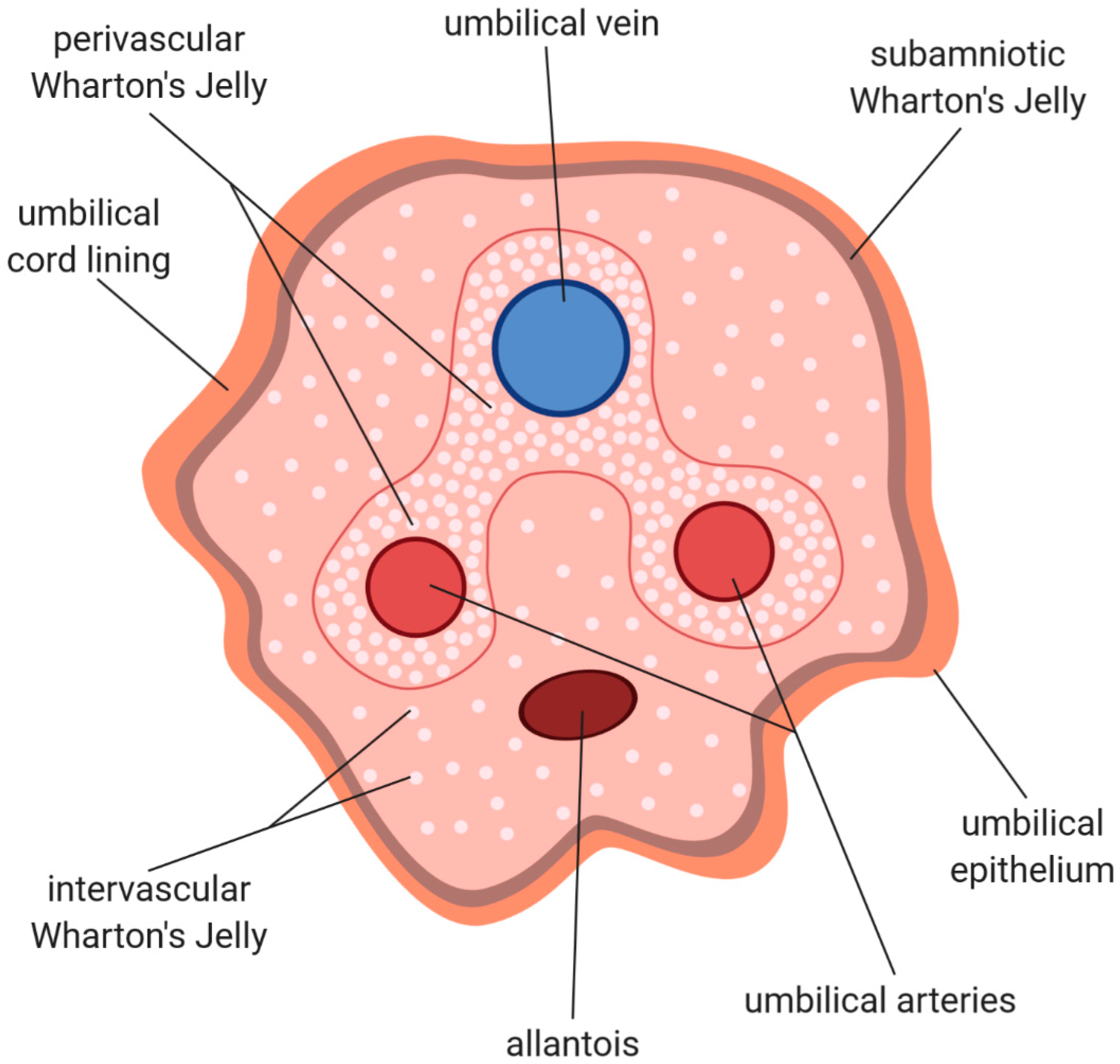
2. Histological Structure and Cellular Composition of Umbilical Cord
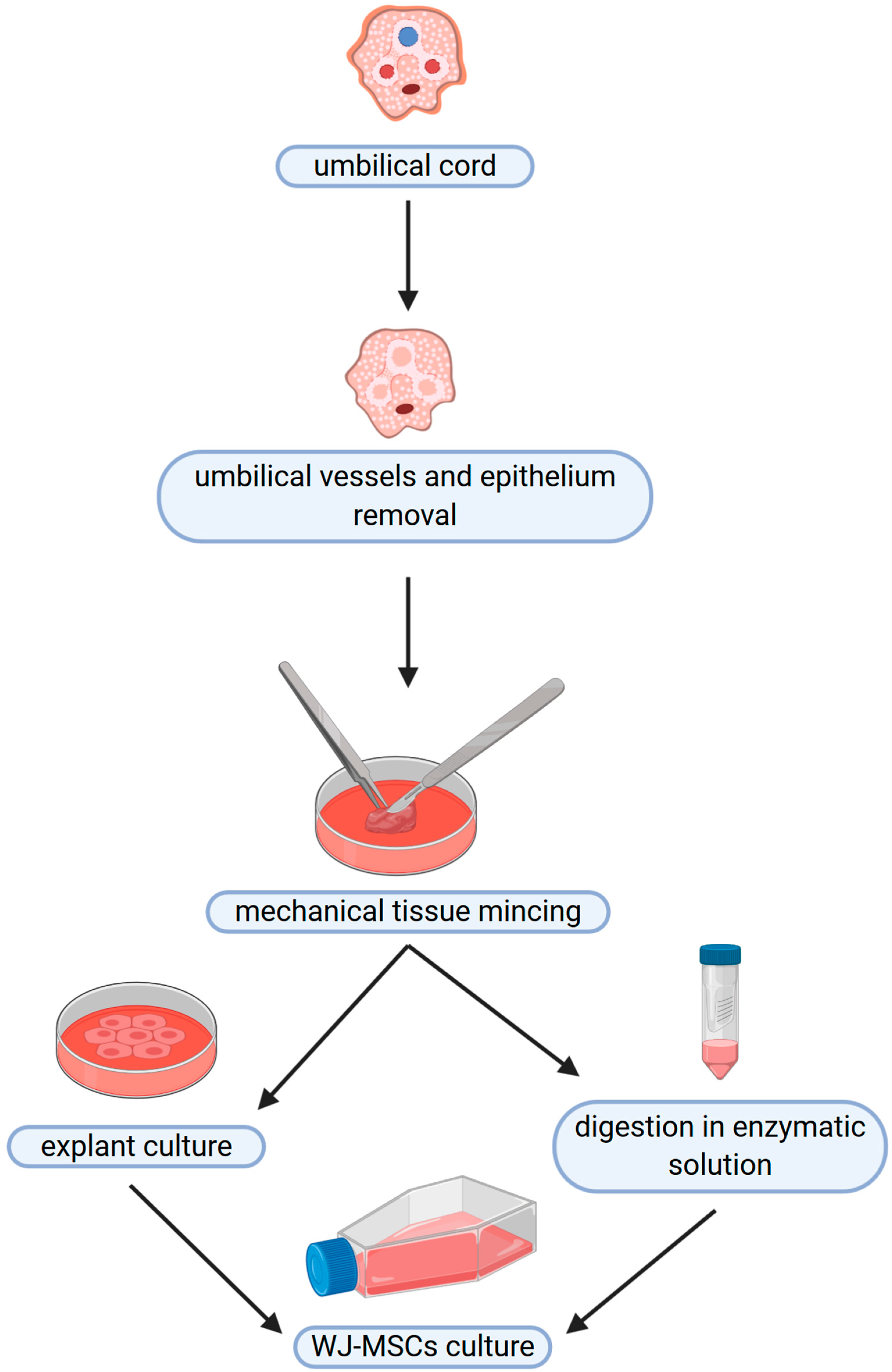
3. Wharton’s Jelly—Cellular Structure
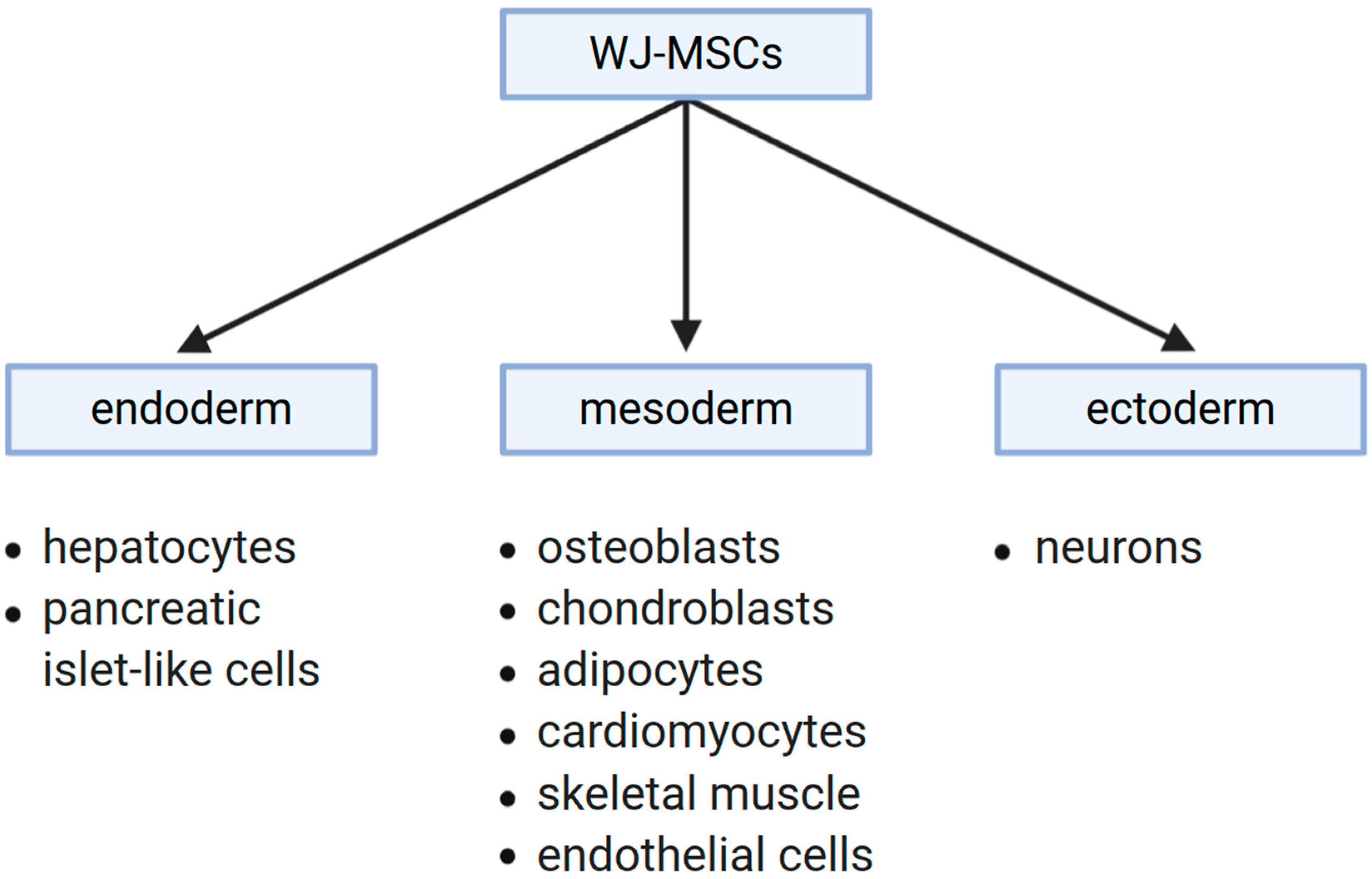
4. Stemness Specificity
5. Immunomodulatory Properties of WJ-MSCs
6. Selected Animal Models
7. Clinical application of Wharton’s jelly
8. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Doğan, A. Embryonic stem cells in development and regenerative medicine. Adv. Exp. Med. Biol. 2018, 1079, 1–15. [Google Scholar] [PubMed]
- Huang, G.; Ye, S.; Zhou, X.; Liu, D.; Ying, Q.L. Molecular basis of embryonic stem cell self-renewal: From signaling pathways to pluripotency network. Cell. Mol. Life Sci. 2015, 72, 1741–1757. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Friedenstein, A.J.; Chailakhjan, R.K.; Lalykina, K.S. the Development of fibroblast colonies in monolayer cultures of guinea-pig bone marrow and spleen sells. Cell Prolif. 1970, 3, 393–403. [Google Scholar] [CrossRef] [PubMed]
- Gruber, H.E.; Deepe, R.; Hoelscher, G.L.; Ingram, J.A.; Norton, H.J.; Scannell, B.; Loeffler, B.J.; Zinchenko, N.; Hanley, E.N.; Tapp, H. Human adipose-derived mesenchymal stem cells: Direction to a phenotype sharing similarities with the disc, gene expression profiling, and coculture with human annulus cells. Tissue Eng. Part A 2010, 16, 2843–2860. [Google Scholar] [CrossRef]
- Piva, E.; Tarlé, S.A.; Nör, J.E.; Zou, D.; Hatfield, E.; Guinn, T.; Eubanks, E.J.; Kaigler, D. Dental pulp tissue regeneration using dental pulp stem cells isolated and expanded in human serum. J. Endod. 2017, 43, 568–574. [Google Scholar] [CrossRef] [Green Version]
- Azizian, S.; Khatami, F.; Modaresifar, K.; Mosaffa, N.; Peirovi, H.; Tayebi, L.; Bahrami, S.; Redl, H.; Niknejad, H. Immunological compatibility status of placenta-derived stem cells is mediated by scaffold 3D structure. Artif. Cells Nanomed. Biotechnol. 2018, 46, 876–884. [Google Scholar] [CrossRef]
- Stefańska, K.; Sibiak, R.; Hutchings, G.; Dompe, C.; Moncrieff, L.; Janowicz, K.; Jeseta, M.; Kempisty, B.; Machatkova, M.; Mozdziak, P. Evidence for existence of molecular stemness markers in porcine ovarian follicular granulosa cells. Med. J. Cell Biol. 2019, 7, 183–188. [Google Scholar] [CrossRef] [Green Version]
- Kranc, W.; Brązert, M.; Budna, J.; Celichowski, P.; Bryja, A.; Nawrocki, M.J.; Ożegowska, K.; Jankowski, M.; Chermuła, B.; Dyszkiewicz-Konwińska, M.; et al. Genes responsible for proliferation, differentiation, and junction adhesion are significantly up-regulated in human ovarian granulosa cells during a long-term primary in vitro culture. Histochem. Cell Biol. 2019, 151, 125–143. [Google Scholar] [CrossRef] [Green Version]
- Dyszkiewicz-Konwińska, M.; Bryja, A.; Jopek, K.; Budna, J.; Khozmi, R.; Jeseta, M.; Bukowska, D.; Antosik, P.; Bruska, M.; Nowicki, M.; et al. Expression of genes responsible for cell morphogenesis involved in differentiation in porcine buccal pouch mucosal cells during long-term primary culture and real-time proliferation in vitro. J. Biol. Regul. Homeost. Agents 2017, 31, 855–864. [Google Scholar]
- Mohamadi, Y.; Noori Moghahi, S.M.H.; Mousavi, M.; Borhani-Haghighi, M.; Abolhassani, F.; Kashani, I.R.; Hassanzadeh, G. Intrathecal transplantation of Wharton’s jelly mesenchymal stem cells suppresses the NLRP1 inflammasome in the rat model of spinal cord injury. J. Chem. Neuroanat. 2019, 97, 1–8. [Google Scholar] [CrossRef]
- Baksh, D.; Yao, R.; Tuan, R.S. Comparison of proliferative and multilineage differentiation potential of human mesenchymal stem cells derived from umbilical cord and bone marrow. Stem Cells 2007, 25, 1384–1392. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takahashi, K.; Tanabe, K.; Ohnuki, M.; Narita, M.; Ichisaka, T.; Tomoda, K.; Yamanaka, S. Induction of pluripotent stem cells from adult human fibroblasts by defined factors. Cell 2007, 131, 861–872. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brix, J.; Zhou, Y.; Luo, Y. The epigenetic reprogramming roadmap in generation of iPSCs from somatic cells. J. Genet. Genom. 2015, 42, 661–670. [Google Scholar] [CrossRef] [PubMed]
- Noguchi, H.; Miyagi-Shiohira, C.; Nakashima, Y. Induced tissue-specific stem cells and epigenetic memory in induced pluripotent stem cells. Int. J. Mol. Sci. 2018, 19, 930. [Google Scholar] [CrossRef] [Green Version]
- Bar-Nur, O.; Russ, H.A.; Efrat, S.; Benvenisty, N. Epigenetic memory and preferential lineage-specific differentiation in induced pluripotent stem cells derived from human pancreatic islet beta cells. Cell Stem Cell 2011, 9, 17–23. [Google Scholar] [CrossRef] [Green Version]
- Subramanian, A.; Fong, C.Y.; Biswas, A.; Bongso, A. Comparative characterization of cells from the various compartments of the human umbilical cord shows that the Wharton’s jelly compartment provides the best source of clinically utilizable mesenchymal stem cells. PLoS ONE 2015, 10, e0127992. [Google Scholar] [CrossRef]
- Arutyunyan, I.; Elchaninov, A.; Makarov, A.; Fatkhudinov, T. Umbilical cord as prospective source for mesenchymal stem cell-based therapy. Stem Cells Int. 2016, 2016, 6901286. [Google Scholar] [CrossRef] [Green Version]
- Conconi, M.T.; Di Liddo, R.; Tommasini, M.; Calore, C.; Parnigotto, P.P. Phenotype and differentiation potential of stromal populations obtained from various zones of human umbilical cord: An overview. Open Tissue Eng. Regen. Med. J. 2011, 4, 6–20. [Google Scholar] [CrossRef] [Green Version]
- Di Naro, E.; Ghezzi, F.; Raio, L.; Franchi, M.; D’Addario, V. Umbilical cord morphology and pregnancy outcome. Eur. J. Obstet. Gynecol. Reprod. Biol. 2001, 96, 150–157. [Google Scholar] [CrossRef]
- Spurway, J.; Logan, P.; Pak, S. The development, structure and blood flow within the umbilical cord with particular reference to the venous system. Australas. J. Ultrasound Med. 2012, 15, 97–102. [Google Scholar] [CrossRef] [Green Version]
- Covas, D.T.; Siufi, J.L.C.; Silva, A.R.L.; Orellana, M.D. Isolation and culture of umbilical vein mesenchymal stem cells. Braz. J. Med. Biol. Res. 2003, 36, 1179–1183. [Google Scholar] [CrossRef] [Green Version]
- Panepucci, R.A.; Siufi, J.L.C.; Silva, W.A.; Proto-Siquiera, R.; Neder, L.; Orellana, M.; Rocha, V.; Covas, D.T.; Zago, M.A. Comparison of gene expression of umbilical cord vein and bone marrow-derived mesenchymal stem cells. Stem Cells 2004, 22, 1263–1278. [Google Scholar] [CrossRef]
- Raio, L.; Ghezzi, F.; Di Naro, E.; Franchi, M.; Briihwiler, H. Prenatal assessment of the Hyrtl anastomosis and evaluation of its function. Hum. Reprod. 1999, 14, 1890–1893. [Google Scholar] [CrossRef] [Green Version]
- Lyons, F.G.; Mattei, T.A. Sources, identification, and clinical implications of heterogeneity in human umbilical cord stem cells. Adv. Exp. Med. Biol. 2019, 1169, 243–256. [Google Scholar] [PubMed]
- Nanaev, A.K.; Kohnen, G.; Milovanov, A.P.; Domogatsky, S.P.; Kaufmann, P. Stromal differentiation and architecture of the human umbilical cord. Placenta 1997, 18, 53–64. [Google Scholar] [CrossRef]
- Can, A.; Karahuseyinoglu, S. Concise review: Human umbilical cord stroma with regard to the source of fetus-derived stem cells. Stem Cells 2007, 25, 2886–2895. [Google Scholar] [CrossRef] [PubMed]
- Parry, E.W. Some electron microscope observations on the mesenchymal structures of full-term umbilical cord. J. Anat. 1970, 107, 505–518. [Google Scholar] [PubMed]
- Davies, J.E.; Walker, J.T.; Keating, A. Concise review: Wharton’s jelly: The rich, but enigmatic, source of mesenchymal stromal cells. Stem Cells Transl. Med. 2017, 6, 1620–1630. [Google Scholar] [CrossRef]
- Lim, I.J.; Phan, T.T. Epithelial and mesenchymal stem cells from the umbilical cord lining membrane. Cell Transplant. 2014, 23, 497–503. [Google Scholar] [CrossRef]
- Ruetze, M.; Gallinat, S.; Lim, I.J.; Chow, E.; Phan, T.T.; Staeb, F.; Wenck, H.; Deppert, W.; Knott, A. Common features of umbilical cord epithelial cells and epidermal keratinocytes. J. Dermatol. Sci. 2008, 50, 227–231. [Google Scholar] [CrossRef]
- Mizoguchi, M.; Ikeda, S.; Suga, Y.; Ogawa, H. Expression of cytokeratins and cornified cell envelope-associated proteins in umbilical cord epithelium: A comparative study of the umbilical cord, amniotic epithelia, and fetal skin. J. Invest. Dermatol. 2000, 115, 133–134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mizoguchi, M.; Suga, Y.; Sanmano, B.; Ikeda, S.; Ogawa, H. Organotypic culture and surface plantation using umbilical cord epithelial cells: Morphogenesis and expression of differentiation markers mimicking cutaneous epidermis. J. Dermatol. Sci. 2004, 35, 199–206. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez, R.; Griparic, L.; Umana, M.; Burgee, K.; Vargas, V.; Nasrallah, R.; Silva, F.; Patel, A. An efficient approach to isolation and characterization of pre- and postnatal umbilical cord lining stem cells for clinical applications. Cell Transplant. 2010, 19, 1439–1449. [Google Scholar] [CrossRef] [PubMed]
- McElreavey, K.D.; Irvine, A.I.; Ennis, K.T.; McLean, W.H.I. Isolation, culture and characterisation of fibroblast-like cells derived from the Wharton’s jelly portion of human umbilical cord. Proc. Biochem. Soc. Trans. 1991, 19, 29S. [Google Scholar] [CrossRef] [PubMed]
- Takechi, K.; Kuwabara, Y.; Mizuno, M. Ultrastructural and immunohistochemical studies of Wharton’s jelly umbilical cord cells. Placenta 1993, 14, 235–245. [Google Scholar] [CrossRef]
- Kouloumenta, A.; Mavroidis, M.; Capetanaki, Y. Proper perinuclear localization of the TRIM-like protein myospryn requires its binding partner desmin. J. Biol. Chem. 2007, 282, 35211–35221. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lazarides, E. Intermediate filaments: A chemically heterogeneous, developmentally regulated class of proteins. Annu. Rev. Biochem. 1982, 51, 219–250. [Google Scholar] [CrossRef]
- Gabbiani, G.; Ryan, G.B.; Majno, G. Presence of modified fibroblasts in granulation tissue and their possible role in wound contraction. Experientia 1971, 27, 549–550. [Google Scholar] [CrossRef]
- Kobayashi, K.; Kubota, T.; Aso, T. Study on myofibroblast differentiation in the stromal cells of Wharton’s jelly. Expression and localization of α-smooth muscle actin. Early Hum. Dev. 1998, 51, 223–233. [Google Scholar] [CrossRef]
- Deasy, B.M.; Schugar, R.C.; Chirieleison, S.M.; Wescoe, K.E.; Schmidt, B.T.; Askew, Y.; Nance, J.J.; Evron, J.M.; Peault, B. High harvest yield, high expansion, and phenotype stability of CD146 mesenchymal stromal cells from whole primitive human umbilical cord tissue. J. Biomed. Biotechnol. 2009, 2009, 789526. [Google Scholar]
- Meyer, F.A.; Laver-Rudich, Z.; Tanenbaum, R. Evidence for a mechanical coupling of glycoprotein microfibrils with collagen fibrils in Wharton’s jelly. BBA-Gen. Subj. 1983, 755, 376–387. [Google Scholar] [CrossRef]
- Sobolewski, K.; Bańkowski, E.; Chyczewski, L.; Jaworski, S. Collagen and glycosaminoglycans of wharton’s jelly. Neonatology 1997, 71, 11–21. [Google Scholar] [CrossRef] [PubMed]
- Ryynanen, J.; Tan, E.M.L.; Hoffren, J.; Woodley, D.T.; Sollberg, S. Type VII collagen gene expression in human umbilical tissue and cells. Lab. Investig. 1993, 69, 300–304. [Google Scholar]
- Franc, S.; Rousseau, J.C.; Garrone, R.; Van Der Rest, M.; Moradi-Améli, M. Microfibrillar composition of umbilical cord matrix: Characterization of fibrillin, collagen VI and intact collagen V. Placenta 1998, 19, 95–104. [Google Scholar] [CrossRef]
- Damsgaard, T.M.E.; Nielsen, B.W.; Sørensen, F.B.; Henriques, U.; Schiøtz, P.O. Estimation of the total number of mast cells in the human umbilical cord: A methodological study. Apmis 1992, 100, 845–850. [Google Scholar] [CrossRef]
- Dominici, M.; Le Blanc, K.; Mueller, I.; Slaper-Cortenbach, I.; Marini, F.C.; Krause, D.S.; Deans, R.J.; Keating, A.; Prockop, D.J.; Horwitz, E.M. Minimal criteria for defining multipotent mesenchymal stromal cells. The International Society for Cellular Therapy position statement. Cytotherapy 2006, 8, 315–317. [Google Scholar] [CrossRef]
- Mushahary, D.; Spittler, A.; Kasper, C.; Weber, V.; Charwat, V. Isolation, cultivation, and characterization of human mesenchymal stem cells. Cytom. Part A 2018, 93, 19–31. [Google Scholar] [CrossRef] [Green Version]
- Romanov, Y.A. Searching for alternative sources of postnatal human mesenchymal stem cells: Candidate MSC-like cells from umbilical cord. Stem Cells 2003, 21, 105–110. [Google Scholar] [CrossRef] [Green Version]
- Kadivar, M.; Khatami, S.; Mortazavi, Y.; Shokrgozar, M.A.; Taghikhani, M.; Soleimani, M. In vitro cardiomyogenic potential of human umbilical vein-derived mesenchymal stem cells. Biochem. Biophys. Res. Commun. 2006, 340, 639–647. [Google Scholar] [CrossRef]
- Reza, H.M.; Ng, B.Y.; Phan, T.T.; Tan, D.T.H.; Beuerman, R.W.; Ang, L.P.K. Characterization of a novel umbilical cord lining cell with CD227 positivity and unique pattern of P63 expression and function. Stem Cell Rev. Rep. 2011, 7, 624–638. [Google Scholar] [CrossRef]
- Campard, D.; Lysy, P.A.; Najimi, M.; Sokal, E.M. Native umbilical cord matrix stem cells express hepatic markers and differentiate into hepatocyte-like cells. Gastroenterology 2008, 134, 833–848. [Google Scholar] [CrossRef] [PubMed]
- Weiss, M.L.; Anderson, C.; Medicetty, S.; Seshareddy, K.B.; Weiss, R.J.; VanderWerff, I.; Troyer, D.; McIntosh, K.R. Immune properties of human umbilical cord Wharton’s jelly-derived cells. Stem Cells 2008, 26, 2865–2874. [Google Scholar] [CrossRef] [PubMed]
- Martin-Rendon, E.; Sweeney, D.; Lu, F.; Girdlestone, J.; Navarrete, C.; Watt, S.M. 5-Azacytidine-treated human mesenchymal stem/progenitor cells derived from umbilical cord, cord blood and bone marrow do not generate cardiomyocytes in vitro at high frequencies. Vox Sang. 2008, 95, 137–148. [Google Scholar] [CrossRef] [PubMed]
- Sarugaser, R.; Lickorish, D.; Baksh, D.; Hosseini, M.M.; Davies, J.E. Human umbilical cord perivascular (HUCPV) cells: A source of mesenchymal progenitors. Stem Cells 2005, 23, 220–229. [Google Scholar] [CrossRef]
- Sarugaser, R.; Hanoun, L.; Keating, A.; Stanford, W.L.; Davies, J.E. Human mesenchymal stem cells self-renew and differentiate according to a deterministic hierarchy. PLoS ONE 2009, 4, e6498. [Google Scholar] [CrossRef] [Green Version]
- Vangsness, C.T.; Sternberg, H.; Harris, L. Umbilical cord tissue offers the greatest number of harvestable mesenchymal stem cells for research and clinical application: A literature review of different harvest sites. Arthrosc.-J. Arthrosc. Relat. Surg. 2015, 31, 1836–1843. [Google Scholar] [CrossRef]
- Carlin, R.; Davis, D.; Weiss, M.; Schultz, B.; Troyer, D. Expression of early transcription factors Oct-4, Sox-2 and Nanog by porcine umbilical cord (PUC) matrix cells. Reprod. Biol. Endocrinol. 2006, 4, 8. [Google Scholar] [CrossRef] [Green Version]
- Tong, C.K.; Vellasamy, S.; Chong Tan, B.; Abdullah, M.; Vidyadaran, S.; Fong Seow, H.; Ramasamy, R. Generation of mesenchymal stem cell from human umbilical cord tissue using a combination enzymatic and mechanical disassociation method. Cell Biol. Int. 2011, 35, 221–226. [Google Scholar] [CrossRef]
- Oktar, P.A.; Yildirim, S.; Balci, D.; Can, A. Continual expression throughout the cell cycle and downregulation upon adipogenic differentiation makes cucleostemin a vital human MSC proliferation marker. Stem Cell Rev. Rep. 2011, 7, 413–424. [Google Scholar] [CrossRef]
- Fong, C.Y.; Richards, M.; Manasi, N.; Biswas, A.; Bongso, A. Comparative growth behaviour and characterization of stem cells from human Wharton’s jelly. Reprod. Biomed. Online 2007, 15, 708–718. [Google Scholar] [CrossRef]
- Wang, H.-S.; Hung, S.-C.; Peng, S.-T.; Huang, C.-C.; Wei, H.-M.; Guo, Y.-J.; Fu, Y.-S.; Lai, M.-C.; Chen, C.-C. Mesenchymal stem cells in the Wharton’s jelly of the human umbilical cord. Stem Cells 2004, 22, 1330–1337. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.N.; Lie, P.C.; Wei, X. Differentiation of mesenchymal stromal cells derived from umbilical cord Wharton’s jelly into hepatocyte-like cells. Cytotherapy 2009, 11, 548–558. [Google Scholar] [CrossRef]
- Xu, M.; Zhang, B.; Liu, Y.; Zhang, J.; Sheng, H.; Shi, R.; Liao, L.; Liu, N.; Hu, J.; Wang, J.; et al. The immunologic and hematopoietic profiles of Mesenchymal stem cells derived from different sections of human umbilical cord. Acta Biochim. Biophys. Sin. 2014, 46, 1056–1065. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ranjbaran, H.; Abediankenari, S.; Mohammadi, M.; Jafari, N.; Khalilian, A.; Rahmani, Z.; Amiri, M.M.; Ebrahimi, P. Wharton’s jelly derived-mesenchymal stem cells: Isolation and characterization. Acta Med. Iran. 2018, 56, 28–33. [Google Scholar] [PubMed]
- Bharti, D.; Shivakumar, S.B.; Park, J.K.; Ullah, I.; Subbarao, R.B.; Park, J.S.; Lee, S.L.; Park, B.W.; Rho, G.J. Comparative analysis of human Wharton’s jelly mesenchymal stem cells derived from different parts of the same umbilical cord. Cell Tissue Res. 2018, 372, 51–65. [Google Scholar] [CrossRef] [Green Version]
- Karahuseyinoglu, S.; Cinar, O.; Kilic, E.; Kara, F.; Akay, G.G.; Demiralp, D.Ö.; Tukun, A.; Uckan, D.; Can, A. Biology of stem cells in human umbilical cord stroma: In situ and in vitro surveys. Stem Cells 2007, 25, 319–331. [Google Scholar] [CrossRef] [PubMed]
- Conconi, M.T.; Burra, P.; Di Liddo, R.; Calore, C.; Turetta, M.; Bellini, S.; Bo, P.; Nussdorfer, G.G.; Parnigotto, P.P. CD105(+) cells from Wharton’s jelly show in vitro and in vivo myogenic differentiative potential. Int. J. Mol. Med. 2006, 18, 1089–1096. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mitchell, K.E.; Weiss, M.L.; Mitchell, B.M.; Martin, P.; Davis, D.; Morales, L.; Helwig, B.; Beerenstrauch, M.; Abou-Easa, K.; Hildreth, T.; et al. Matrix cells from Wharton’s jelly form neurons and glia. Stem Cells 2003, 21, 50–60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fu, Y.S.; Shih, Y.T.; Cheng, Y.C.; Min, M.Y. Transformation of human umbilical mesenchymal cells into neurons in vitro. J. Biomed. Sci. 2004, 11, 652–660. [Google Scholar] [CrossRef]
- Liang, J.; Wu, S.; Zhao, H.; Li, S.L.; Liu, Z.X.; Wu, J.; Zhou, L. Human umbilical cord mesenchymal stem cells derived from Wharton’s jelly differentiate into cholinergic-like neurons in vitro. Neurosci. Lett. 2013, 532, 59–63. [Google Scholar] [CrossRef]
- Chao, K.C.; Chao, K.F.; Fu, Y.S.; Liu, S.H. Islet-like clusters derived from mesenchymal stem cells in Wharton’s jelly of the human umbilical cord for transplantation to control type 1 diabetes. PLoS ONE 2008, 3, e1451. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, K.H.; Zhou, B.; Lu, S.H.; Feng, B.; Yang, S.G.; Du, W.T.; Gu, D.S.; Han, Z.C.; Liu, Y.L. In vitro and in vivo differentiation of human umbilical cord derived stem cells into endothelial cells. J. Cell. Biochem. 2007, 100, 608–616. [Google Scholar] [CrossRef] [PubMed]
- Huang, P.; Lin, L.M.; Wu, X.Y.; Tang, Q.L.; Feng, X.Y.; Lin, G.Y.; Lin, X.; Wang, H.W.; Huang, T.H.; Ma, L. Differentiation of human umbilical cord Wharton’s jelly-derived mesenchymal stem cells into germ-like cells in vitro. J. Cell. Biochem. 2010, 109, 747–754. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; Liang, J.; Cui, H.P.; Wang, X.M.; Rong, H.; Shao, B.; Cui, H. Wharton’s jelly mesenchymal stem cells differentiate into retinal progenitor cells. Neural Regen. Res. 2013, 8, 1783–1792. [Google Scholar] [PubMed]
- Zhou, C.; Yang, B.; Tian, Y.; Jiao, H.; Zheng, W.; Wang, J.; Guan, F. Immunomodulatory effect of human umbilical cord Wharton’s jelly-derived mesenchymal stem cells on lymphocytes. Cell. Immunol. 2011, 272, 33–38. [Google Scholar] [CrossRef] [Green Version]
- Karaöz, E.; Demircan, P.Ç.; Erman, G.; Güngörürler, E.; Sarıboyacı, A.E. Comparative analyses of immunosuppressive characteristics of bone-marrow, Wharton’s jelly, and adipose tissue-derived human mesenchymal stem cells. Turk. J. Hematol. 2017, 34, 213–225. [Google Scholar]
- Deng, Y.; Yi, S.; Wang, G.; Cheng, J.; Zhang, Y.; Chen, W.; Tai, Y.; Chen, S.; Chen, G.; Liu, W.; et al. Umbilical cord-derived mesenchymal stem cells instruct dendritic cells to acquire tolerogenic phenotypes through the IL-6-mediated upregulation of SOCS1. Stem Cells Dev. 2014, 23, 2080–2092. [Google Scholar] [CrossRef]
- Paladino, F.V.; Sardinha, L.R.; Piccinato, C.A.; Goldberg, A.C. Intrinsic variability present in Wharton’s jelly mesenchymal stem cells and T cell responses may impact cell therapy. Stem Cells Int. 2017, 2017, 8492797. [Google Scholar] [CrossRef] [Green Version]
- Donders, R.; Bogie, J.F.J.; Ravanidis, S.; Gervois, P.; Vanheusden, M.; Marée, R.; Schrynemackers, M.; Smeets, H.J.M.; Pinxteren, J.; Gijbels, K.; et al. Human Wharton’s jelly-derived stem cells display a distinct immunomodulatory and proregenerative transcriptional signature compared to bone marrow-derived stem cells. Stem Cells Dev. 2018, 27, 65–84. [Google Scholar] [CrossRef]
- Gauthaman, K.; Fong, C.Y.; Suganya, C.A.; Subramanian, A.; Biswas, A.; Choolani, M.; Bongso, A. Extra-embryonic human Wharton’s jelly stem cells do not induce tumorigenesis, unlike human embryonic stem cells. Reprod. Biomed. Online 2012, 24, 235–246. [Google Scholar] [CrossRef] [Green Version]
- Weiss, M.L.; Medicetty, S.; Bledsoe, A.R.; Rachakatla, R.S.; Choi, M.; Merchav, S.; Luo, Y.; Rao, M.S.; Velagaleti, G.; Troyer, D. Human umbilical cord matrix stem cells: Preliminary characterization and effect of transplantation in a rodent model of Parkinson’s disease. Stem Cells 2006, 24, 781–792. [Google Scholar] [CrossRef] [PubMed]
- Fu, Y.-S.; Cheng, Y.-C.; Lin, M.-Y.A.; Cheng, H.; Chu, P.-M.; Chou, S.-C.; Shih, Y.-H.; Ko, M.-H.; Sung, M.-S. Conversion of human umbilical cord mesenchymal stem cells in Wharton’s jelly to dopaminergic neurons in vitro: Potential therapeutic application for Parkinsonism. Stem Cells 2006, 24, 115–124. [Google Scholar] [CrossRef] [PubMed]
- Krupa, P.; Vackova, I.; Ruzicka, J.; Zaviskova, K.; Dubisova, J.; Koci, Z.; Turnovcova, K.; Urdzikova, L.M.; Kubinova, S.; Rehak, S.; et al. The effect of human mesenchymal stem cells derived from Wharton’s Jelly in spinal cord injury treatment is dose-dependent and can be facilitated by repeated application. Int. J. Mol. Sci. 2018, 19, 1503. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chudickova, M.; Vackova, I.; Urdzikova, L.M.; Jancova, P.; Kekulova, K.; Rehorova, M.; Turnovcova, K.; Jendelova, P.; Kubinova, S. The effect of Wharton jelly-derived mesenchymal stromal cells and their conditioned media in the treatment of a rat spinal cord injury. Int. J. Mol. Sci. 2019, 20, 4516. [Google Scholar] [CrossRef] [Green Version]
- Ding, D.C.; Shyu, W.C.; Chiang, M.F.; Lin, S.Z.; Chang, Y.C.; Wang, H.J.; Su, C.Y.; Li, H. Enhancement of neuroplasticity through upregulation of β1-integrin in human umbilical cord-derived stromal cell implanted stroke model. Neurobiol. Dis. 2007, 27, 339–353. [Google Scholar] [CrossRef]
- Kadam, S.S.; Bhonde, R.R. Islet neogenesis from the constitutively nestin expressing human umbilical cord matrix derived mesenchymal stem cells. Islets 2010, 2, 112–120. [Google Scholar] [CrossRef]
- Moodley, Y.; Atienza, D.; Manuelpillai, U.; Samuel, C.S.; Tchongue, J.; Ilancheran, S.; Boyd, R.; Trounson, A. Human umbilical cord mesenchymal stem cells reduce fibrosis of bleomycin-induced lung injury. Am. J. Pathol. 2009, 175, 303–313. [Google Scholar] [CrossRef] [Green Version]
- Tsai, P.C.; Fu, T.W.; Chen, Y.M.A.; Ko, T.L.; Chen, T.H.; Shih, Y.H.; Hung, S.C.; Fu, Y.S. The therapeutic potential of human umbilical mesenchymal stem cells from Wharton’s jelly in the treatment of rat liver fibrosis. Liver Transplant. 2009, 15, 484–495. [Google Scholar] [CrossRef]
- Sabapathy, V.; Sundaram, B.; Vm, S.; Mankuzhy, P.; Kumar, S. Human Wharton’s jelly mesenchymal stem cells plasticity augments scar-free skin wound healing with hair growth. PLoS ONE 2014, 9, e93726. [Google Scholar] [CrossRef]
- Zhang, W.; Liu, X.C.; Yang, L.; Zhu, D.L.; Zhang, Y.D.; Chen, Y.; Zhang, H.Y. Wharton’s jelly-derived mesenchymal stem cells promote myocardial regeneration and cardiac repair after miniswine acute myocardial infarction. Coron. Artery Dis. 2013, 24, 549–558. [Google Scholar] [CrossRef]
- Nascimento, D.S.; Mosqueira, D.; Sousa, L.M.; Teixeira, M.; Filipe, M.; Resende, T.P.; Araújo, A.F.; Valente, M.; Almeida, J.; Martins, J.P.; et al. Human umbilical cord tissue-derived mesenchymal stromal cells attenuate remodeling after myocardial infarction by proangiogenic, antiapoptotic, and endogenous cell-activation mechanisms. Stem Cell Res. Ther. 2014, 5, 5. [Google Scholar] [CrossRef] [Green Version]
- Matsuzuka, T.; Rachakatla, R.S.; Doi, C.; Maurya, D.K.; Ohta, N.; Kawabata, A.; Pyle, M.M.; Pickel, L.; Reischman, J.; Marini, F.; et al. Human umbilical cord matrix-derived stem cells expressing interferon-β gene significantly attenuate bronchioloalveolar carcinoma xenografts in SCID mice. Lung Cancer 2010, 70, 28–36. [Google Scholar] [CrossRef] [Green Version]
- Ayuzawa, R.; Doi, C.; Rachakatla, R.S.; Pyle, M.M.; Maurya, D.K.; Troyer, D.; Tamura, M. Naïve human umbilical cord matrix derived stem cells significantly attenuate growth of human breast cancer cells in vitro and in vivo. Cancer Lett. 2009, 280, 31–37. [Google Scholar] [CrossRef] [Green Version]
- Ma, Y.; Hao, X.; Zhang, S.; Zhang, J. The in vitro and in vivo effects of human umbilical cord mesenchymal stem cells on the growth of breast cancer cells. Breast Cancer Res. Treat. 2012, 133, 473–485. [Google Scholar] [CrossRef]
- Can, A.; Celikkan, F.T.; Cinar, O. Umbilical cord mesenchymal stromal cell transplantations: A systemic analysis of clinical trials. Cytotherapy 2017, 19, 1351–1382. [Google Scholar] [CrossRef]
- Gao, L.R.; Chen, Y.; Zhang, N.K.; Yang, X.L.; Liu, H.L.; Wang, Z.G.; Yan, X.Y.; Wang, Y.; Zhu, Z.M.; Li, T.C.; et al. Intracoronary infusion of Wharton’s jelly-derived mesenchymal stem cells in acute myocardial infarction: Double-blind, randomized controlled trial. BMC Med. 2015, 13, 162. [Google Scholar] [CrossRef] [Green Version]
- Barczewska, M.; Grudniak, M.; Maksymowicz, S.; Siwek, T.; Oldak, T.; Jezierska-Wozniak, K.; Gladysz, D.; Maksymowicz, W. Safety of intrathecal injection of Wharton’s jellyderived mesenchymal stem cells in amyotrophic lateral sclerosis therapy. Neural Regen. Res. 2019, 14, 313–318. [Google Scholar]
- Zhang, Z.; Fu, J.; Xu, X.; Wang, S.; Xu, R.; Zhao, M.; Nie, W.; Wang, X.; Zhang, J.; Li, T.; et al. Safety and immunological responses to human mesenchymal stem cell therapy in difficult-to-treat HIV-1-infected patients. Aids 2013, 27, 1283–1293. [Google Scholar] [CrossRef] [Green Version]
- Wang, D.; Li, J.; Zhang, Y.; Zhang, M.; Chen, J.; Li, X.; Hu, X.; Jiang, S.; Shi, S.; Sun, L. Umbilical cord mesenchymal stem cell transplantation in active and refractory systemic lupus erythematosus: A multicenter clinical study. Arthritis Res. Ther. 2014, 16, R79. [Google Scholar] [CrossRef] [Green Version]
- Lu, Z.; Ye, D.; Qian, L.; Zhu, L.; Wang, C.; Guan, D.; Zhang, X.; Xu, Y. Human umbilical cord mesenchymal stem cell therapy on neuromyelitis optica. Curr. Neurovasc. Res. 2012, 9, 250–255. [Google Scholar] [CrossRef]
- Wang, L.; Wang, L.; Cong, X.; Liu, G.; Zhou, J.; Bai, B.; Li, Y.; Bai, W.; Li, M.; Ji, H.; et al. Human umbilical cord mesenchymal stem cell therapy for patients with active rheumatoid arthritis: Safety and efficacy. Stem Cells Dev. 2013, 22, 3192–3202. [Google Scholar] [CrossRef] [Green Version]
- Wang, D.; Feng, X.; Lu, L.; Konkel, J.E.; Zhang, H.; Chen, Z.; Li, X.; Gao, X.; Lu, L.; Shi, S.; et al. A CD8 T cell/indoleamine 2,3-dioxygenase axis is required for mesenchymal stem cell suppression of human systemic lupus erythematosus. Arthritis Rheumatol. 2014, 66, 2234–2245. [Google Scholar] [CrossRef] [Green Version]
- Hu, J.; Yu, X.; Wang, Z.; Wang, F.; Wang, L.; Gao, H.; Chen, Y.; Zhao, W.; Jia, Z.; Yan, S.; et al. Long term effects of the implantation of Wharton’s jelly-derived mesenchymal stem cells from the umbilical cord for newly-onset type 1 diabetes mellitus. Endocr. J. 2013, 60, 347–357. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.; Zheng, P.; Wang, X.; Dai, G.; Cheng, H.; Zhang, Z.; Hua, R.; Niu, X.; Shi, J.; An, Y. A preliminary evaluation of efficacy and safety of Wharton’s jelly mesenchymal stem cell transplantation in patients with type 2 diabetes mellitus. Stem Cell Res. Ther. 2014, 5, 57. [Google Scholar] [CrossRef] [Green Version]
- Lv, Y.T.; Zhang, Y.; Liu, M.; Qiuwaxi, J.-n.-t.; Ashwood, P.; Cho, S.C.; Huan, Y.; Ge, R.C.; Chen, X.W.; Wang, Z.J.; et al. Transplantation of human cord blood mononuclear cells and umbilical cord-derived mesenchymal stem cells in autism. J. Transl. Med. 2013, 11, 196. [Google Scholar] [CrossRef] [Green Version]
- Wu, K.H.; Tsai, C.; Wu, H.P.; Sieber, M.; Peng, C.T.; Chao, Y.H. Human application of ex vivo expanded umbilical cord-derived mesenchymal stem cells: Enhance hematopoiesis after cord blood transplantation. Cell Transplant. 2013, 22, 2041–2051. [Google Scholar] [CrossRef] [Green Version]
- Fung, M.; Yuan, Y.; Atkins, H.; Shi, Q.; Bubela, T. Responsible Translation of Stem Cell Research: An Assessment of Clinical Trial Registration and Publications. Stem Cell Rep. 2017, 8, 1190–1201. [Google Scholar] [CrossRef] [Green Version]
- Daley, G.Q.; Hyun, I.; Apperley, J.F.; Barker, R.A.; Benvenisty, N.; Bredenoord, A.L.; Breuer, C.K.; Caulfield, T.; Cedars, M.I.; Frey-Vasconcells, J.; et al. Setting global standards for stem cell research and clinical translation: The 2016 ISSCR guidelines. Stem Cell Rep. 2016, 6, 787–797. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Treated Condition | Animal Model | Mean Dose of Stem Cells in One Injection | Source of MSCs | Delivery Method | Author | Publication Date | Journal |
|---|---|---|---|---|---|---|---|
| Parkinson’s disease | Hemiparkinsonian Sprague-Dawley rats | 1 × 103 | Wharton’s jelly | Intrastriatal injection | Weiss et al. [81] | 2006 | Stem Cells |
| Parkinsonian Sprague-Dawley rats | 1 × 105 | Wharton’s jelly | Intrastriatal injection | Fu et al. [82] | 2006 | Stem Cells | |
| Spinal cord injury | Wistar rats with spinal cord injury | 0.5 or 1.5 × 106 | Wharton’s jelly | Intrathecal infusion | Krupa et al. [83] | 2018 | International Journal of Molecular Sciences |
| Wistar rats with spinal cord injury | 1.5 × 106 | Wharton’s jelly | Intrathecal infusion | Chudickova et al. [84] | 2019 | International Journal of Molecular Sciences | |
| Wistar rats with spinal cord injury | 3 × 105 | Wharton’s jelly | Intrathecal infusion | Mohamadi et al. [10] | 2019 | Journal of Chemical Neuroanatomy | |
| Hindlimb ischemia | Athymic nude mice with hindlimb ischemia | 1 × 106 | Umbilical cord (not specified) | Injection to the adductor muscle | Wu et al. [72] | 2007 | Journal of Cellular Biochemistry |
| Brain ischemia | Sprague-Dawley rats with brain ischemia | 1 × 106 | Wharton’s jelly | Intracerebral injection | Ding et al. [85] | 2007 | Neurobiology of Disease |
| Diabetes | Streptozotocin-induced diabetic Balb/C mice | 3 × 106 of undifferentiated cells or 1 × 103 islet-like clusters | Umbilical cord (not specified) | Transplantation of encapsulated cells into abdomen | Kadam et al. [86] | 2010 | Islets |
| Pulmonary fibrosis | Bleomycin-induced lung injury in SCID mice | 1 × 106 | Wharton’s jelly | Intravenous injection | Moodley et al. [87] | 2009 | The American Journal of Pathology |
| Liver fibrosis | Sprague-Dawley rats with CCl4-induced liver fibrosis | 5 × 105 | Wharton’s jelly | Injection into the right lobe of the liver | Tsai et al. [88] | 2009 | Liver Transplantation |
| Skin injury | SCID mice with skin injury | 1 × 106 | Wharton’s jelly | Injection into the skin or | Sabapathy et al. [89] | 2014 | PLoS One |
| Myocardial infarction | Guangxi Bama miniswines with acute myocardial infarction | 4 × 107 | Wharton’s jelly | Injection into the ischemic region of the heart | Zhang et al. [90] | 2013 | Coronary Artery Disease |
| C57BL/6 mice with myocardial infarction | 2 × 105 | Umbilical cord (not specified) | Intramyocardial injection | Nascimento et al. [91] | 2014 | Stem Cell Research and Therapy | |
| Cancer | CB17 SCID mice with bronchioloalveolar carcinoma | 3 × 105 | Wharton’s jelly | Intravenous injection | Matsuzuka et al. [92] | 2010 | Lung Cancer |
| CB17 SCID mice transplanted with MDA 231 human breast carcinoma cells | 5 × 105 | Wharton’s jelly | Intravenous injection | Ayuzawa et al. [93] | 2009 | Cancer Letters | |
| CB17 SCID mice transplanted with MDA 231 human breast carcinoma cells | 0.5 × 106 or1 × 106 or 3 × 106 | Wharton’s jelly | Subcutaneous injection at the tumor site | Ma et al. [94] | 2012 | Breast Cancer Research and Treatment |
| Treated Condition | Number of Study Participants | Type of Study | Source of Stem Cells/Number of Donors | Mean Dose of Stem Cells in One Injection | Delivery Method | Author | Publication Date | Journal |
|---|---|---|---|---|---|---|---|---|
| Acute myocardial infarction | 116 | Randomized double-blind controlled trial | Wharton’s jelly/pooled from 21 donors | 6 × 106 | Intracoronary infusion | Gao et al. [96] | 2015 | BMC Medicine |
| Amyotrophic lateral sclerosis | 43 | Not specified | Wharton’s jelly/pooled from three donors | 0.42 × 106/kg of body weight | Intrathecal injection | Barczewska et al. [97] | 2019 | Neural Regeneration Research |
| HIV-1-infected immune nonresponders | 13 | Prospective open-labeled controlled trial | Wharton’s jelly/not specified | 0.5 × 106/kg of body weight | Intravenous infusion | Zhang et al. [98] | 2013 | AIDS |
| Active and refractory systemic lupus erythematosus | 40 | Multicenter trial | Umbilical cord (not specified)/not specified | 1 × 106/kg of body weight | Intravenous infusion | Wang et al. [99,102] | 2014; 2014 | Arthritis Research and Therapy; Arthritis and Rheumatology |
| Neuromyelitis optica | 5 | Not specified | Umbilical cord (not specified)/not specified | 4 × 107 or 2 × 107 | Intravenous and intrathecal infusion | Lu et al. [100] | 2012 | Current Neurovascular Research |
| Rheumatoid arthritis | 172 | Single center trial | Umbilical cord (not specified)/not specified | 4 × 107 | Intravenous infusion | Wang et al. [101] | 2013 | Stem Cells and Development |
| Type 1 diabetes mellitus | 29 | Randomized double-blind controlled trial | Wharton’s jelly/one donor | 2.6 ± 1.2 × 107 | Intravenous infusion | Hu et al. [103] | 2013 | Endocrine Journal |
| Type 2 diabetes mellitus | 22 | Single center prospective trial | Wharton’s jelly/not specified | 1 × 106/kg of body weight | Intravenous infusion and intra-pancreatic endovascular injection | Liu et al. [104] | 2014 | Stem Cell Research and Therapy |
| Autism | 37 | Non-randomized open-label, single center trial | Wharton’s jelly/not specified | 1 × 106/kg of body weight | Intravenous and intrathecal infusion | Lv et al. [105] | 2013 | Journal of Translational Medicine |
| High risk leukemia | 20 | Randomized trial | Wharton’s jelly/not specified | 1 × 106/kg of body weight | Intravenous infusion | Wu et al. [106] | 2013 | Cell Transplantation |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Stefańska, K.; Ożegowska, K.; Hutchings, G.; Popis, M.; Moncrieff, L.; Dompe, C.; Janowicz, K.; Pieńkowski, W.; Gutaj, P.; Shibli, J.A.; et al. Human Wharton’s Jelly—Cellular Specificity, Stemness Potency, Animal Models, and Current Application in Human Clinical Trials. J. Clin. Med. 2020, 9, 1102. https://doi.org/10.3390/jcm9041102
Stefańska K, Ożegowska K, Hutchings G, Popis M, Moncrieff L, Dompe C, Janowicz K, Pieńkowski W, Gutaj P, Shibli JA, et al. Human Wharton’s Jelly—Cellular Specificity, Stemness Potency, Animal Models, and Current Application in Human Clinical Trials. Journal of Clinical Medicine. 2020; 9(4):1102. https://doi.org/10.3390/jcm9041102
Chicago/Turabian StyleStefańska, Katarzyna, Katarzyna Ożegowska, Greg Hutchings, Małgorzata Popis, Lisa Moncrieff, Claudia Dompe, Krzysztof Janowicz, Wojciech Pieńkowski, Paweł Gutaj, Jamil A. Shibli, and et al. 2020. "Human Wharton’s Jelly—Cellular Specificity, Stemness Potency, Animal Models, and Current Application in Human Clinical Trials" Journal of Clinical Medicine 9, no. 4: 1102. https://doi.org/10.3390/jcm9041102
APA StyleStefańska, K., Ożegowska, K., Hutchings, G., Popis, M., Moncrieff, L., Dompe, C., Janowicz, K., Pieńkowski, W., Gutaj, P., Shibli, J. A., Prado, W. M., Piotrowska-Kempisty, H., Mozdziak, P., Bruska, M., Zabel, M., Kempisty, B., & Nowicki, M. (2020). Human Wharton’s Jelly—Cellular Specificity, Stemness Potency, Animal Models, and Current Application in Human Clinical Trials. Journal of Clinical Medicine, 9(4), 1102. https://doi.org/10.3390/jcm9041102