Biocompatibility of Bone Marrow-Derived Mesenchymal Stem Cells in the Rat Inner Ear following Trans-Tympanic Administration


 , , , ,
, , , , 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Experimental Design
2.3. Generation of Rat BM-MSCs
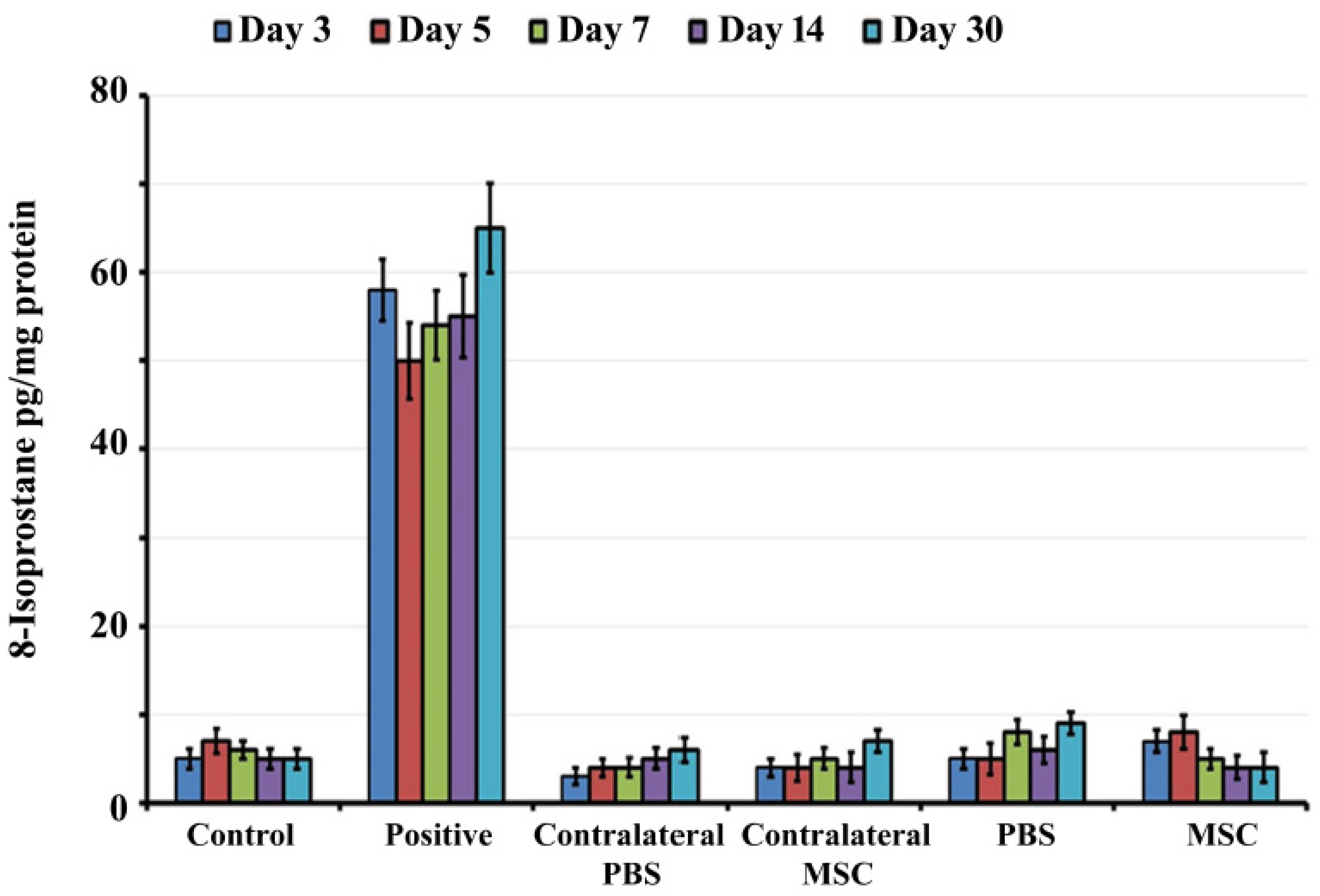
2.4. 8-Isoprostane Determination
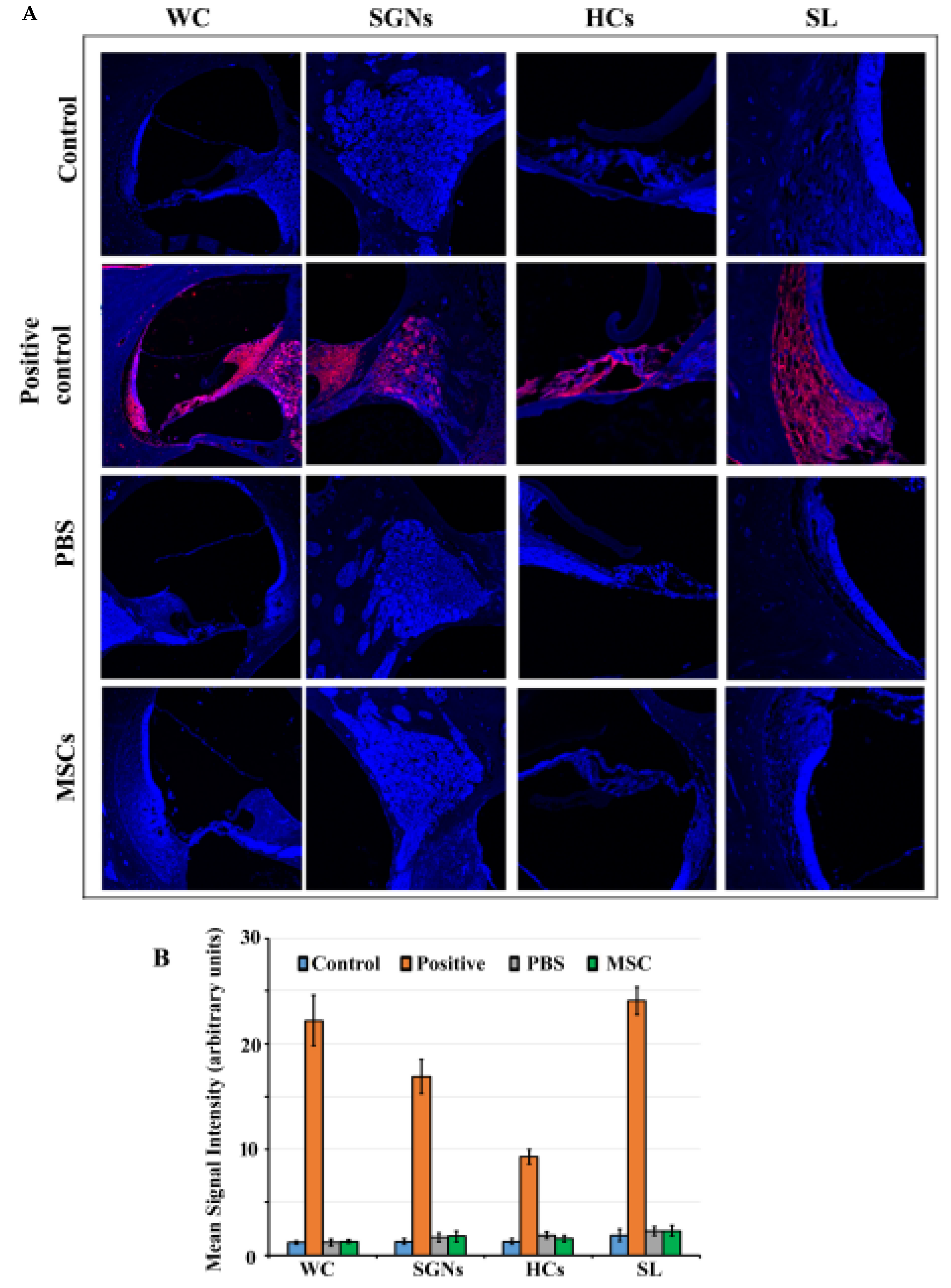
2.5. Cleaved-Caspase 3 Immunostaining
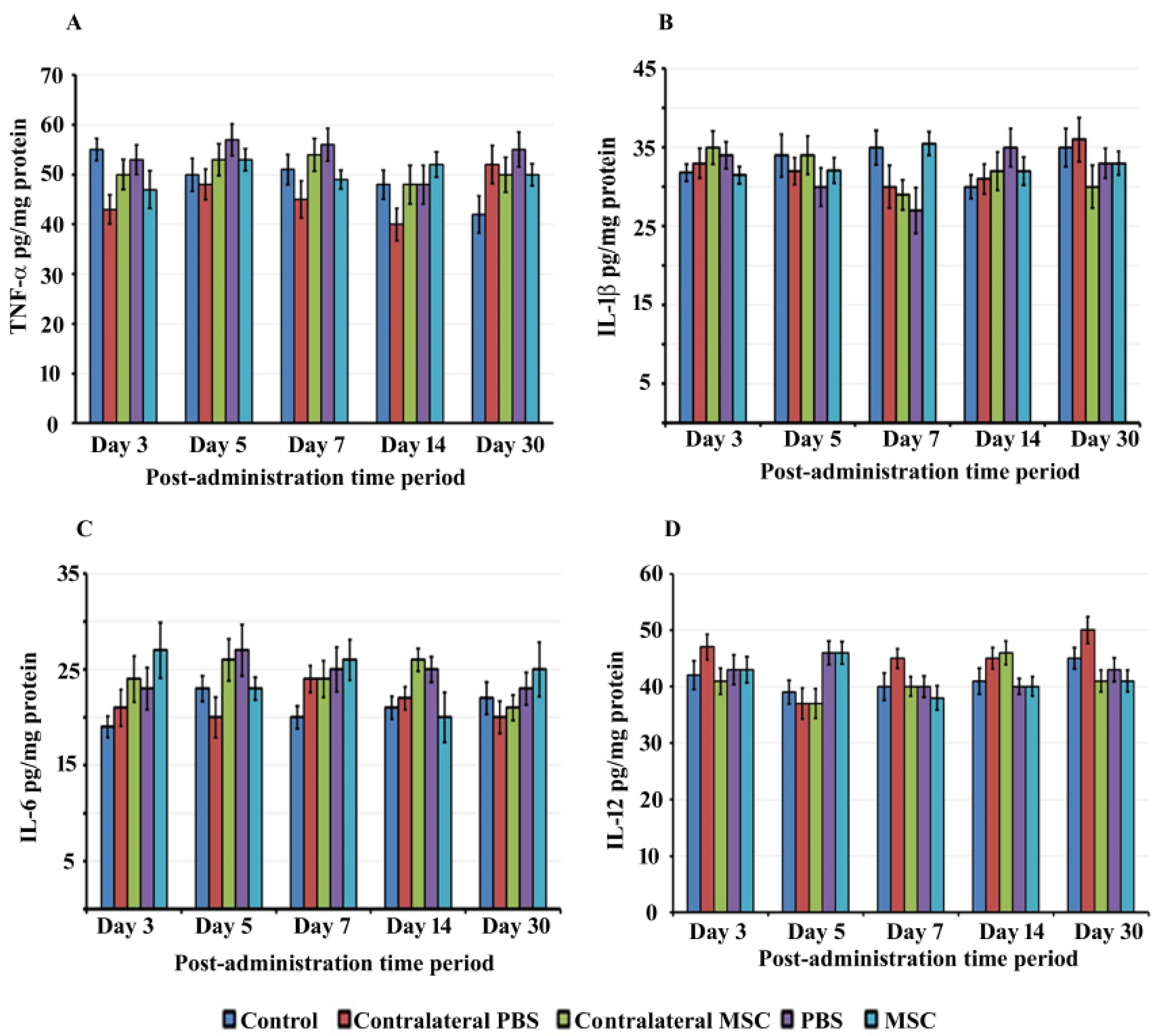
2.6. Cytokine Determination:
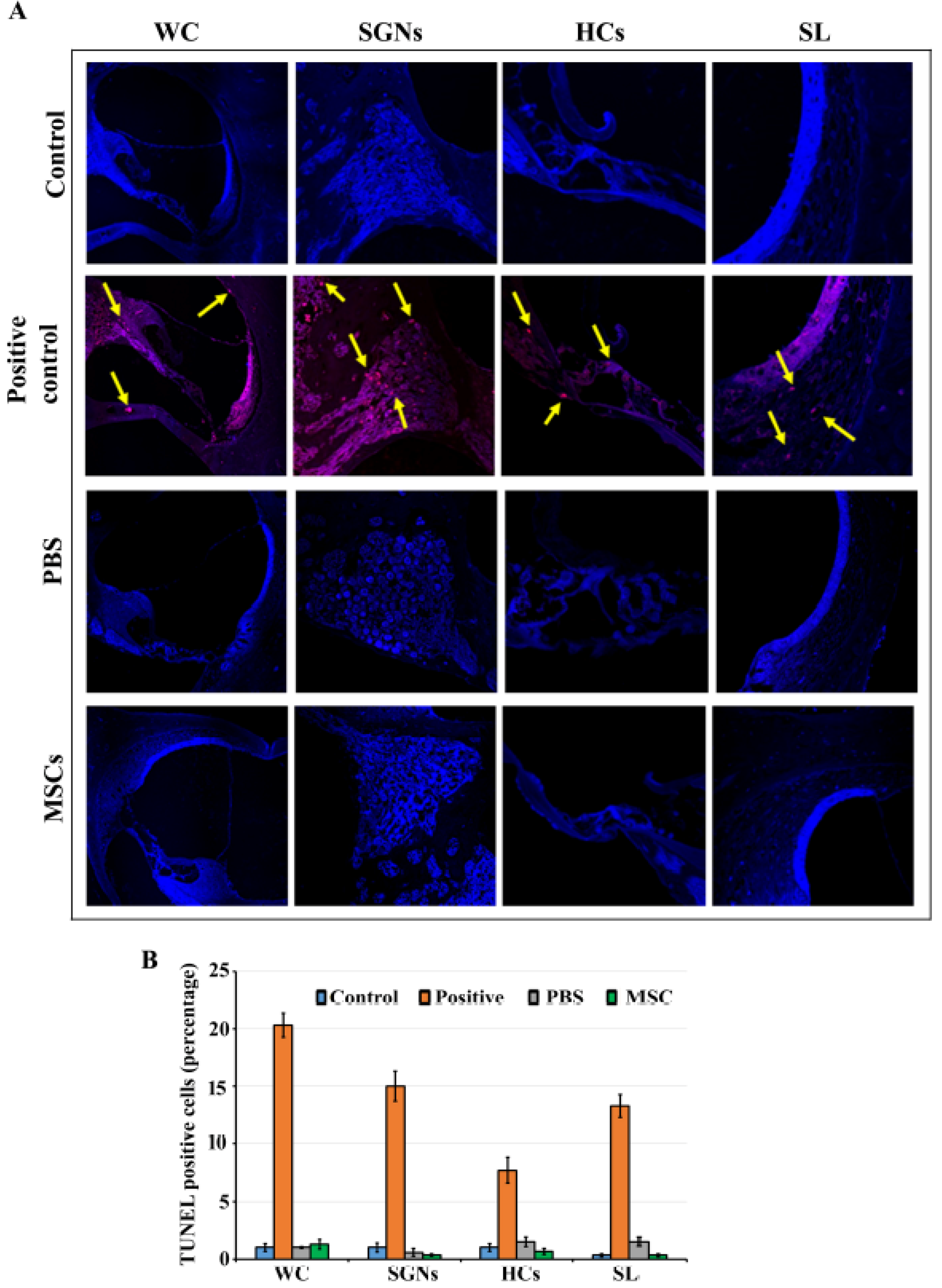
2.7. TUNEL Staining
2.8. Statistical Analysis
3. Results
3.1. Transtympanic Administration of BM-MSCs do not Induce Oxidative Stress in Rat Cochlea
3.2. Caspase 3 Pathway is not Activated in Rat Cochlea in Response to BM-MSC Administration
3.3. BM-MSCs did not Trigger Proinflammatory Cytokine Production in rat Cochlea
3.4. BM-MSCs did not Induce Cell Death in Rat Cochlea
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Graydon, K.; Waterworth, C.; Miller, H.; Gunasekera, H. Global burden of hearing impairment and ear disease. J. Laryngol. Otol. 2018, 133, 18–25. [Google Scholar] [CrossRef] [Green Version]
- Sheffield, A.M.; Smith, B.J. The Epidemiology of Deafness. Cold Spring Harb. Perspect. Med. 2018, 9, a033258. [Google Scholar] [CrossRef] [PubMed]
- Blazer, D.G. Hearing Loss. Psychiatr. Clin. North Am. 2018, 41, 19–27. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Puel, J.-L. Presbycusis: An Update on Cochlear Mechanisms and Therapies. J. Clin. Med. 2020, 9, 218. [Google Scholar] [CrossRef] [Green Version]
- Ding, T.; Yan, A.; Liu, K. What is noise-induced hearing loss? Br. J. Hosp. Med. 2019, 80, 525–529. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elliott, S.J.; Ni, G. An elemental approach to modelling the mechanics of the cochlea. Hear. Res. 2017, 360, 14–24. [Google Scholar] [CrossRef] [PubMed]
- Homans, N.C.; Metselaar, R.M.; Dingemanse, G.; van der Schroeff, M.P.; Brocaar, M.P.; Wieringa, M.H.; de Jong, R.J.B.; Hofman, A.; Goedegebure, A. Prevalence of age-related hearing loss, including sex differences, in older adults in a large cohort study. Laryngoscope 2016, 127, 725–730. [Google Scholar] [CrossRef]
- Wattamwar, K.; Qian, Z.J.; Otter, J.; Leskowitz, M.J.; Caruana, F.F.; Siedlecki, B.; Spitzer, J.B.; Lalwani, A.K. Increases in the Rate of Age-Related Hearing Loss in the Older Old. JAMA Otolaryngol. Neck Surg. 2017, 143, 41. [Google Scholar] [CrossRef]
- Ferguson, M.; Kitterick, P.T.; Chong, L.Y.; Edmondson-Jones, M.; Barker, F.; Hoare, D.J. Hearing aids for mild to moderate hearing loss in adults. Cochrane Database Syst. Rev. 2017. [Google Scholar] [CrossRef]
- Orji, A.; Kamenov, K.; Dirac, M.; Davis, A.; Chadha, S.; Vos, T. Global and regional needs, unmet needs, and access to hearing aids. Int. J. Audiol. 2020, 59, 166–172. [Google Scholar] [CrossRef]
- Eshraghi, A.; Nazarian, R.; Telischi, F.F.; Rajguru, S.M.; Truy, E.; Gupta, C. The cochlear implant: Historical aspects and prospects. Anat. Rec. Adv. Integr. Anat. Evol. Boil. 2012, 295, 1967–1980. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lesica, N.A. Why Do Hearing Aids Fail to Restore Normal Auditory Perception? Trends Neurosci. 2018, 41, 174–185. [Google Scholar] [CrossRef] [PubMed]
- McCormack, A.; Fortnum, H. Why do people fitted with hearing aids not wear them? Int. J. Audiol. 2013, 52, 360–368. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qiu, Y.; Qiu, J. Stem Cells: A New Hope for Hearing Loss Therapy. In Single Molecule and Single Cell Sequencing; Springer Science and Business Media LLC: Berlin/Heidelberg, Germany, 2019; pp. 165–180. [Google Scholar]
- Ma, Y.; Wise, A.K.; Shepherd, R.K.; Richardson, R.T. New molecular therapies for the treatment of hearing loss. Pharmacol. Ther. 2019, 200, 190–209. [Google Scholar] [CrossRef]
- Wang, J.; Puel, J.-L. Toward Cochlear Therapies. Physiol. Rev. 2018, 98, 2477–2522. [Google Scholar] [CrossRef]
- Xia, M.; Ma, J.; Sun, S.; Li, W.; Li, H. The biological strategies for hearing re-establishment based on the stem/progenitor cells. Neurosci. Lett. 2019, 711, 134406. [Google Scholar] [CrossRef]
- Mittal, R.; Nguyen, D.; Patel, A.P.; Debs, L.H.; Mittal, J.; Yan, D.; Eshraghi, A.A.; van de Water, T.R.; Liu, X.Z. Recent Advancements in the Regeneration of Auditory Hair Cells and Hearing Restoration. Front. Mol. Neurosci. 2017, 10, 236. [Google Scholar] [CrossRef] [Green Version]
- Gunewardene, N.; Crombie, D.; Dottori, M.; Nayagam, B.A. Innervation of Cochlear Hair Cells by Human Induced Pluripotent Stem Cell-Derived Neurons In Vitro. Stem Cells Int. 2016, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Takeda, H.; Hosoya, M.; Fujioka, M.; Saegusa, C.; Saeki, T.; Miwa, T.; Okano, H.; Minoda, R. Engraftment of Human Pluripotent Stem Cell-derived Progenitors in the Inner Ear of Prenatal Mice. Sci. Rep. 2018, 8, 1941. [Google Scholar] [CrossRef] [Green Version]
- Tang, Z.-H.; Chen, J.-R.; Zheng, J.; Shi, H.-S.; Ding, J.; Qian, X.-D.; Zhang, C.; Chen, J.-L.; Wang, C.-C.; Li, L.; et al. Genetic Correction of Induced Pluripotent Stem Cells From a Deaf Patient WithMYO7AMutation Results in Morphologic and Functional Recovery of the Derived Hair Cell-Like Cells. Stem Cells Transl. Med. 2016, 5, 561–571. [Google Scholar] [CrossRef] [Green Version]
- Ullah, I.; Subbarao, R.B.; Rho, G.J. Human mesenchymal stem cells current trends and future prospective. Biosci. Rep. 2015, 35, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Murphy, M.B.; Moncivais, K.; Caplan, A. Mesenchymal stem cells: Environmentally responsive therapeutics for regenerative medicine. Exp. Mol. Med. 2013, 45, e54. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Miguel, M.P.; Fuentes-Julian, S.; Blazquez-Martinez, A.; Pascual, C.Y.; Aller, M.Á.; Arias, J.; Arnalich-Montiel, F. Immunosuppressive properties of mesenchymal stem cells: Advances and applications. Curr. Mol. Med. 2012, 12, 574–591. [Google Scholar] [CrossRef] [PubMed]
- Chiu, C.-H.; Chang, T.-H.; Chang, S.-S.; Chang, G.-J.; Chen, A.C.-Y.; Cheng, C.-Y.; Chen, S.-C.; Fu, J.-F.; Wen, C.-J.; Chan, Y.-S. Application of Bone Marrow–Derived Mesenchymal Stem Cells for Muscle Healing After Contusion Injury in Mice. Am. J. Sports Med. 2020, 48, 1226–1235. [Google Scholar] [CrossRef]
- Afshari, A.; Shamdani, S.; Uzan, G.; Naserian, S.; Azarpira, N. Different approaches for transformation of mesenchymal stem cells into hepatocyte-like cells. Stem Cell Res. Ther. 2020, 11, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Jiang, Y.; Jahagirdar, B.N.; Reinhardt, R.L.; Schwartz, R.E.; Keene, C.D.; Ortiz-Gonzalez, X.; Reyes, M.; Lenvik, T.; Lund, T.; Blackstad, M.; et al. Pluripotency of mesenchymal stem cells derived from adult marrow. Nature 2002, 418, 41–49. [Google Scholar] [CrossRef] [Green Version]
- Cselenyák, A.; Pankotai, E.; Horvath, E.M.; Kiss, L.; Lacza, Z. Mesenchymal stem cells rescue cardiomyoblasts from cell death in an in vitro ischemia model via direct cell-to-cell connections. BMC Cell Boil. 2010, 11, 29. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.-Y.; Lee, J.-H.; Kim, H.J.; Park, M.K.; Huh, J.W.; Ro, J.Y.; Oh, Y.-M.; Lee, S.-D.; Lee, Y.-S. Mesenchymal stem cell-conditioned media recovers lung fibroblasts from cigarette smoke-induced damage. Am. J. Physiol. Cell. Mol. Physiol. 2012, 302, 891–908. [Google Scholar] [CrossRef] [Green Version]
- Li, N.; Sarojini, H.; An, J.; Wang, E. Prosaposin in the secretome of marrow stroma-derived neural progenitor cells protects neural cells from apoptotic death. J. Neurochem. 2010, 112, 1527–1538. [Google Scholar] [CrossRef]
- Tanyeli, T.T.; Karadas, H.; Akyildiz, I.; Gokdogan, O.; Sonmez, C.; Cavus, M.E.; Kaptan, Z.; Uzunkulaoglu, H.; Arslan, N.; Zeybek, N.D. Effect of Folic Acid on Cisplatin-Induced Ototoxicity: A Functional and Morphological Study. J. Int. Adv. Otol. 2019, 15, 237–246. [Google Scholar] [CrossRef]
- Simsek, G.; Taş, B.M.; Muluk, N.B.; Azman, M.; Kılıç, R. Comparison of the protective efficacy between intratympanic dexamethasone and resveratrol treatments against cisplatin-induced ototoxicity: An experimental study. Eur. Arch. Otorhinolaryngol. 2019, 276, 3287–3293. [Google Scholar] [CrossRef] [PubMed]
- Eren, S.B.; Doğan, R.; Yenigun, A.; Veyseller, B.; Tugrul, S.; Ozturan, O.; Aydin, M.S. Evaluation of ototoxicity of intratympanic administration of Methotrexate in rats. Int. J. Pediatr. Otorhinolaryngol. 2017, 100, 132–136. [Google Scholar] [CrossRef] [PubMed]
- Göl, A.; Şengül, E.; Yılmaz, B.; Özkurt, F.E.; Akdağ, M.; Keleş, A.; Topçu, I. The Protective Effect of Intratympanic Dexamethasone on Streptomycin Ototoxicity in Rats. Ear Nose Throat J. 2017, 96, 12–17. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.; Chan, C. Isolation and Enrichment of Rat Mesenchymal Stem Cells (MSCs) and Separation of Single-colony Derived MSCs. J. Vis. Exp. 2010, 37. [Google Scholar] [CrossRef] [PubMed]
- Dominici, M.; le Blanc, K.; Mueller, I.; Slaper-Cortenbach, I.; Marini, F.; Krause, D.; Deans, R.; Keating, A.; Prockop, D.; Horwitz, E. Minimal criteria for defining multipotent mesenchymal stromal cells. The International Society for Cellular Therapy position statement. Cytotherapy 2006, 8, 315–317. [Google Scholar] [CrossRef]
- Elfsmark, L.; Agren, L.; Akfur, C.; Bucht, A.; Jonasson, S. 8-Isoprostane is an early biomarker for oxidative stress in chlorine-induced acute lung injury. Toxicol. Lett. 2018, 282, 1–7. [Google Scholar] [CrossRef]
- Mistry, R.J.; Klamt, F.; Ramsden, D.B.; Parsons, R.B. Nicotinamide N -methyltransferase expression in SH-SY5Y human neuroblastoma cells decreases oxidative stress. J. Biochem. Mol. Toxicol. 2020, 34, e22439. [Google Scholar] [CrossRef]
- Cabello-Verrugio, C.; Simon, F.; Trollet, C.; Santibanez, J. Oxidative Stress in Disease and Aging: Mechanisms and Therapies 2016. Oxidative Med. Cell. Longev. 2017, 1–2. [Google Scholar] [CrossRef]
- Sies, H. Oxidative stress: A concept in redox biology and medicine. Redox Boil. 2015, 4, 180–183. [Google Scholar] [CrossRef] [Green Version]
- Valle-Prieto, A.; Conget, P. Human Mesenchymal Stem Cells Efficiently Manage Oxidative Stress. Stem Cells Dev. 2010, 19, 1885–1893. [Google Scholar] [CrossRef]
- Silva, W.A.; Covas, D.T.; Panepucci, R.A.; Proto-Siqueira, R.; Siufi, J.L.; Zanette, D.L.; Santos, A.R.; Zago, M.A. The Profile of Gene Expression of Human Marrow Mesenchymal Stem Cells. Stem Cells 2003, 21, 661–669. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Motegi, S.-I.; Sekiguchi, A.; Uchiyama, A.; Uehara, A.; Fujiwara, C.; Yamazaki, S.; Perera, B.; Nakamura, H.; Ogino, S.; Yokoyama, Y.; et al. Protective effect of mesenchymal stem cells on the pressure ulcer formation by the regulation of oxidative and endoplasmic reticulum stress. Sci. Rep. 2017, 7, 17186. [Google Scholar] [CrossRef] [PubMed]
- Deng, L.; Liu, G.; Wu, X.; Wang, Y.; Tong, M.; Liu, B.; Wang, K.; Peng, Y.; Kong, X. Adipose Derived Mesenchymal Stem Cells Efficiently Rescue Carbon Tetrachloride-Induced Acute Liver Failure in Mouse. Sci. World J. 2014, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ciorba, A.; Gasparini, P.; Chicca, M.; Pinamonti, S.; Martini, A. Reactive oxygen species in human inner ear perilymph. Acta OtoLaryngol. 2010, 130, 240–246. [Google Scholar] [CrossRef] [PubMed]
- Ohlemiller, K.K.; Wright, J.S.; Dugan, L.L. Early Elevation of Cochlear Reactive Oxygen Species following Noise Exposure. Audiol. Neurotol. 1999, 4, 229–236. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Liu, Y.; Han, N.; Chen, X.; Yu, W.; Zhang, W.; Zou, F. Profiles of oxidative stress-related microRNA and mRNA expression in auditory cells. Brain Res. 2010, 1346, 14–25. [Google Scholar] [CrossRef] [PubMed]
- Yamane, H.; Nakai, Y.; Takayama, M.; Iguchi, H.; Nakagawa, T.; Kojima, A. Appearance of free radicals in the guinea pig inner ear after noise-induced acoustic trauma. Eur. Arch. Otorhinolaryngol. 1995, 252, 504–508. [Google Scholar] [CrossRef]
- Shi, X.; Nuttall, A. Upregulated iNOS and oxidative damage to the cochlear stria vascularis due to noise stress. Brain Res. 2003, 967, 1–10. [Google Scholar] [CrossRef]
- van Campen, L.; Murphy, W.J.; Franks, J.R.; I Mathias, P.; A Toraason, M. Oxidative DNA damage is associated with intense noise exposure in the rat. Hear. Res. 2002, 164, 29–38. [Google Scholar] [CrossRef]
- Kim, H.-J.; Lee, J.H.; Kim, S.-J.; Oh, G.S.; Moon, H.-D.; Kwon, K.-B.; Park, C.; Park, B.H.; Lee, H.-K.; Chung, S.-Y.; et al. Roles of NADPH Oxidases in Cisplatin-Induced Reactive Oxygen Species Generation and Ototoxicity. J. Neurosci. 2010, 30, 3933–3946. [Google Scholar] [CrossRef] [Green Version]
- Kovacic, P.; Somanathan, R. Ototoxicity and noise trauma: Electron transfer, reactive oxygen species, cell signaling, electrical effects, and protection by antioxidants: Practical medical aspects. Med Hypotheses 2008, 70, 914–923. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.E.; Nakagawa, T.; Kim, T.-S.; Endo, T.; Shiga, A.; Iguchi, F.; Lee, S.H.; Ito, J. Role of reactive radicals in degeneration of the auditory system of mice following cisplatin treatment. Acta Otolaryngol. 2004, 124, 1131–1135. [Google Scholar] [CrossRef] [PubMed]
- Benkafadar, N.; François, C.; Affortit, C.; Casas, F.; Ceccato, J.-C.; Menardo, J.; Venail, F.; Malfroy-Camine, B.; Puel, J.-L.; Wang, J. ROS-Induced Activation of DNA Damage Responses Drives Senescence-Like State in Postmitotic Cochlear Cells: Implication for Hearing Preservation. Mol. Neurobiol. 2019, 56, 5950–5969. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fetoni, A.R.; Eramo, S.L.M.; di Pino, A.; Rolesi, R.; Paciello, F.; Grassi, C.; Troiani, D.; Troiani, G.P. The Antioxidant Effect of Rosmarinic Acid by Different Delivery Routes in the Animal Model of Noise-Induced Hearing Loss. Otol. Neurotol. 2018, 39, 378–386. [Google Scholar] [CrossRef]
- Ohlemiller, K.K.; McFadden, S.L.; Ding, D.-L.; Lear, P.M.; Ho, Y.-S. Targeted Mutation of the Gene for Cellular Glutathione Peroxidase (Gpx1) Increases Noise-Induced Hearing Loss in Mice. J. Assoc. Res. Otolaryngol. 2000, 1, 243–254. [Google Scholar] [CrossRef] [Green Version]
- Masuda, M.; Kanzaki, S.; Minami, S.; Kikuchi, J.; Kanzaki, J.; Sato, H.; Ogawa, K. Correlations of Inflammatory Biomarkers with the Onset and Prognosis of Idiopathic Sudden Sensorineural Hearing Loss. Otol. Neurotol. 2012, 33, 1142–1150. [Google Scholar] [CrossRef] [Green Version]
- He, W.; Yu, J.; Sun, Y.; Kong, W. Macrophages in Noise-Exposed Cochlea: Changes, Regulation, and the Potential Role. Aging Dis. 2020, 11, 191–199. [Google Scholar] [CrossRef] [Green Version]
- Hirose, K.; Discolo, C.M.; Keasler, J.R.; Ransohoff, R. Mononuclear phagocytes migrate into the murine cochlea after acoustic trauma. J. Comp. Neurol. 2005, 489, 180–194. [Google Scholar] [CrossRef]
- Fujioka, M.; Kanzaki, S.; Okano, H.J.; Masuda, M.; Ogawa, K.; Okano, H. Proinflammatory cytokines expression in noise-induced damaged cochlea. J. Neurosci. Res. 2006, 83, 575–583. [Google Scholar] [CrossRef] [Green Version]
- Shin, S.-A.; Lyu, A.-R.; Jeong, S.-H.; Kim, T.; Park, M.J.; Park, Y.-H. Acoustic Trauma Modulates Cochlear Blood Flow and Vasoactive Factors in a Rodent Model of Noise-Induced Hearing Loss. Int. J. Mol. Sci. 2019, 20, 5316. [Google Scholar] [CrossRef] [Green Version]
- Hsu, H.; Xiong, J.; Goeddel, D.V. The TNF receptor 1-associated protein TRADD signals cell death and NF-κB activation. Cell 1995, 81, 495–504. [Google Scholar] [CrossRef] [Green Version]
- Satoh, H.; Firestein, G.S.; Billings, P.; Harris, J.P.; Keithley, E.M. Tumor Necrosis Factor-? An Initiator, and Etanercept, an Inhibitor of Cochlear Inflammation. Laryngoscope 2002, 112, 1627–1634. [Google Scholar] [CrossRef] [PubMed]
- Satoh, H.; Firestein, G.S.; Billings, P.; Harris, J.P.; Keithley, E.M. Proinflammatory cytokine expression in the endolymphatic sac during inner ear inflammation. J. Assoc. Res. Otolaryngol. 2003, 4, 139–147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsujimoto, M.; Yokota, S.; Vilcek, J.; Weissmann, G. Tumor necrosis factor provokes superoxide anion generation from neutrophils. Biochem. Biophys. Res. Commun. 1986, 137, 1094–1100. [Google Scholar] [CrossRef]
- Woo, C.-H.; Eom, Y.-W.; Yoo, M.-H.; You, H.-J.; Han, H.J.; Song, W.K.; Yoo, Y.J.; Chun, J.-S.; Kim, J.-H. Tumor Necrosis Factor-α Generates Reactive Oxygen Species via a Cytosolic Phospholipase A2-linked Cascade. J. Boil. Chem. 2000, 275, 32357–32362. [Google Scholar] [CrossRef] [Green Version]
- Zou, J.; Pyykkö, I.; Sutinen, P.; Toppila, E. Vibration induced hearing loss in guinea pig cochlea: Expression of TNF-α and VEGF. Hear. Res. 2005, 202, 13–20. [Google Scholar] [CrossRef]
- Dinh, C.T.; Goncalves, S.; Bas, E.; van de Water, T.R.; Zine, A. Molecular regulation of auditory hair cell death and approaches to protect sensory receptor cells and/or stimulate repair following acoustic trauma. Front. Cell. Neurosci. 2015, 9. [Google Scholar] [CrossRef] [Green Version]
- Cheng, A.G.; Cunningham, L.; Rubel, E.W. Mechanisms of hair cell death and protection. Curr. Opin. Otolaryngol. Head Neck Surg. 2005, 13, 343–348. [Google Scholar] [CrossRef]
- Porter, A.G.; Jänicke, R.U. Emerging roles of caspase-3 in apoptosis. Cell Death Differ. 1999, 6, 99–104. [Google Scholar] [CrossRef]
- Łos, M.J.; Wesselborg, S.; Schulze-Osthoff, K. The Role of Caspases in Development, Immunity, and Apoptotic Signal Transduction. Immunity 1999, 10, 629–639. [Google Scholar] [CrossRef] [Green Version]
- Enari, M.; Sakahira, H.; Yokoyama, H.; Okawa, K.; Iwamatsu, A.; Nagata, S. A caspase-activated DNase that degrades DNA during apoptosis, and its inhibitor ICAD. Nature 1998, 391, 43–50. [Google Scholar] [CrossRef]
- Wolf, B.B.; Schuler, M.; Echeverri, F.; Green, D.R. Caspase-3 Is the Primary Activator of Apoptotic DNA Fragmentation via DNA Fragmentation Factor-45/Inhibitor of Caspase-activated DNase Inactivation. J. Boil. Chem. 1999, 274, 30651–30656. [Google Scholar] [CrossRef] [Green Version]
- Kovacsovics, M.; Martinon, F.; Micheau, O.; Bodmer, J.L.; Hofmann, K.; Tschopp, J. Overexpression of Helicard, a CARD-Containing Helicase Cleaved during Apoptosis, Accelerates DNA Degradation. Curr. Boil. 2002, 12, 838–843. [Google Scholar] [CrossRef] [Green Version]
- van de Water, T.R.; Lallemend, F.; Eshraghi, A.A.; Ahsan, S.; He, J.; Guzman, J.; Polak, M.; Malgrange, B.; Lefebvre, P.P.; Staecker, H.; et al. Caspases, the Enemy Within, and Their Role in Oxidative Stress-Induced Apoptosis of Inner Ear Sensory Cells. Otol. Neurotol. 2004, 25, 627–632. [Google Scholar] [CrossRef]
- Okuda, T.; Sugahara, K.; Takemoto, T.; Shimogori, H.; Yamashita, H. Inhibition of caspases alleviates gentamicin-induced cochlear damage in guinea pigs. Auris Nasus Larynx 2005, 32, 33–37. [Google Scholar] [CrossRef]
- Sheth, S.; Mukherjea, D.; Rybak, L.P.; Ramkumar, V. Mechanisms of Cisplatin-Induced Ototoxicity and Otoprotection. Front. Cell. Neurosci. 2017, 11, 338. [Google Scholar] [CrossRef]
- Kabakov, A.; Gabai, V. Cell Death and Survival Assays; Springer Science and Business Media LLC: Berlin/Heidelberg, Germany, 2017; pp. 107–127. [Google Scholar]
- Kyrylkova, K.; Kyryachenko, S.; Leid, M.; Kioussi, C. Detection of Apoptosis by TUNEL Assay. Adv. Struct. Saf. Stud. 2012, 887, 41–47. [Google Scholar] [CrossRef]
- Zhu, G.; Shen, Q.; Jiang, H.; Ji, O.; Zhu, L.; Zhang, L. Curcumin inhibited the growth and invasion of human monocytic leukaemia SHI-1 cells in vivo by altering MAPK and MMP signalling. Pharm. Biol. 2020, 58, 25–34. [Google Scholar] [CrossRef] [Green Version]
- Kamiya, K.; Fujinami, Y.; Hoya, N.; Okamoto, Y.; Kouike, H.; Komatsuzaki, R.; Kusano, R.; Nakagawa, S.; Satoh, H.; Fujii, M.; et al. Mesenchymal Stem Cell Transplantation Accelerates Hearing Recovery through the Repair of Injured Cochlear Fibrocytes. Am. J. Pathol. 2007, 171, 214–226. [Google Scholar] [CrossRef] [Green Version]
- Lang, H.; Schulte, B.A.; Goddard, J.C.; Hedrick, M.; Schulte, J.B.; Wei, L.; Schmiedt, R.A. Transplantation of Mouse Embryonic Stem Cells into the Cochlea of an Auditory-Neuropathy Animal Model: Effects of Timing after Injury. J. Assoc. Res. Otolaryngol. 2008, 9, 225–240. [Google Scholar] [CrossRef] [Green Version]
- Nacher-Soler, G.; Garrido, J.M.; Rodríguez-Serrano, F. Hearing regeneration and regenerative medicine: Present and future approaches. Arch. Med. Sci. 2019, 15, 957–967. [Google Scholar] [CrossRef]
- Maacha, S.; Sidahmed, H.; Jacob, S.; Gentilcore, G.; Calzone, R.; Grivel, J.-C.; Cugno, C. Paracrine Mechanisms of Mesenchymal Stromal Cells in Angiogenesis. Stem Cells Int. 2020. [Google Scholar] [CrossRef]
- Fan, X.-L.; Zhang, Y.; Li, X.; Fu, Q.-L. Mechanisms underlying the protective effects of mesenchymal stem cell-based therapy. Cell. Mol. Life Sci. 2020, 1–4. [Google Scholar] [CrossRef] [Green Version]
- Linero, I.; Chaparro, O. Paracrine Effect of Mesenchymal Stem Cells Derived from Human Adipose Tissue in Bone Regeneration. PLoS ONE 2014, 9, e107001. [Google Scholar] [CrossRef] [Green Version]
- Ankrum, J.; Ong, J.F.; Karp, J.M. Mesenchymal stem cells: Immune evasive, not immune privileged. Nat. Biotechnol. 2014, 32, 252–260. [Google Scholar] [CrossRef] [Green Version]
- Kim, K.H.; Jo, J.H.; Cho, H.J.; Park, T.S.; Kim, T.M. Therapeutic potential of stem cell-derived extracellular vesicles in osteoarthritis: Preclinical study findings. Lab. Anim. Res. 2020, 36, 10–17. [Google Scholar] [CrossRef] [Green Version]
- Wang, C.; Börger, V.; Sardari, M.; Murke, F.; Skuljec, J.; Pul, R.; Hagemann, N.; Dzyubenko, E.; Dittrich, R.; Gregorius, J.; et al. Mesenchymal Stromal Cell–Derived Small Extracellular Vesicles Induce Ischemic Neuroprotection by Modulating Leukocytes and Specifically Neutrophils. Stroke 2020, 51, 1825–1834. [Google Scholar] [CrossRef]
- Huang, C.-C.; Kang, M.; Lu, Y.; Shirazi, S.; Diaz, J.I.; Cooper, L.F.; Gajendrareddy, P.; Ravindran, S. Functionally Engineered Extracellular Vesicles Improve Bone Regeneration. Acta Biomater 2020. [Google Scholar] [CrossRef]
- Vizoso, F.; Eiro, N.; Cid, S.; Schneider, J.; Perez-Fernandez, R. Mesenchymal Stem Cell Secretome: Toward Cell-Free Therapeutic Strategies in Regenerative Medicine. Int. J. Mol. Sci. 2017, 18, 1852. [Google Scholar] [CrossRef] [Green Version]
- Carling, G.; Greenwald, J.; Sadiq, S.; Harris, V. Paracrine Modulation of Microglial Activation and Oligodendroglial Differentiation as Potential Therapeutic Mechanisms of Mesenchymal Stem Cell-Derived Neural Progenitors (MSC-NPs) Cellular Therapy in MS (P2.2-059). Neurology 2019, 92. [Google Scholar]
- Yoo, T.; Du, X.; Zhou, B. The paracrine effect of mesenchymal human stem cells restored hearing in β-tubulin induced autoimmune sensorineural hearing loss. Hear. Res. 2015, 330, 57–61. [Google Scholar] [CrossRef] [PubMed]
- Liang, X.; Ding, Y.; Zhang, Y.; Tse, H.-F.; Lian, Q. Paracrine Mechanisms of Mesenchymal Stem Cell-Based Therapy: Current Status and Perspectives. Cell Transplant. 2014, 23, 1045–1059. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choi, M.Y.; Yeo, S.W.; Park, K.H. Hearing restoration in a deaf animal model with intravenous transplantation of mesenchymal stem cells derived from human umbilical cord blood. Biochem. Biophys. Res. Commun. 2012, 427, 629–636. [Google Scholar] [CrossRef]
- Ma, Y.; Guo, W.; Yi, H.; Ren, L.; Zhao, L.; Zhang, Y.; Yuan, S.; Liu, R.; Xu, L.; Cong, T.; et al. Transplantation of human umbilical cord mesenchymal stem cells in cochlea to repair sensorineural hearing. Am. J. Transl. Res. 2016, 8, 5235–5245. [Google Scholar]
- Patel, J.; Szczupak, M.; Rajguru, S.; Balaban, C.; Hoffer, M.E. Inner Ear Therapeutics: An Overview of Middle Ear Delivery. Front. Cell. Neurosci. 2019, 13, 261. [Google Scholar] [CrossRef] [Green Version]




© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Eshraghi, A.A.; Ocak, E.; Zhu, A.; Mittal, J.; Davies, C.; Shahal, D.; Bulut, E.; Sinha, R.; Shah, V.; Perdomo, M.M.; et al. Biocompatibility of Bone Marrow-Derived Mesenchymal Stem Cells in the Rat Inner Ear following Trans-Tympanic Administration. J. Clin. Med. 2020, 9, 1711. https://doi.org/10.3390/jcm9061711
Eshraghi AA, Ocak E, Zhu A, Mittal J, Davies C, Shahal D, Bulut E, Sinha R, Shah V, Perdomo MM, et al. Biocompatibility of Bone Marrow-Derived Mesenchymal Stem Cells in the Rat Inner Ear following Trans-Tympanic Administration. Journal of Clinical Medicine. 2020; 9(6):1711. https://doi.org/10.3390/jcm9061711
Chicago/Turabian StyleEshraghi, Adrien A., Emre Ocak, Angela Zhu, Jeenu Mittal, Camron Davies, David Shahal, Erdogan Bulut, Rahul Sinha, Viraj Shah, Mario M. Perdomo, and et al. 2020. "Biocompatibility of Bone Marrow-Derived Mesenchymal Stem Cells in the Rat Inner Ear following Trans-Tympanic Administration" Journal of Clinical Medicine 9, no. 6: 1711. https://doi.org/10.3390/jcm9061711
APA StyleEshraghi, A. A., Ocak, E., Zhu, A., Mittal, J., Davies, C., Shahal, D., Bulut, E., Sinha, R., Shah, V., Perdomo, M. M., & Mittal, R. (2020). Biocompatibility of Bone Marrow-Derived Mesenchymal Stem Cells in the Rat Inner Ear following Trans-Tympanic Administration. Journal of Clinical Medicine, 9(6), 1711. https://doi.org/10.3390/jcm9061711