Palmitic Acid Targets Human Testicular Peritubular Cells and Causes a Pro-Inflammatory Response

 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Human Samples: HTPCs
2.2. Samples from Normal and Obese Monkeys
2.3. Stimulation Experiments with Palmitic Acid (PA)
2.4. Isolation of RNA, Reverse Transcription (RT-PCR) and Quantitative Real Time PCR (qPCR)
2.5. IL6 ELISA Measurements
2.6. Statistical Analysis
3. Results
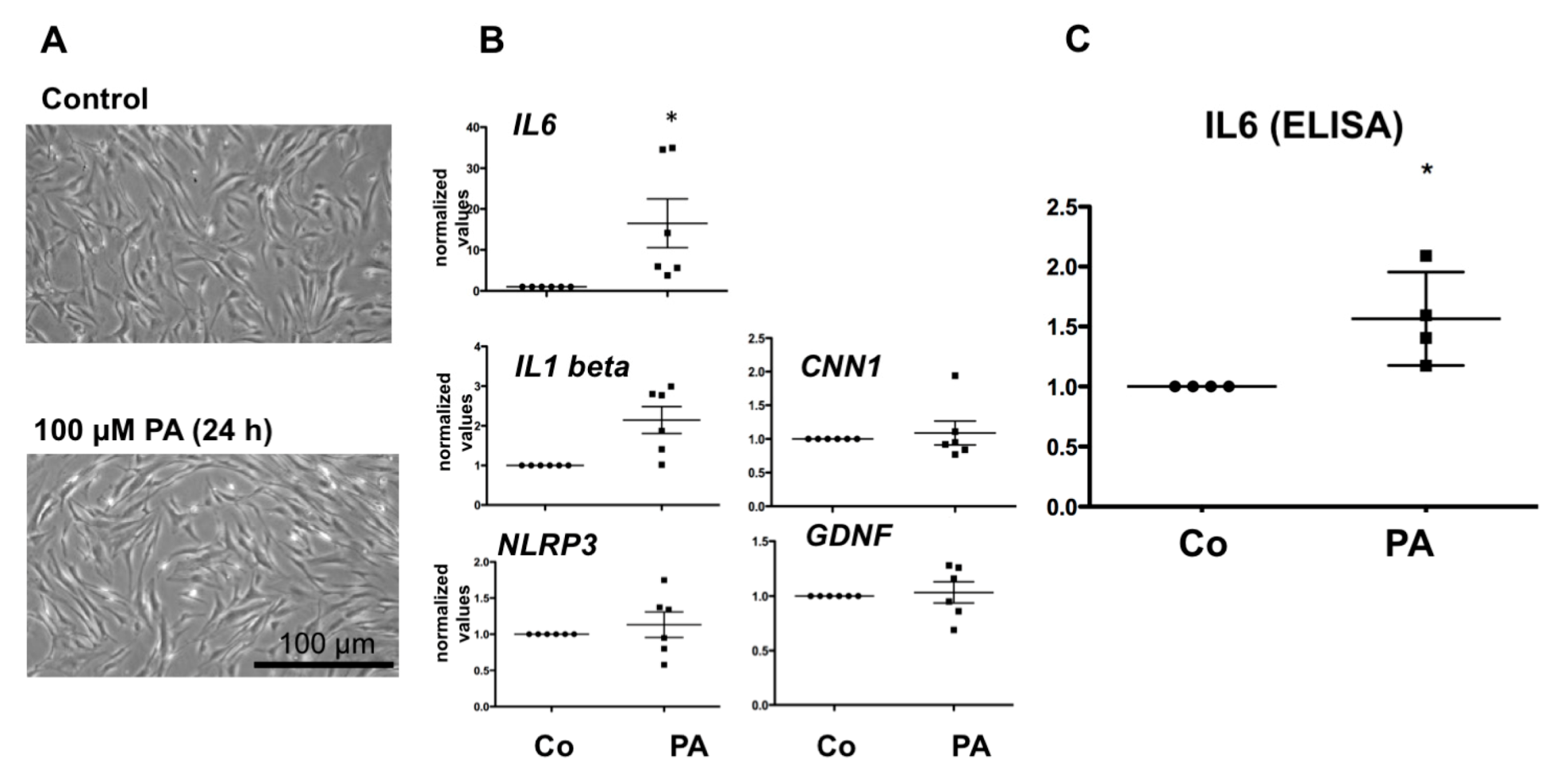
3.1. Effect of PA on HTPC mRNA Level of IL6, NLRP3, IL1beta and CNN1
3.2. Effect of PA on IL6 Secretion in HTPCs
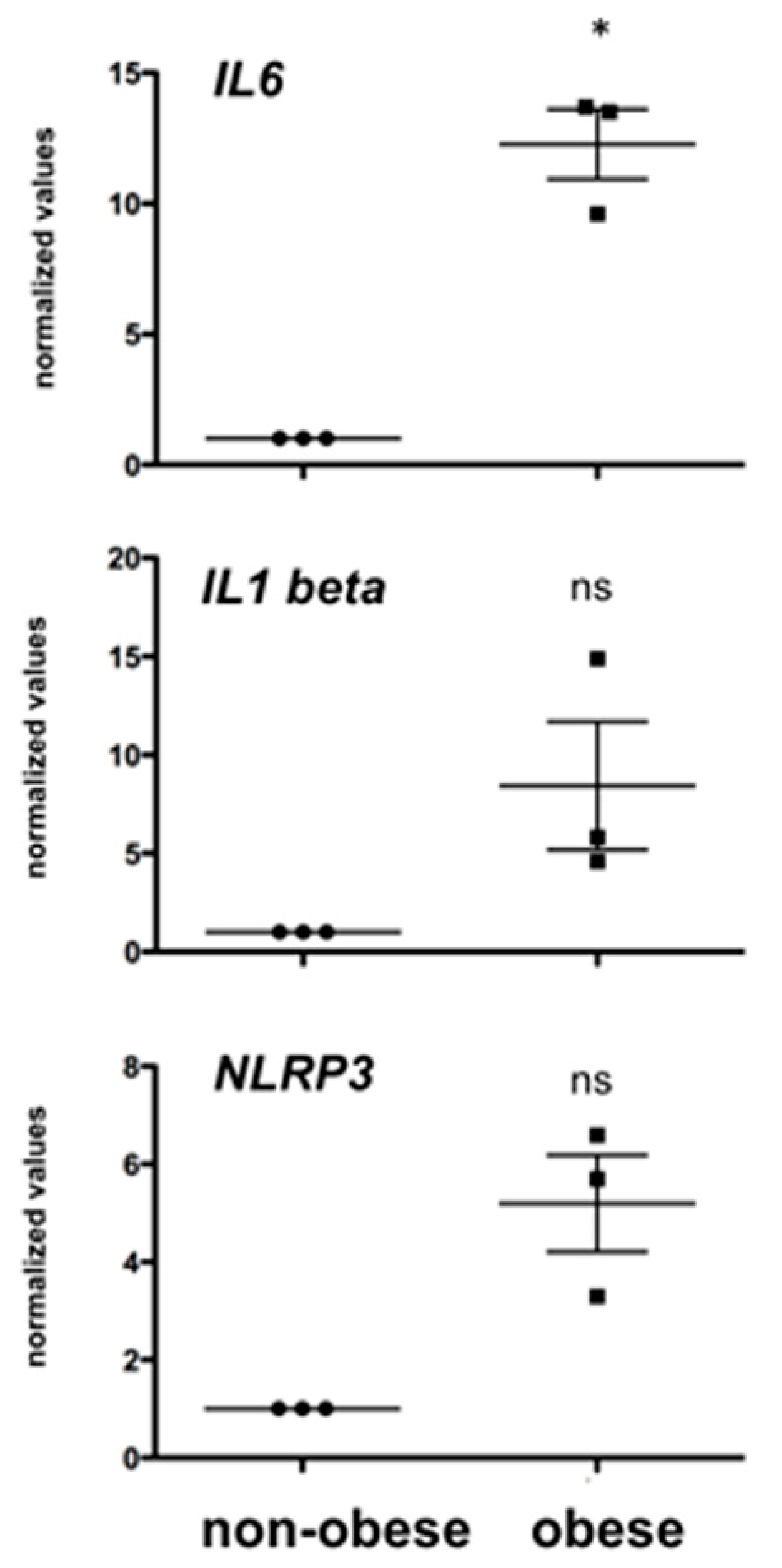
3.3. Obesity and Testicular IL6 Levels in Rhesus Monkeys
4. Discussion
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Mayer, C.; Adam, M.; Glashauser, L.; Dietrich, K.; Schwarzer, J.U.; Kohn, F.M.; Strauss, L.; Welter, H.; Poutanen, M.; Mayerhofer, A. Sterile inflammation as a factor in human male infertility: Involvement of Toll like receptor 2, biglycan and peritubular cells. Sci. Rep. 2016, 6, 37128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walenta, L.; Schmid, N.; Schwarzer, J.U.; Kohn, F.M.; Urbanski, H.F.; Behr, R.; Strauss, L.; Poutanen, M.; Mayerhofer, A. NLRP3 in somatic non-immune cells of rodent and primate testes. Reproduction 2018, 156, 231–238. [Google Scholar] [CrossRef] [PubMed]
- Mayerhofer, A. Human testicular peritubular cells: More than meets the eye. Reproduction 2013, 145, R107–R116. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maekawa, M.; Kamimura, K.; Nagano, T. Peritubular myoid cells in the testis: Their structure and function. Arch. Histol. Cytol. 1996, 59, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Welter, H.; Kohn, F.M.; Mayerhofer, A. Mast cells in human testicular biopsies from patients with mixed atrophy: Increased numbers, heterogeneity, and expression of cyclooxygenase 2 and prostaglandin D2 synthase. Fertil. Steril. 2011, 96, 309–313. [Google Scholar] [CrossRef]
- Welter, H.; Huber, A.; Lauf, S.; Einwang, D.; Mayer, C.; Schwarzer, J.U.; Kohn, F.M.; Mayerhofer, A. Angiotensin II regulates testicular peritubular cell function via AT1 receptor: A specific situation in male infertility. Mol. Cell. Endocrinol. 2014, 393, 171–178. [Google Scholar] [CrossRef]
- Meineke, V.; Frungieri, M.B.; Jessberger, B.; Vogt, H.; Mayerhofer, A. Human testicular mast cells contain tryptase: Increased mast cell number and altered distribution in the testes of infertile men. Fertil. Steril. 2000, 74, 239–244. [Google Scholar] [CrossRef]
- Schell, C.; Albrecht, M.; Mayer, C.; Schwarzer, J.U.; Frungieri, M.B.; Mayerhofer, A. Exploring human testicular peritubular cells: Identification of secretory products and regulation by tumor necrosis factor-alpha. Endocrinology 2008, 149, 1678–1686. [Google Scholar] [CrossRef]
- Frungieri, M.B.; Calandra, R.S.; Lustig, L.; Meineke, V.; Kohn, F.M.; Vogt, H.J.; Mayerhofer, A. Number, distribution pattern, and identification of macrophages in the testes of infertile men. Fertil. Steril. 2002, 78, 298–306. [Google Scholar] [CrossRef]
- Frungieri, M.B.; Weidinger, S.; Meineke, V.; Kohn, F.M.; Mayerhofer, A. Proliferative action of mast-cell tryptase is mediated by PAR2, COX2, prostaglandins, and PPARgamma: Possible relevance to human fibrotic disorders. Proc. Natl. Acad. Sci. USA 2002, 99, 15072–15077. [Google Scholar] [CrossRef] [Green Version]
- Flenkenthaler, F.; Windschuttl, S.; Frohlich, T.; Schwarzer, J.U.; Mayerhofer, A.; Arnold, G.J. Secretome analysis of testicular peritubular cells: A window into the human testicular microenvironment and the spermatogonial stem cell niche in man. J. Proteome Res. 2014, 13, 1259–1269. [Google Scholar] [CrossRef]
- Schmid, N.; Flenkenthaler, F.; Stockl, J.B.; Dietrich, K.G.; Kohn, F.M.; Schwarzer, J.U.; Kunz, L.; Luckner, M.; Wanner, G.; Arnold, G.J.; et al. Insights into replicative senescence of human testicular peritubular cells. Sci. Rep. 2019, 9, 15052. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walenta, L.; Fleck, D.; Frohlich, T.; von Eysmondt, H.; Arnold, G.J.; Spehr, J.; Schwarzer, J.U.; Kohn, F.M.; Spehr, M.; Mayerhofer, A. ATP-mediated Events in Peritubular Cells Contribute to Sterile Testicular Inflammation. Sci. Rep. 2018, 8, 1431. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spinnler, K.; Kohn, F.M.; Schwarzer, U.; Mayerhofer, A. Glial cell line-derived neurotrophic factor is constitutively produced by human testicular peritubular cells and may contribute to the spermatogonial stem cell niche in man. Hum. Reprod. 2010, 25, 2181–2187. [Google Scholar] [CrossRef] [PubMed]
- Eubler, K.; Herrmann, C.; Tiefenbacher, A.; Kohn, F.M.; Schwarzer, J.U.; Kunz, L.; Mayerhofer, A. Ca2+ Signaling and IL-8 Secretion in Human Testicular Peritubular Cells Involve the Cation Channel TRPV2. Int. J. Mol. Sci. 2018, 19, 2829. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johnson, A.R.; Milner, J.J.; Makowski, L. The inflammation highway: Metabolism accelerates inflammatory traffic in obesity. Immunol. Rev. 2012, 249, 218–238. [Google Scholar] [CrossRef] [Green Version]
- Korbecki, J.; Bajdak-Rusinek, K. The effect of palmitic acid on inflammatory response in macrophages: An overview of molecular mechanisms. Inflamm. Res. 2019, 68, 915–932. [Google Scholar] [CrossRef] [Green Version]
- Nemecz, M.; Constantin, A.; Dumitrescu, M.; Alexandru, N.; Filippi, A.; Tanko, G.; Georgescu, A. The Distinct Effects of Palmitic and Oleic Acid on Pancreatic Beta Cell Function: The Elucidation of Associated Mechanisms and Effector Molecules. Front. Pharmacol. 2018, 9, 1554. [Google Scholar] [CrossRef] [Green Version]
- Fatima, S.; Hu, X.; Gong, R.H.; Huang, C.; Chen, M.; Wong, H.L.X.; Bian, Z.; Kwan, H.Y. Palmitic acid is an intracellular signaling molecule involved in disease development. Cell. Mol. Life Sci. 2019, 76, 2547–2557. [Google Scholar] [CrossRef]
- Jutte, N.H.; Eikvar, L.; Levy, F.O.; Hansson, V. Metabolism of palmitate in cultured rat Sertoli cells. J. Reprod. Fertil. 1985, 73, 497–503. [Google Scholar] [CrossRef] [Green Version]
- Hu, X.; Ge, X.; Liang, W.; Shao, Y.; Jing, J.; Wang, C.; Zeng, R.; Yao, B. Effects of saturated palmitic acid and omega-3 polyunsaturated fatty acids on Sertoli cell apoptosis. Syst. Biol. Reprod. Med. 2018, 64, 368–380. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luo, D.; Zhang, M.; Su, X.; Liu, L.; Zhou, X.; Zhang, X.; Zheng, D.; Yu, C.; Guan, Q. High fat diet impairs spermatogenesis by regulating glucose and lipid metabolism in Sertoli cells. Life Sci. 2020, 257, 118028. [Google Scholar] [CrossRef] [PubMed]
- Dorfman, M.D.; Thaler, J.P. Hypothalamic inflammation and gliosis in obesity. Curr. Opin. Endocrinol. Diabetes Obes. 2015, 22, 325–330. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Osborn, O.; Olefsky, J.M. The cellular and signaling networks linking the immune system and metabolism in disease. Nat. Med. 2012, 18, 363–374. [Google Scholar] [CrossRef]
- Lainez, N.M.; Jonak, C.R.; Nair, M.G.; Ethell, I.M.; Wilson, E.H.; Carson, M.J.; Coss, D. Diet-Induced Obesity Elicits Macrophage Infiltration and Reduction in Spine Density in the Hypothalami of Male but Not Female Mice. Front. Immunol. 2018, 9, 1992. [Google Scholar] [CrossRef]
- Olefsky, J.M.; Glass, C.K. Macrophages, inflammation, and insulin resistance. Annu. Rev. Physiol. 2010, 72, 219–246. [Google Scholar] [CrossRef]
- Perez, C.V.; Sobarzo, C.M.; Jacobo, P.V.; Pellizzari, E.H.; Cigorraga, S.B.; Denduchis, B.; Lustig, L. Loss of occludin expression and impairment of blood-testis barrier permeability in rats with autoimmune orchitis: Effect of interleukin 6 on Sertoli cell tight junctions. Biol. Reprod. 2012, 87, 122. [Google Scholar] [CrossRef]
- Fan, Y.; Liu, Y.; Xue, K.; Gu, G.; Fan, W.; Xu, Y.; Ding, Z. Diet-induced obesity in male C57BL/6 mice decreases fertility as a consequence of disrupted blood-testis barrier. PLoS ONE 2015, 10, e0120775. [Google Scholar] [CrossRef]
- Liu, Y.; Ding, Z. Obesity, a serious etiologic factor for male subfertility in modern society. Reproduction 2017, 154, R123–R131. [Google Scholar] [CrossRef] [Green Version]
- Chavarro, J.E.; Toth, T.L.; Wright, D.L.; Meeker, J.D.; Hauser, R. Body mass index in relation to semen quality, sperm DNA integrity, and serum reproductive hormone levels among men attending an infertility clinic. Fertil. Steril. 2010, 93, 2222–2231. [Google Scholar] [CrossRef] [Green Version]
- Lainez, N.M.; Coss, D. Obesity, Neuroinflammation, and Reproductive Function. Endocrinology 2019, 160, 2719–2736. [Google Scholar] [CrossRef]
- Ulven, T.; Christiansen, E. Dietary Fatty Acids and Their Potential for Controlling Metabolic Diseases Through Activation of FFA4/GPR120. Annu. Rev. Nutr. 2015, 35, 239–263. [Google Scholar] [CrossRef] [PubMed]
- Rossi, S.P.; Walenta, L.; Rey-Ares, V.; Kohn, F.M.; Schwarzer, J.U.; Welter, H.; Calandra, R.S.; Frungieri, M.B.; Mayerhofer, A. Alpha 1 adrenergic receptor-mediated inflammatory responses in human testicular peritubular cells. Mol. Cell. Endocrinol. 2018, 474, 1–9. [Google Scholar] [CrossRef] [PubMed]
- MInguez-Alarcón, L.; Chavarro, J.E.; Mendiola, J.; Roca, M.; Tanrikut, C.; Vioque, J.; Jorgensen, N.; Torres-Cantero, A.M. Fatty acid intake in relation to reproductive hormones and testicular volume among young healthy men. Asian J. Androl. 2017, 19, 184–190. [Google Scholar] [CrossRef] [PubMed]
- Sermondade, N.; Faure, C.; Fezeu, L.; Shayeb, A.G.; Bonde, J.P.; Jensen, T.K.; Van Wely, M.; Cao, J.; Martini, A.C.; Eskandar, M.; et al. BMI in relation to sperm count: An updated systematic review and collaborative meta-analysis. Hum. Reprod. Update 2013, 19, 221–231. [Google Scholar] [CrossRef] [PubMed]
- Hart, R.J.; Doherty, D.A.; McLachlan, R.I.; Walls, M.L.; Keelan, J.A.; Dickinson, J.E.; Skakkebaek, N.E.; Norman, R.J.; Handelsman, D.J. Testicular function in a birth cohort of young men. Hum. Reprod. 2015, 30, 2713–2724. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Du Plessis, S.S.; Cabler, S.; McAlister, D.A.; Sabanegh, E.; Agarwal, A. The effect of obesity on sperm disorders and male infertility. Nat. Rev. Urol. 2010, 7, 153–161. [Google Scholar] [CrossRef]
- Hammoud, A.O.; Meikle, A.W.; Reis, L.O.; Gibson, M.; Peterson, C.M.; Carrell, D.T. Obesity and male infertility: A practical approach. Semin. Reprod. Med. 2012, 30, 486–495. [Google Scholar] [CrossRef]
- Andersen, J.M.; Herning, H.; Aschim, E.L.; Hjelmesaeth, J.; Mala, T.; Hanevik, H.I.; Bungum, M.; Haugen, T.B.; Witczak, O. Body Mass Index Is Associated with Impaired Semen Characteristics and Reduced Levels of Anti-Mullerian Hormone across a Wide Weight Range. PLoS ONE 2015, 10, e0130210. [Google Scholar] [CrossRef]
- McPherson, N.O.; Lane, M. Male obesity and subfertility, is it really about increased adiposity? Asian J. Androl. 2015, 17, 450–458. [Google Scholar] [CrossRef]
- Leisegang, K.; Henkel, R. The impact of male overweight on semen quality and outcome of assisted reproduction. Asian J. Androl. 2014, 16, 787. [Google Scholar] [CrossRef] [PubMed]
- Cardoso-Moreira, M.; Halbert, J.; Valloton, D.; Velten, B.; Chen, C.; Shao, Y.; Liechti, A.; Ascencao, K.; Rummel, C.; Ovchinnikova, S.; et al. Gene expression across mammalian organ development. Nature 2019, 571, 505–509. [Google Scholar] [CrossRef] [PubMed]
- Lumeng, C.N.; Saltiel, A.R. Inflammatory links between obesity and metabolic disease. J. Clin. Investig. 2011, 121, 2111–2117. [Google Scholar] [CrossRef] [PubMed] [Green Version]



{kind=link}
{kind=link}
| Gene | Gene Name | Reference ID | Nucleotide Sequence | Amplicon Size |
|---|---|---|---|---|
| mk IL6 | Interleukin 6 | NM_001042733.2 | 5′-TGGCTGAAAAAGATGGATGCT-3′ 5′-TTGCTGCTCACTACTCTCAAACCT-3′ | 134 |
| mk IL1 β | Interleukin 1 beta | NM_001042756.1 | 5′-AAAGCTTGGTGATGTCTGGTC-3′ 5′-GGACATGGAGAACACCACTTG-3′ | 89 |
| NLRP3 | NLR family, pyrin domain containing 3 | NM_001114351.1 | 5′-GTGTTTCGAATCCCACTGTG-3′ 5′-TCTGCTTCTCACGTACTTTCTG-3′ | 143 |
| IL6 | Interleukin 6 | NM_000600.4 | 5′-AACCTGAACCTTCCAAAGATGG-3′ 5′-TCTGGCTTGTTCCTCACTACT-3′ | 159 |
| CNN1 | Calponin | NM_001308342.2 | 5′-CGAAGACGAAAGGAAACAAGGT-3′ 5′-GCTTGGGGTCGTAGAGGTG-3′ | 186 |
| GDNF | Glial cell line derived neurotrophic factor | NM_000514.3 | 5’-GCAGACCCATCGCCTTTGAT-3′ 5′-ATCCACACCTTTTAGCGGAATG-3′ | 93 |
| RPL19 | Ribosomal protein L19 | NM_000981 | 5′-AGGCACATGGGCATAGGTAA-3′ 5′-CCATGAGAATCCGCTTGTTT-3′ | 199 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mayerhofer, A.; Dietrich, K.-G.; Urbanski, H.F.; Köhn, F.-M.; Pickl, U.; Trottmann, M.; Kievit, P.; Welter, H. Palmitic Acid Targets Human Testicular Peritubular Cells and Causes a Pro-Inflammatory Response. J. Clin. Med. 2020, 9, 2655. https://doi.org/10.3390/jcm9082655
Mayerhofer A, Dietrich K-G, Urbanski HF, Köhn F-M, Pickl U, Trottmann M, Kievit P, Welter H. Palmitic Acid Targets Human Testicular Peritubular Cells and Causes a Pro-Inflammatory Response. Journal of Clinical Medicine. 2020; 9(8):2655. https://doi.org/10.3390/jcm9082655
Chicago/Turabian StyleMayerhofer, Artur, Kim-Gwendolyn Dietrich, Henryk F. Urbanski, Frank-Michael Köhn, Ulrich Pickl, Matthias Trottmann, Paul Kievit, and Harald Welter. 2020. "Palmitic Acid Targets Human Testicular Peritubular Cells and Causes a Pro-Inflammatory Response" Journal of Clinical Medicine 9, no. 8: 2655. https://doi.org/10.3390/jcm9082655
APA StyleMayerhofer, A., Dietrich, K. -G., Urbanski, H. F., Köhn, F. -M., Pickl, U., Trottmann, M., Kievit, P., & Welter, H. (2020). Palmitic Acid Targets Human Testicular Peritubular Cells and Causes a Pro-Inflammatory Response. Journal of Clinical Medicine, 9(8), 2655. https://doi.org/10.3390/jcm9082655