Carbohydrate and Amino Acid Profiles of Cotton Plant Biomass Products



Abstract
:1. Introduction
2. Materials and Methods
2.1. Cotton Plant Collection and Treatments
2.2. Determination of Amino Acids
2.3. Determination of Carbohydrates
2.4. Data Treatment and Statiscal Analysis
3. Results and Discussion
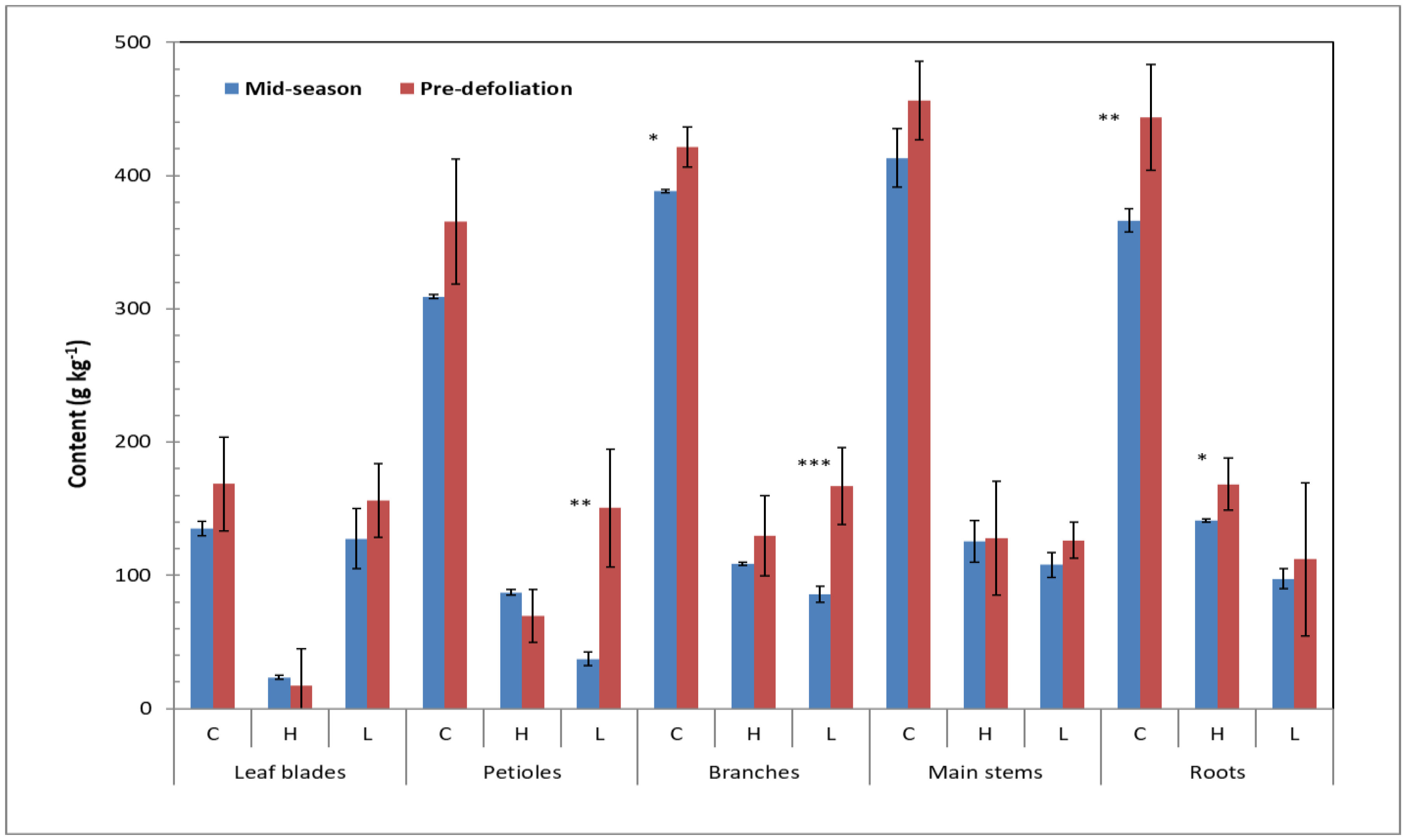
3.1. Carbohydrate Profiles
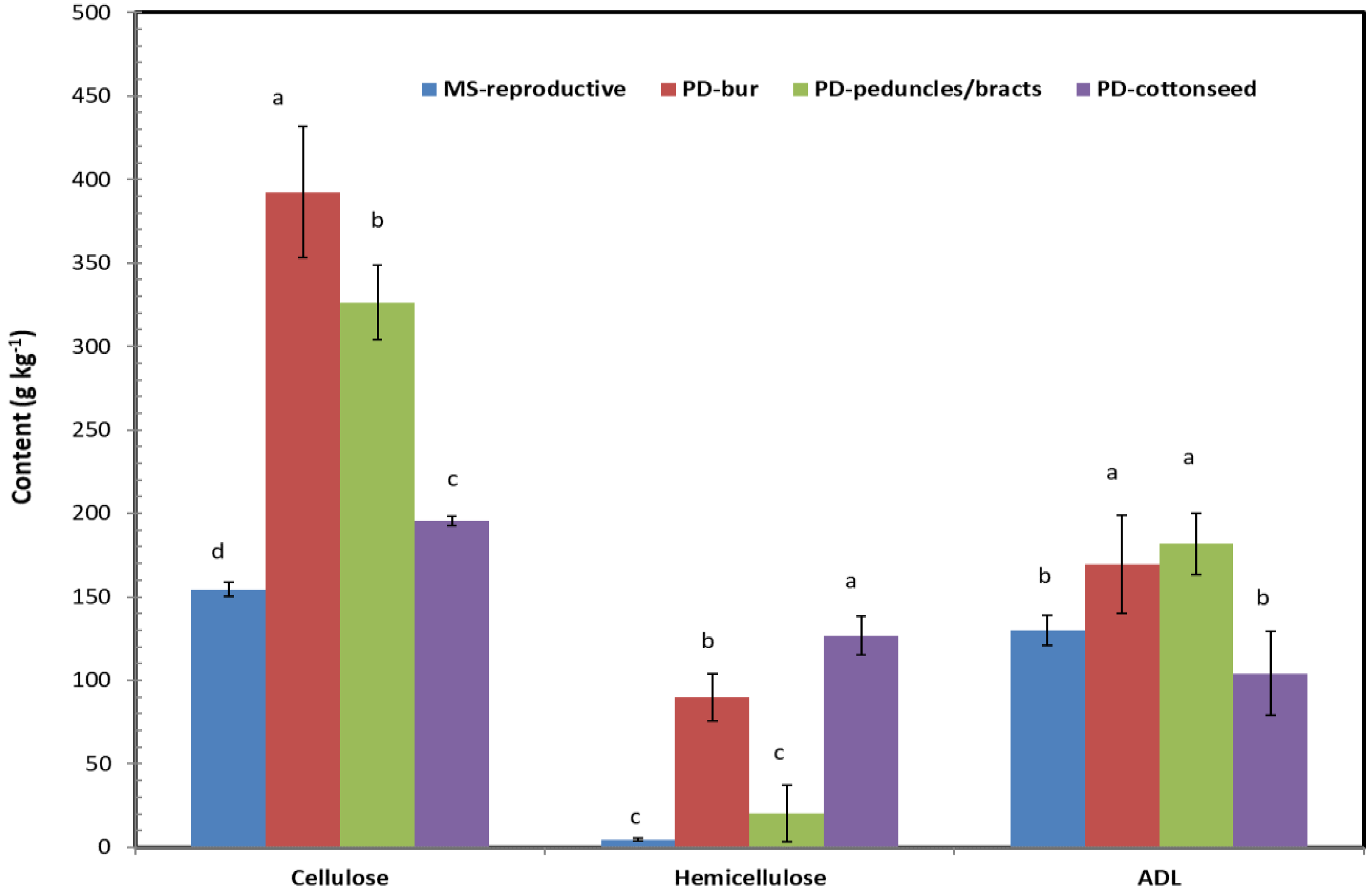
3.2. Structural Carbohydrate Profiles
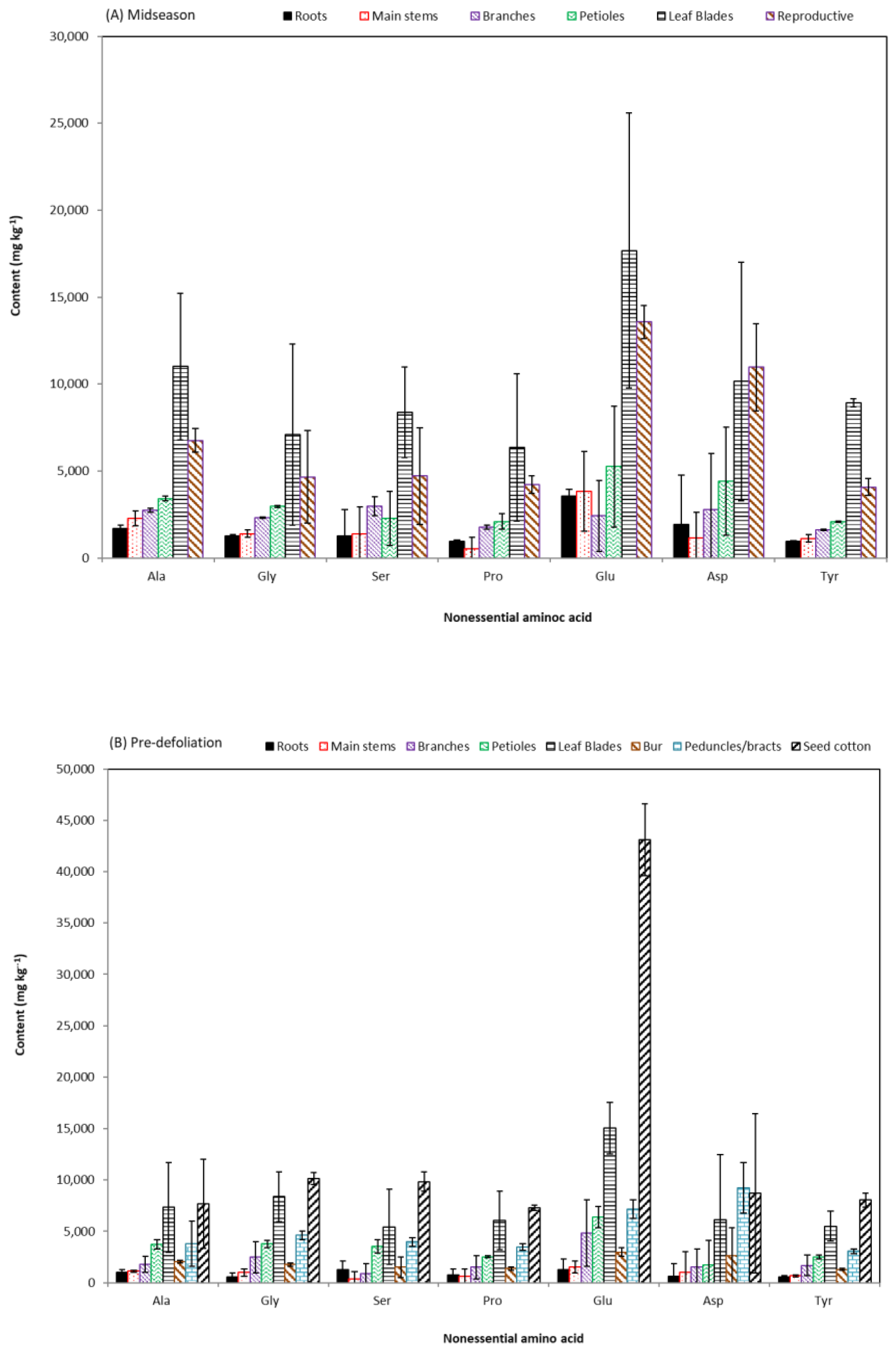
3.3. Amino Acid Profiles
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- He, Z.; Shankle, M.; Zhang, H.; Way, T.R.; Tewolde, H.; Uchimiya, M. Mineral composition of cottonseed is affected by fertilization management practices. Agron. J. 2013, 105, 341–350. [Google Scholar] [CrossRef] [Green Version]
- Tewolde, H.; Shankle, M.W.; Way, T.R.; Adeli, A.; Brooks, J.P.; He, Z. Enhancing management of fall-applied poultry litter with cover crop and subsurface band placement in no-till cotton. Agron. J. 2015, 107, 449–458. [Google Scholar] [CrossRef]
- Mahan, J.; Payton, P. Design and implementation of a rainfed matrix for cotton. Agriculture 2018, 8, 193. [Google Scholar] [CrossRef] [Green Version]
- Mauget, S.; Ulloa, M.; Dever, J. Planting date effects on cotton lint yield and fiber quality in the U.S. Southern High Plains. Agriculture 2019, 9, 82. [Google Scholar] [CrossRef] [Green Version]
- He, Z.; Cheng, H.N.; Chapital, D.C.; Dowd, M.K. Sequential fractionation of cottonseed meal to improve its wood adhesive properties. J. Am. Oil Chem. Soc. 2014, 91, 151–158. [Google Scholar] [CrossRef]
- He, Z.; Uchimiya, S.M.; Guo, M. Production and characterization of biochar from agricultural by-products: Overview and use of cotton biomass residues. In Agricultural and Environmental Applications of Biochar: Advances and Barriers; Guo, M., He, Z., Uchimiya, S.M., Eds.; Soil Science Society of America, Inc.: Madison, WI, USA, 2016; pp. 63–86. [Google Scholar] [CrossRef]
- He, Z.; Guo, M.; Sleighter, R.L.; Zhang, H.; Fortier, C.A.; Hatcher, P.G. Characterization of defatted cottonseed meal-derived pyrolysis bio-oil by ultrahigh resolution electrospray ionization Fourier transform ion cyclotron resonance mass spectrometry. J. Anal. Appl. Pyrol. 2018, 136, 96–106. [Google Scholar] [CrossRef]
- Ren, J.; Li, N.; Li, L.; An, J.-K.; Zhao, L.; Ren, N.-Q. Granulation and ferric oxides loading enable biochar derived from cotton stalk to remove phosphate from water. Bioresour. Technol. 2015, 178, 119–125. [Google Scholar] [CrossRef]
- Kutlu, O.; Kocar, G. Upgrading lignocellulosic waste to fuel by torrefaction: Characterisation and process optimization by response surface methodology. Int. J. Energy Res. 2018, 42, 4746–4760. [Google Scholar] [CrossRef]
- Al Afif, R.; Anayah, S.S.; Pfeifer, C. Batch pyrolysis of cotton stalks for evaluation of biochar energy potential. Renew. Energy 2020, 147, 2250–2258. [Google Scholar] [CrossRef] [Green Version]
- Zubair, M.; Latif, A.; Rao, A.Q.; Azam, S.; Shahid, N.; Samiullah, T.R.; Yasmeen, A.; Shahid, A.A.; Nasir, I.A.; Husnain, T. A combinational approach of enhanced methanol production and double Bt genes for broad spectrum insect resistance in transgenic cotton. Mol. Biotechnol. 2019, 61, 663–673. [Google Scholar] [CrossRef]
- Kirkan, B.; Sarikurkcu, C.; Copuroglu, M.; Cengiz, M.; Tepe, B. Is it possible to use the stalks of Gossypium hirsitum L., an important by-product of cotton cultivation, as an alternative source of bioactive components? Eur. Food Res. Technol. 2018, 244, 1065–1071. [Google Scholar] [CrossRef]
- Fidan, M.S.; Alma, M.H.; Bektas, I. Liquefacation of cotton stalks (Gossypium hirsutum L.) with phenol. Wood Res. 2010, 55, 71–80. [Google Scholar]
- Egbuta, M.; McIntosh, S.; Waters, D.; Vancov, T.; Liu, L. Biological importance of cotton by-products relative to chemical constituents of the cotton plant. Molecules 2017, 22, 93. [Google Scholar] [CrossRef] [Green Version]
- Wanjura, J.D.; Barnes, E.M.; Kelley, M.S.; Holt, G.A.; Pelletier, M.G. Quantification and characterization of cotton crop biomass residue. Ind. Crop. Prod. 2014, 56, 94–104. [Google Scholar] [CrossRef]
- Liu, Y.; He, Z.; Shankle, M.; Tewolde, H. Compositional features of cotton plant biomass fractions characterized by attenuated total reflection Fourier transform infrared spectroscopy. Ind. Crop. Prod. 2016, 79, 283–286. [Google Scholar] [CrossRef]
- He, Z.; Zhang, H.; Tewolde, H.; Shankle, M. Chemical characterization of cotton plant parts for multiple uses. Agric. Environ. Lett. 2017, 2, 110044. [Google Scholar] [CrossRef] [Green Version]
- Guo, M.; Xiao, P.; Li, H. Valorization of agricultural byproducts through conversion to biochar and bio-oil. In Byproducts from Agriculture and Fisheries: Adding Value for Food, Feed, Pharma, and Fuels; Simpson, B.K., Aryee, A.N., Toldrá, F., Eds.; John Wiley & Sons Ltd.: Hoboken, NY, USA, 2020; pp. 501–522. [Google Scholar]
- He, Z.; Senwo, Z.N.; Zou, H.; Tazisong, I.A.; Martens, D.A. Amino compounds in poultry litter, litter-amended pasture soils and grass shoots. Pedosphere 2014, 24, 178–185. [Google Scholar] [CrossRef]
- He, Z.; Olk, D.C.; Waldrip, H.M. Soil amino compound and carbohydrate contents influenced by organic amendments. In Applied Manure and Nutrient Chemistry for Sustainable Agriculture and Environment; He, Z., Zhang, H., Eds.; Springer: Amsterdam, The Netherland, 2014; pp. 69–82. [Google Scholar]
- Martens, D.A.; Loeffelmann, K.L. Improved accounting of carbohydrate carbon from plants and soils. Soil Biol. Biochem. 2002, 34, 1393–1399. [Google Scholar] [CrossRef] [Green Version]
- Guidi, C.; Cannella, D.; Leifeld, J.; Rodeghiero, M.; Magid, J.; Gianelle, D.; Vesterdal, L. Carbohydrates and thermal properties indicate a decrease in stable aggregate carbon following forest colonization of mountain grassland. Soil Biol. Biochem. 2015, 86, 135–145. [Google Scholar] [CrossRef]
- Ghasemian, M.; Zilouei, H.; Asadinezhad, A. Enhanced biogas and biohydrogen production from cotton plant wastes using alkaline pretreatment. Energy Fuels 2016, 30, 10484–10493. [Google Scholar] [CrossRef]
- He, Z.; Zhang, H.; Olk, D.C.; Shankle, M.; Way, T.R.; Tewolde, H. Protein and fiber profiles of cottonseed from upland cotton with different fertilizations. Mod. Appl. Sci. 2014, 8, 97–105. [Google Scholar] [CrossRef] [Green Version]
- He, Z.; Zhang, H.; Olk, D.C. Chemical composition of defatted cottonseed and soy meal products. PLoS ONE 2015, 10, e0129933. [Google Scholar] [CrossRef] [PubMed]
- Amthor, J.S.; Bar-Even, A.; Hanson, A.D.; Millar, A.H.; Stitt, M.; Sweetlove, L.J.; Tyerman, S.D. Engineering strategies to boost crop productivity by cutting respiratory carbon loss. Plant Cell 2019, 31, 297–314. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, S.; Wang, X.; Yin, X.; Savoy, H.J.; McClure, A.; Essington, M.E. Ammonia volatilizationloss and corn nitrogen nutrition and productivity with efficiency enhanced UAN and urea under no-tillage. Sci. Rep. 2019, 9, 6610. [Google Scholar] [CrossRef]
- Tewolde, H.; Shankle, M.W.; Way, T.R.; Pote, D.H.; Sistani, K.R.; He, Z. Poultry litter band placement affects accessibility and conservation of nutrients and cotton yield. Agron. J. 2018, 110, 675–684. [Google Scholar] [CrossRef]
- Olk, D.C.; Fortuna, A.; Honeycutt, C.W. Using anion chromatography-pulsed amperometry to measure amino compounds in dairy manure-amended soils. Soil Sci. Soc. Am. J. 2008, 72, 1711–1720. [Google Scholar] [CrossRef]
- Olk, D.C. Improved analytical techniques for carbohydrates, amino compounds, and phenols: Tools for understanding soil processes. Soil Sci. Soc. Am. J. 2008, 72, 1672–1682. [Google Scholar] [CrossRef]
- He, Z.; Klasson, K.T.; Wang, D.; Li, N.; Zhang, H.; Zhang, D.; Wedegaertner, T.C. Pilot-scale production of washed cottonseed meal and co-products. Mod. Appl. Sci. 2016, 10, 25–33. [Google Scholar] [CrossRef] [Green Version]
- Martins, M.d.R.; Angers, D.A.; Corá, J.E. Carbohydrate composition and water-stable aggregation of an Oxisol as affected by crop sequence under no-till. Soil Sci. Soc. Am. J. 2012, 76, 475–484. [Google Scholar] [CrossRef]
- Zhang, Y.; Idowu, O.J.; Brewer, C.E. Using agricultural residue biochar to improve soil quality of desert soils. Agriculture 2016, 6, 10. [Google Scholar] [CrossRef] [Green Version]
- Vancov, T.; Palmer, J.; Keen, B. A two stage pretreatment process to maximise recovery of sugars from cotton gin trash. Bioresour. Technol. Rep. 2018, 4, 114–122. [Google Scholar] [CrossRef]
- McIntosh, S.; Palmer, J.; Egbuta, M.; Liu, L.; Vancov, T. Refining spent cotton gin trash following essential oil extraction for value added cellulosic sugars. Bioresour. Technol. Rep. 2019, 7, 100223. [Google Scholar] [CrossRef]
- Guan, Z.-H.; Li, X.G.; Wang, L.; Mou, X.M.; Kuzyakov, Y. Conversion of Tibetan grasslands to croplands decreases accumulation of microbially synthesized compounds in soil. Soil Biol. Biochem. 2018, 123, 10–20. [Google Scholar] [CrossRef]
- Sugiura, Y.; Watanabe, M.; Nonoyama, Y.; Sakagami, N.; Guo, Y.; Murayama, S. Saccharides of ectomycorrhizal fungal sclerotia as sources of forest soil polysaccharides. Soil Sci. Plant Nutr. 2017, 63, 426–433. [Google Scholar] [CrossRef] [Green Version]
- Keshav, P.K.; Naseeruddin, S.; Rao, L.V. Improved enzymatic saccharification of steam exploded cotton stalk using alkaline extraction and fermentation of cellulosic sugars into ethanol. Bioresour. Technol. 2016, 214, 363–370. [Google Scholar] [CrossRef]
- Amthor, J.S. Efficiency of lignin biosynthesis: A quantitative analysis. Ann. Bot. 2003, 91, 673–695. [Google Scholar] [CrossRef] [Green Version]
- Pauly, M.; Albersheim, P.; Darvill, A.; York, W.S. Molecular domains of the cellulose/xyloglucan network in the cell walls of higher plants. Plant J. 1999, 20, 629–639. [Google Scholar] [CrossRef]
- Wakabayashi, K.; Soga, K.; Hoson, T. Modification of cell wall architecture in gramineous plants under altered gravity conditions. Biol. Sci. Space 2009, 23, 137–142. [Google Scholar] [CrossRef]
- Binod, P.; Kuttiraja, M.; Archana, M.; Janu, K.U.; Sindhu, R.; Sukumaran, R.K.; Pandey, A. High temperature pretreatment and hydrolysis of cotton stalk for producing sugars for bioethanol production. Fuel 2012, 92, 340–345. [Google Scholar] [CrossRef]
- Mortimer, J.C.; Miles, G.P.; Brown, D.M.; Zhang, Z.; Segura, M.P.; Weimar, T.; Yu, X.; Seffen, K.A.; Stephens, E.; Turner, S.R. Absence of branches from xylan in Arabidopsis gux mutants reveals potential for simplification of lignocellulosic biomass. Proc. Natl. Acad. Sci. USA 2010, 107, 17409–17414. [Google Scholar] [CrossRef] [Green Version]
- Bertrand, J.A.; Sudduth, T.Q.; Condon, A.; Jenkins, T.C.; Calhoun, M.C. Nutrient content of whole cottonseed. J. Dairy Sci. 2005, 88, 1470–1477. [Google Scholar] [CrossRef] [Green Version]
- Dahl-Lassen, R.; van Hecke, J.; Jørgensen, H.; Bukh, C.; Andersen, B.; Schjoerring, J.K. High-throughput analysis of amino acids in plant materials by single quadrupole mass spectrometry. Plant Methods 2018, 14, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wyngaard, N.; Cabrera, M.L.; Shober, A.; Kanwar, R. Fertilization strategy can affect the estimation of soil nitrogen mineralization potential with chemical methods. Plant Soil 2018, 432, 75–89. [Google Scholar] [CrossRef]
- Grewal, J.; Khare, S. 2-Pyrrolidone synthesis from g-aminobutyric acid produced by Lactobacillus brevis under solid-state fermentation utilizing toxic deoiled cottonseed cake. Bioprocess Biosyst. Eng. 2017, 40, 145–152. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Fucose | Rhamnose | Arabinose | Galactose | Glucose | Xylose | Mannose | GM/AX | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| MS | PD | MS | PD | MS | PD | MS | PD | MS | PD | MS | PD | MS | PD | MS | PD | ||
| Leaf blades | A | 0.9 | 1.1 | 10.9 | 9.9 | 17.9 | 14.0 | 20.9 | 23.9 | 33.8 | 30.0 | 12.1 | 14.3 | 5.6 | 8.6 | 0.89 | 1.15 |
| SD | 0.1 | 0.1 | 2.0 | 1.3 | 2.2 | 1.4 | 2.1 | 2.0 | 3.6 | 2.0 | 2.4 | 1.7 | 1.0 | 0.2 | 0.06 | 0.05 | |
| Petioles | A | 1.5 | 1.8 | 11.8 | 13.5 | 23.9 | 23.7 | 17.4 | 23.7 | 43.0 | 22.3 | 40.8 | 56.5 | 7.7 | 8.1 | 0.40 | 0.40 |
| SD | 0.2 | 0.1 | 1.2 | 1.7 | 4.1 | 4.9 | 2.7 | 3.2 | 8.0 | 3.8 | 10.9 | 5.8 | 1.3 | 0.9 | 0.05 | 0.01 | |
| Branch stems | A | 0.7 | 1.0 | 7.4 | 8.6 | 15.5 | 13.2 | 12.3 | 13.2 | 66.1 | 23.9 | 61.9 | 82.2 | 5.8 | 6.1 | 0.23 | 0.20 |
| SD | 0.1 | 0.2 | 0.6 | 0.6 | 1.3 | 1.8 | 0.6 | 1.1 | 7.3 | 8.5 | 3.2 | 3.4 | 0.6 | 0.2 | 0.00 | 0.01 | |
| Main stems | A | 0.3 | 0.8 | 4.8 | 7.1 | 7.1 | 9.7 | 7.0 | 9.3 | 56.3 | 40.4 | 40.0 | 71.7 | 3.2 | 5.3 | 0.22 | 0.19 |
| SD | 0.1 | 0.2 | 1.0 | 1.4 | 1.0 | 1.1 | 0.9 | 0.9 | 8.9 | 14.2 | 7.6 | 19.2 | 1.3 | 0.5 | 0.03 | 0.06 | |
| Roots | A | 0.2 | 0.4 | 5.2 | 5.0 | 6.4 | 7.1 | 6.1 | 7.1 | 65.3 | 37.9 | 27.5 | 65.1 | 2.8 | 3.2 | 0.26 | 0.14 |
| SD | 0.1 | 0.1 | 0.4 | 0.8 | 0.7 | 0.8 | 0.5 | 0.5 | 8.8 | 9.6 | 2.3 | 7.4 | 0.2 | 0.3 | 0.03 | 0.01 | |
| Fucose | Rhamnose | Arabinose | Galactose | Glucose | Xylose | Mannose | Cellulose | Hemicellulose | |
|---|---|---|---|---|---|---|---|---|---|
| Rhamnose | 0.904 *** | ||||||||
| Arabinose | 0.673 ** | 0.691 ** | |||||||
| Galactose | 0.802 *** | 0.854 *** | 0.744 ** | ||||||
| Glucose | −0.412 | −0.350 | −0.232 | −0.474 | |||||
| Xylose | 0.011 | −0.329 | −0.114 | −0.342 | −0.207 | ||||
| Mannose | 0.834 *** | 0.861 *** | 0.612 * | 0.778 ** | −0.116 | -0.302 | |||
| Cellulose | −0.038 | −0.313 | −0.456 | −0.554 * | 0.238 | 0.719 ** | −0.245 | ||
| Hemi cellulose | −0.468 | −0.739 ** | −0.422 | −0.749 ** | 0.312 | 0.726 ** | −0.640 * | 0.731 ** | |
| ADL | 0.243 | 0.27 | −0.242 | 0.306 | −0.640 * | 0.041 | 0.161 | −0.007 | −0.368 |
| Mid-Season | Leaf Blades | Petioles | Branches | Main Stems | Roots | Reproductive | ||
|---|---|---|---|---|---|---|---|---|
| A | 8.6 | 56.2 | 120.9 | 140.1 | 142.5 | 14.2 | ||
| SD | 5.6 | 22.8 | 13.6 | 40.6 | 28.7 | 6.3 | ||
| Pre-Defoliation | Leaf Blades | Petioles | Branches | Main Stems | Roots | Bur | Peduncles/Bracts | Cottonseed |
| A | 17.8 | 0.2 | 54.9 | 30.2 | 0.5 | 0.6 | 0.1 | 4.9 |
| SD | 5.2 | 0.2 | 46.8 | 39.7 | 0.3 | 0.5 | 0.0 | 5.2 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
He, Z.; Olk, D.C.; Tewolde, H.; Zhang, H.; Shankle, M. Carbohydrate and Amino Acid Profiles of Cotton Plant Biomass Products. Agriculture 2020, 10, 2. https://doi.org/10.3390/agriculture10010002
He Z, Olk DC, Tewolde H, Zhang H, Shankle M. Carbohydrate and Amino Acid Profiles of Cotton Plant Biomass Products. Agriculture. 2020; 10(1):2. https://doi.org/10.3390/agriculture10010002
Chicago/Turabian StyleHe, Zhongqi, Dan C. Olk, Haile Tewolde, Hailin Zhang, and Mark Shankle. 2020. "Carbohydrate and Amino Acid Profiles of Cotton Plant Biomass Products" Agriculture 10, no. 1: 2. https://doi.org/10.3390/agriculture10010002
APA StyleHe, Z., Olk, D. C., Tewolde, H., Zhang, H., & Shankle, M. (2020). Carbohydrate and Amino Acid Profiles of Cotton Plant Biomass Products. Agriculture, 10(1), 2. https://doi.org/10.3390/agriculture10010002