Vertical Root Distribution of Different Cover Crops Determined with the Profile Wall Method


 and
and 
Abstract
:1. Introduction
2. Materials and Methods
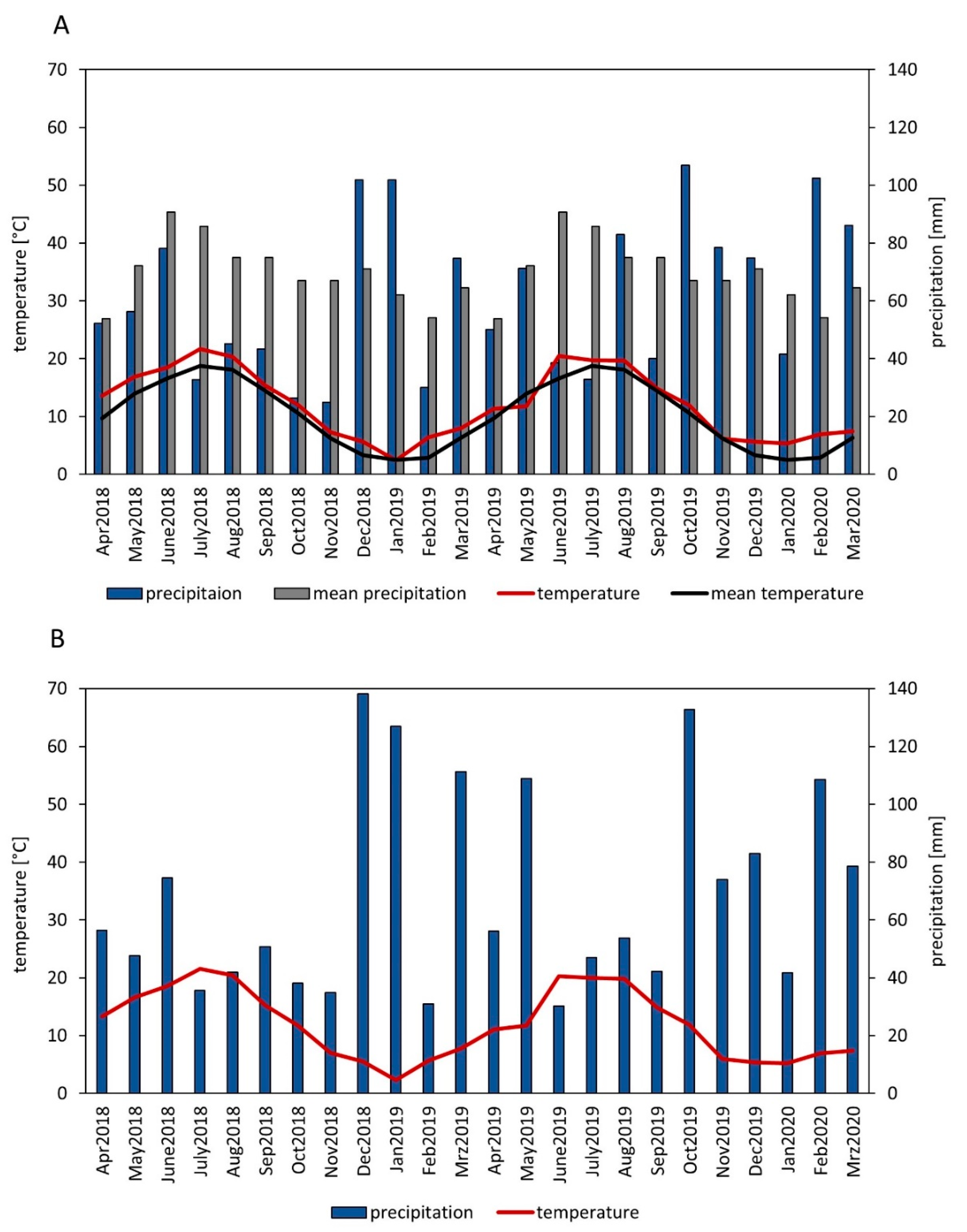
2.1. Experimental Site
2.2. Experimental Design and Management Practices
2.3. Profile Wall Method
2.4. Statistical Analysis
3. Results
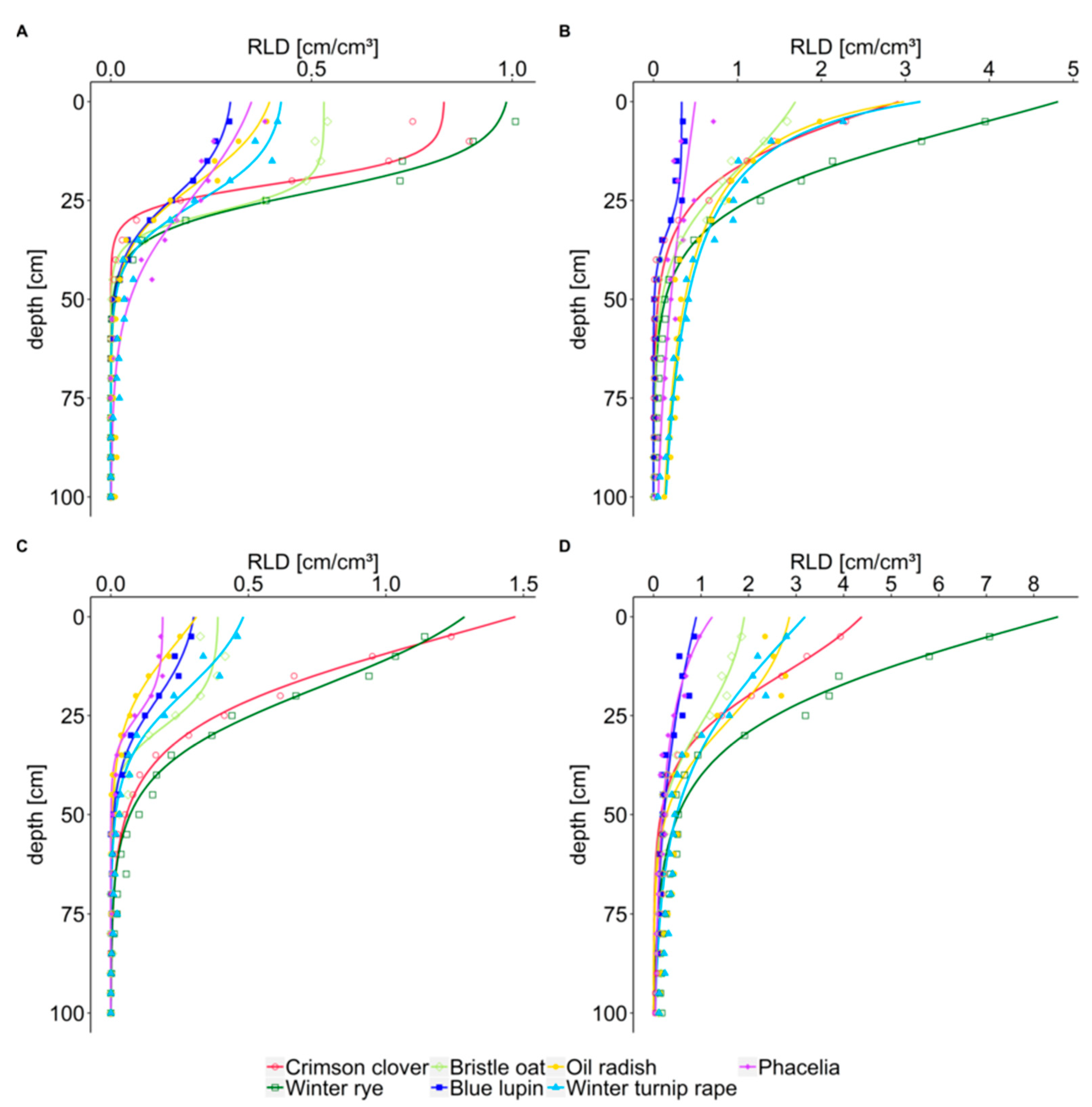
3.1. Root Length Density and Rooting Depth
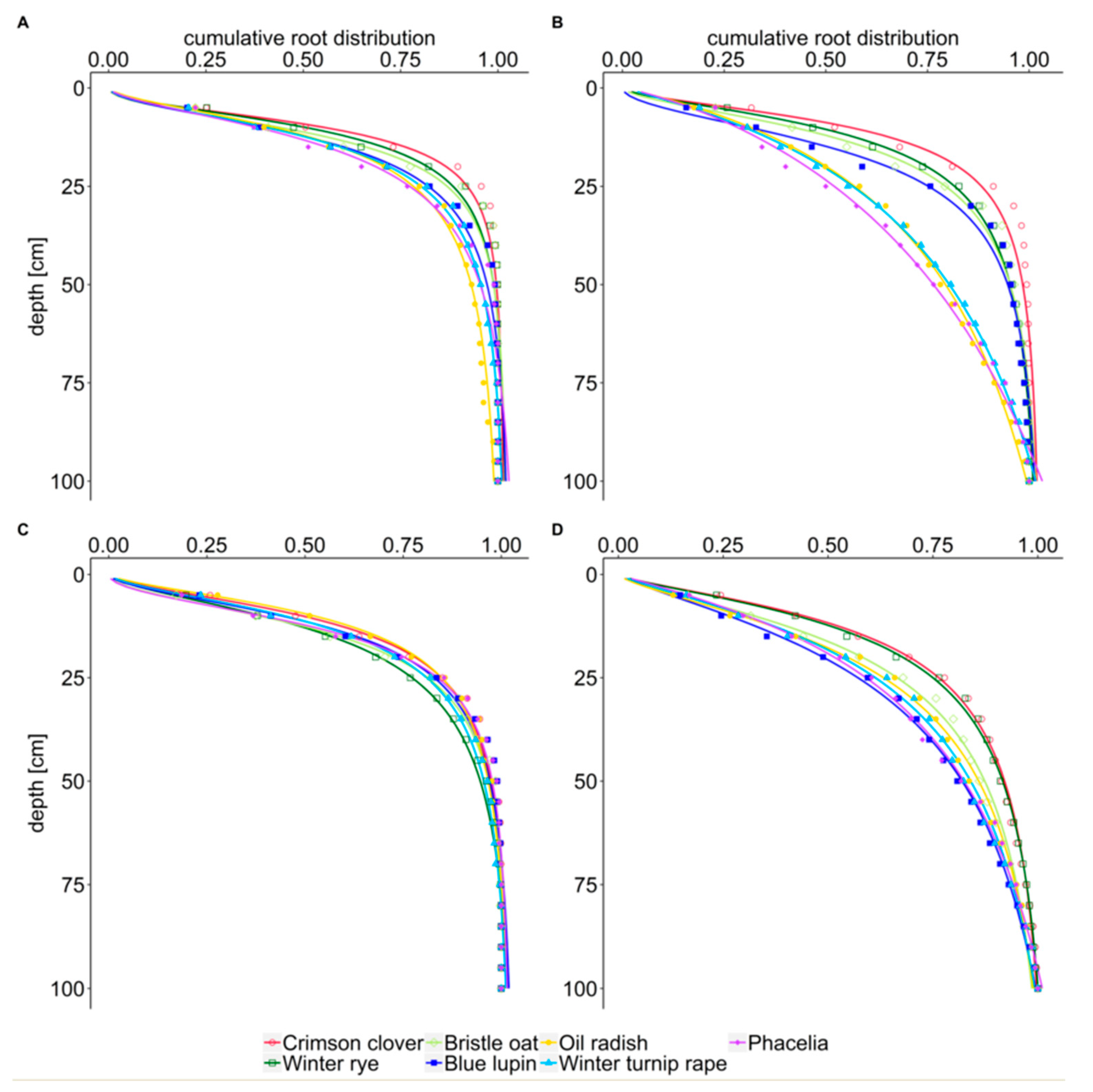
3.2. Cumulative Distribution of Root Length
3.3. Share of Root Length in Large-Sized Biopores in the Subsoil
4. Discussion
4.1. Characterization of Cover Crop Rooting: Root Length Density and Rooting Depth
4.2. Cumulative Distribution of Root Length
4.3. Winter Hardiness
4.4. Share of Root Length in Large-Sized Biopores in the Subsoil
4.5. Cover Crop Rooting Types
- (1)
- Topsoil-allocated with large diameters: e.g., legumes like vetch, faba bean, Egyptian clover
- (2)
- Topsoil-allocated with very dense root system and many fine roots: e.g., rye, buckwheat
- (3)
- Root density group with highly branched dense rooting systems: e.g., oil radish, camelina, phacelia
- (4)
- Low-order axes types like linseed and bristle oat with low-branched primary roots.
4.6. The Profile Wall Method
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Wezel, A.; Casagrande, M.; Celette, F.; Vian, J.-F.; Ferrer, A.; Peigné, J. Agroecological practices for sustainable agriculture. A review. Agron. Sustain. Dev. 2014, 34, 1–20. [Google Scholar] [CrossRef] [Green Version]
- Wittwer, R.A.; Dorn, B.; Jossi, W.; van der Heijden, M.G.A. Cover crops support ecological intensification of arable cropping systems. Sci. Rep. 2017, 7, 41911. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaye, J.P.; Quemada, M. Using cover crops to mitigate and adapt to climate change. A review. Agron. Sustain. Dev. 2017, 37. [Google Scholar] [CrossRef]
- Hartwig, N.L.; Ammon, H.U. Cover crops and living mulches. Weed Sci. 2002, 50, 688–699. [Google Scholar] [CrossRef]
- Thorup-Kristensen, K.; Magid, J.; Jensen, L.S. Catch crops and green manures as biological tools in nitrogen management in temperate zones. Adv. Agron. 2003, 79, 227–302. [Google Scholar] [CrossRef]
- de Baets, S.; Poesen, J.; Gyssels, G.; Knapen, A. Effects of grass roots on the erodibility of topsoils during concentrated flow. Geomorphology 2006, 76, 54–67. [Google Scholar] [CrossRef]
- Poeplau, C.; Don, A. Carbon sequestration in agricultural soils via cultivation of cover crops—A meta-analysis. Agric. Ecosyst. Environ. 2015, 200, 33–41. [Google Scholar] [CrossRef]
- Jian, J.; Du, X.; Reiter, M.S.; Stewart, R.D. A meta-analysis of global cropland soil carbon changes due to cover cropping. Soil Biol. Biochem. 2020, 143, 107735. [Google Scholar] [CrossRef]
- Liu, A.; Ma, B.L.; Bomke, A.A. Effects of Cover Crops on Soil Aggregate Stability, total Organic Carbon, and Polysaccharides. Soil Sci. Soc. Am. J. 2005, 69, 2041–2048. [Google Scholar] [CrossRef]
- Bodner, G.; Leitner, D.; Nakhforoosh, A.; Sobotik, M.; Moder, K.; Kaul, H.-P. A statistical approach to root system classification. Front. Plant. Sci. 2013, 4, 292. [Google Scholar] [CrossRef] [Green Version]
- Kätterer, T.; Bolinder, M.A.; Andrén, O.; Kirchmann, H.; Menichetti, L. Roots contribute more to refractory soil organic matter than above-ground crop residues, as revealed by a long-term field experiment. Agric. Ecosyst. Environ. 2011, 141, 184–192. [Google Scholar] [CrossRef] [Green Version]
- Yu, Y.; Loiskandl, W.; Kaul, H.-P.; Himmelbauer, M.; Wei, W.; Chen, L.; Bodner, G. Estimation of runoff mitigation by morphologically different cover crop root systems. J. Hydrol. 2016, 538, 667–676. [Google Scholar] [CrossRef] [Green Version]
- Bodner, G.; Leitner, D.; Kaul, H.-P. Coarse and fine root plants affect pore size distributions differently. Plant Soil 2014, 380, 133–151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sainju, U.M.; Singh, B.P.; Whitehead, W.F. Cover Crop Root Distribution and Its Effects on Soil Nitrogen Cycling. Agron. J. 1998, 90, 511–518. [Google Scholar] [CrossRef]
- Herrera, J.M.; Feil, B.; Stamp, P.; Liedgens, M. Root growth and nitrate-nitrogen leaching of catch crops following spring wheat. J. Environ. Qual. 2010, 39, 845–854. [Google Scholar] [CrossRef]
- Kristensen, H.L.; Thorup-Kristensen, K. Root Growth and Nitrate Uptake of Three Different Catch Crops in Deep Soil Layers. Soil Sci. Soc. Am. J. 2004, 68, 529. [Google Scholar] [CrossRef]
- Sapkota, T.B.; Askegaard, M.; Lægdsmand, M.; Olesen, J.E. Effects of catch crop type and root depth on nitrogen leaching and yield of spring barley. Field Crop. Res. 2012, 125, 129–138. [Google Scholar] [CrossRef]
- Thorup-Kristensen, K. Are differences in root growth of nitrogen catch crops important for their ability to reduce soil nitrate-N content, and how can this be measured? Plant Soil 2001, 230, 185–195. [Google Scholar] [CrossRef]
- Thorup-Kristensen, K.; Rasmussen, C.R. Identifying new deep-rooted plant species suitable as undersown nitrogen catch crops. J. Soil Water Conserv. 2015, 70, 399–409. [Google Scholar] [CrossRef]
- Wendling, M.; Büchi, L.; Amossé, C.; Sinaj, S.; Walter, A.; Charles, R. Influence of root and leaf traits on the uptake of nutrients in cover crops. Plant Soil 2016, 409, 419–434. [Google Scholar] [CrossRef] [Green Version]
- Bodner, G.; Himmelbauer, M.; Loiskandl, W.; Kaul, H.-P. Improved evaluation of cover crop species by growth and root factors. Agron. Sustain. Dev. 2010, 30, 455–464. [Google Scholar] [CrossRef]
- Vos, J.; van der Putten, P.E.L.; Hassan Hussein, M.; van Dam, A.M.; Leffelaar, P.A. Field observations on nitrogen catch crops—II. Root length and root length distribution in relation to species and nitrogen supply. Plant Soil 1998, 201, 149–155. [Google Scholar] [CrossRef]
- Fan, J.; McConkey, B.; Wang, H.; Janzen, H. Root distribution by depth for temperate agricultural crops. Field Crop. Res. 2016, 189, 68–74. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Ghodrati, M. Preferential Transport of Nitrate through Soil Columns Containing Root Channels. Soil Sci. Soc. Am. J. 1994, 58, 653–659. [Google Scholar] [CrossRef]
- Kautz, T. Research on subsoil biopores and their functions in organically managed soils: A review. Renew. Agric. Food Syst. 2015, 30, 318–327. [Google Scholar] [CrossRef]
- Perkons, U.; Kautz, T.; Uteau, D.; Peth, S.; Geier, V.; Thomas, K.; Lütke Holz, K.; Athmann, M.; Pude, R.; Köpke, U. Root-length densities of various annual crops following crops with contrasting root systems. Soil Tillage Res. 2014, 137, 50–57. [Google Scholar] [CrossRef]
- Han, E.; Kautz, T.; Perkons, U.; Uteau, D.; Peth, S.; Huang, N.; Horn, R.; Köpke, U. Root growth dynamics inside and outside of soil biopores as affected by crop sequence determined with the profile wall method. Biol. Fertil. Soils 2015, 51, 847–856. [Google Scholar] [CrossRef]
- Kautz, T.; Perkons, U.; Athmann, M.; Pude, R.; Köpke, U. Barley roots are not constrained to large-sized biopores in the subsoil of a deep Haplic Luvisol. Biol. Fertil. Soils 2013, 49, 959–963. [Google Scholar] [CrossRef]
- Athmann, M.; Kautz, T.; Pude, R.; Köpke, U. Root growth in biopores—Evaluation with in situ endoscopy. Plant Soil 2013, 371, 179–190. [Google Scholar] [CrossRef]
- Böhm, W. Methods of Studying Root Systems; Springer: Berlin/Heidelberg, Germany, 1979; ISBN 978-3-642-67284-2. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2018; Available online: https://www.R-project.org/ (accessed on 13 February 2020).
- Schenk, H.J.; Jackson, R.B. The Global Biogeography of roots. Ecol. Monogr. 2002, 72, 311–328. [Google Scholar] [CrossRef]
- Bodner, G.; Loiskandl, W.; Hartl, W.; Erhart, E.; Sobotik, M. Characterization of Cover Crop Rooting Types from Integration of Rhizobox Imaging and Root Atlas Information. Plants 2019, 8, 514. [Google Scholar] [CrossRef] [Green Version]
- Hamblin, A.P.; Tennant, D. Root length density and water uptake in cereals and grain legumes: How well are they correlated. Aust. J. Agric. Res. 1987, 38, 513. [Google Scholar] [CrossRef]
- Munkholm, L.J.; Hansen, E.M. Catch crop biomass production, nitrogen uptake and root development under different tillage systems. Soil Use Manag. 2012, 28, 517–529. [Google Scholar] [CrossRef]
- Athmann, M.; Sondermann, J.; Kautz, T.; Köpke, U. Comparing Macropore Exploration by Faba Bean, Wheat, Barley and Oilseed Rape Roots Using In Situ Endoscopy. J. Soil Sci. Plant. Nutr. 2019, 19, 689–700. [Google Scholar] [CrossRef]
- Kutschera, L.; Lichtenegger, E. Wurzelatlas Mitteleuropäischer Ackerunkräuter und Kulturpflanzen; DLG-Verlag: Frankfurt am Main, Germany, 1960. [Google Scholar]
- Kutschera, L.; Lichtenegger, E.; Sobotik, M. Wurzelatlas der Kulturpflanzen Gemäßigter Gebiete mit Arten des Feldgemüsebaues; DLG-Verlag: Frankfurt am Main, Germany, 2009. [Google Scholar]
- Smith, A.G.; Han, E.; Petersen, J.; Faircloth Olsen, N.A.; Giese, C.; Athmann, M.; Dresbøll, D.B.; Thorup-Kristensen, K. RootPainter: Deep Learning Segmentation of Biological Images with Corrective Annotation. bioRxiv 2020. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| 2018/2019 | 2019/2020 | |
|---|---|---|
| pH (CaCl2) | 6.2 | 6.3 |
| P2O5 [mg/100 g] | 5 | 8 |
| K2O [mg/100 g] | 21 | 15 |
| MgO [mg/100 g] | 12 | 10 |
| total organic carbon [%] | 1.1 | 0.75 |
| total nitrogen [%] | 0.11 | 0.10 |
| 2018/2019 | 2019/2020 | |
|---|---|---|
| 0–30 cm | 17.1 | 53.9 |
| 30–60 cm | 3.2 | 26.1 |
| 60–90 cm | 3.4 | 11.7 |
| Cover Crop Species | Cultivar | Family | Winter Hardiness | Recommended Seed Rate [kg/ha] |
|---|---|---|---|---|
| crimson clover (Trifolium incarnatum L.) | Linkarus | Leguminosae | winter hardy | 30 |
| winter rye (Secale cereale L.) | Bonfire | Poaceae | winter hardy | 120 |
| bristle oat (Avena strigosa Schreb.) | Pratex | Poaceae | non- winter hardy | 80 |
| blue lupin (Lupinus angustifolius L.) | Boruta | Leguminosae | non- winter hardy | 120 |
| oil radish (Raphanus sativus L. var. oleiformis Pers.) | Siletina | Brassicaceae | non- winter hardy | 25 |
| winter turnip rape (Brassica rapa L. var. silvestris (Lam.) Briggs) | Jupiter | Brassicaceae | winter hardy | 15 |
| phacelia (Phacelia tanacetifolia Benth.) | Beehappy | Boraginaceae | non- winter hardy | 12 |
| Year | Date | Cover Crop | Rooting Depth [cm] | RLD5 [cm cm−3] | Soil Depth [cm] at Half Maximal RLD |
|---|---|---|---|---|---|
| 2018/2019 | October | crimson clover | 42 ± 13 b | 0.82 | 21 |
| winter rye | 57 ± 13 ab | 0.97 | 23 | ||
| bristle oat | 65 ± 5 ab | 0.53 | 28 | ||
| blue lupin | 62 ± 8 ab | 0.29 | 25 | ||
| oil radish | 77 ± 20 a | 0.37 | 22 | ||
| winter turnip rape | 75 ± 9 a | 0.41 | 26 | ||
| phacelia | 53 ± 16 ab | 0.33 | 28 | ||
| March | crimson clover | 68 ± 8 | 1.22 | 15 | |
| winter rye | 82 ± 8 | 1.17 | 21 | ||
| bristle oat | 73 ± 20 | 0.39 | 28 | ||
| blue lupin | 62 ± 15 | 0.28 | 23 | ||
| oil radish | 70 ± 0 | 0.26 | 14 | ||
| winter turnip rape | 78 ± 6 | 0.44 | 21 | ||
| phacelia | 55 ± 13 | 0.19 | 25 | ||
| 2019/2020 | October | crimson clover | 82 ± 12 | 2.22 | 11 |
| winter rye | 100 ± 0 | 3.97 | 14 | ||
| bristle oat | 97 ± 3 | 1.53 | 21 | ||
| blue lupin | 80 ± 30 | 0.33 | 33 | ||
| oil radish | 100 ± 0 | 2.00 | 10 | ||
| winter turnip rape | 100 ± 0 | 2.14 | 10 | ||
| phacelia | 100 ± 0 | 0.47 | 41 | ||
| March | crimson clover | 100 ± 0 | 3.90 | 18 | |
| winter rye | 100 ± 0 | 7.05 | 16 | ||
| bristle oat | 100 ± 0 | 1.83 | 28 | ||
| blue lupin | 100 ± 0 | 0.81 | 15 | ||
| oil radish | 100 ± 0 | 2.75 | 28 | ||
| winter turnip rape | 100 ± 0 | 2.81 | 22 | ||
| phacelia | 100 ± 0 | 0.98 | 16 |
| Year | Date | Cover Crop | c | x50 (cm) | x95 (cm) |
|---|---|---|---|---|---|
| 2018/2019 | October | crimson clover | −2.280 | 9.3 | 29.6 |
| winter rye | −2.029 | 10.2 | 34.9 | ||
| bristle oat | −2.184 | 11.5 | 36.6 | ||
| blue lupin | −1.955 | 12.7 | 42.6 | ||
| oil radish | −1.683 | 12.3 | 58.3 | ||
| winter turnip rape | −1.847 | 12.7 | 47.7 | ||
| phacelia | −1.710 | 14.1 | 48.6 | ||
| March | crimson clover | −1.738 | 10.6 | 41.4 | |
| winter rye | −1.681 | 13.7 | 51.1 | ||
| bristle oat | −2.007 | 13.1 | 44.1 | ||
| blue lupin | −1.833 | 11.8 | 42.9 | ||
| oil radish | −1.624 | 9.9 | 43.3 | ||
| winter turnip rape | −1.687 | 11.8 | 48.0 | ||
| phacelia | −2.157 | 12.54 | 40.3 | ||
| 2019/2020 | October | crimson clover | −1.749 | 9.1 | 36.4 |
| winter rye | −1.586 | 11.0 | 48.7 | ||
| bristle oat | −1.691 | 12.7 | 48.9 | ||
| blue lupin | −1.995 | 15.4 | 50.9 | ||
| oil radish | −0.962 | 34.6 | 84.7 | ||
| winter turnip rape | −1.005 | 35.8 | 79.1 | ||
| phacelia | −0.853 | 73.3 | 80.8 | ||
| March | crimson clover | −1.423 | 12.5 | 61.3 | |
| winter rye | −1.403 | 13.3 | 62.1 | ||
| bristle oat | −1.460 | 17.6 | 75.3 | ||
| blue lupin | −1.302 | 25.8 | 80.7 | ||
| oil radish | −1.450 | 19.8 | 76.6 | ||
| winter turnip rape | −1.267 | 22.2 | 79.0 | ||
| phacelia | −1.104 | 28.0 | 77.5 |
| Year | Date | Cover Crop | Share of Root Length in Biopores (%) | RLD in Biopores (cm cm−3) | RLD in Bulk Soil (cm cm−3) |
|---|---|---|---|---|---|
| 2018/2019 | October | crimson clover | 0.0 ± 0.0 | 0.00000 ± 0.00000 | 0.00467 ± 0.00467 |
| winter rye | 5.2 ± 6.6 | 0.00122 ± 0.00184 | 0.01203 ± 0.01064 | ||
| bristle oat | 0.0 ± 0.0 | 0.00000 ± 0.00000 | 0.01037 ± 0.01230 | ||
| blue lupin | 15.0 ± 22.7 | 0.00089 ± 0.00126 | 0.00974 ± 0.00721 | ||
| oil radish | 22.2 ± 25.6 | 0.00378 ± 0.00568 | 0.01191 ± 0.00899 | ||
| winter turnip rape | 9.7 ± 11.4 | 0.00189 ± 0.00201 | 0.02295 ± 0.02722 | ||
| phacelia | 4.0 ± 3.9 | 0.00222 ± 0.00329 | 0.02900 ± 0.03786 | ||
| March | crimson clover | 2.4 ± 4.2 | 0.00156 ± 0.00269 | 0.03617 ± 0.02756 ab | |
| winter rye | 1.8 ± 1.6 | 0.00167 ± 0.00145 | 0.06958 ± 0.04422 a | ||
| bristle oat | 9.6 ± 8.5 | 0.00256 ± 0.00386 | 0.01844 ± 0.01600 ab | ||
| blue lupin | 0.0 ± 0.0 | 0.00000 ± 0.00000 | 0.01228 ± 0.01104 ab | ||
| oil radish | 9.7 ± 13.4 | 0.00044 ± 0.00038 | 0.00712 ± 0.00721 b | ||
| winter turnip rape | 7.4 ± 8.0 | 0.00156 ± 0.00139 | 0.02178 ± 0.01514 ab | ||
| phacelia | 4.8 ± 8.4 | 0.00022 ± 0.00038 | 0.00553 ± 0.00361 b | ||
| 2019/2020 | October | crimson clover | 4.6 ± 8.0 b | 0.00076 ± 0.00132 b | 0.01746 ± 0.00443 c * |
| winter rye | 8.4 ± 8.5 ab | 0.01139 ± 0.01136 ab | 0.12561 ± 0.01412 b * | ||
| bristle oat | 17.5 ± 12.7 ab | 0.01006 ± 0.00599 ab | 0.05394 ± 0.02066 bc | ||
| blue lupin | 2.1 ± 2.2 b | 0.00078 ± 0.00084 b | 0.02637 ± 0.01559 c | ||
| oil radish | 14.8 ± 4.1 ab | 0.04494 ± 0.02116 a | 0.24978 ± 0.03510 a * | ||
| winter turnip rape | 11.3 ± 4.9 ab | 0.03411 ± 0.00618 a | 0.30151 ± 0.11987 a | ||
| phacelia | 31.3 ± 17.3 a | 0.05650 ± 0.03817 a | 0.10777 ± 0.03582 b | ||
| March | crimson clover | 6.1 ± 1.1 | 0.01228 ± 0.00086 ab | 0.19456 ± 0.05342 abc * | |
| winter rye | 6.2 ± 4.9 | 0.02656 ± 0.02384 ab | 0.39517 ± 0.07584 a * | ||
| bristle oat | 5.6 ± 4.4 | 0.01089 ± 0.00746 ab | 0.20400 ± 0.07336 abc * | ||
| blue lupin | 4.9 ± 1.9 | 0.00867 ± 0.00819 b | 0.14786 ± 0.09526 bc | ||
| oil radish | 21.8 ± 8.2 | 0.09406 ± 0.06253 a | 0.29670 ± 0.10020 ab | ||
| winter turnip rape | 16.3 ± 4.4 | 0.06130 ± 0.01706 ab | 0.31382 ± 0.03824 ab * | ||
| phacelia | 31.7 ± 31.3 | 0.04767 ± 0.05273 ab | 0.09699 ± 0.04619 c |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kemper, R.; Bublitz, T.A.; Müller, P.; Kautz, T.; Döring, T.F.; Athmann, M. Vertical Root Distribution of Different Cover Crops Determined with the Profile Wall Method. Agriculture 2020, 10, 503. https://doi.org/10.3390/agriculture10110503
Kemper R, Bublitz TA, Müller P, Kautz T, Döring TF, Athmann M. Vertical Root Distribution of Different Cover Crops Determined with the Profile Wall Method. Agriculture. 2020; 10(11):503. https://doi.org/10.3390/agriculture10110503
Chicago/Turabian StyleKemper, Roman, Tábata A. Bublitz, Phillip Müller, Timo Kautz, Thomas F. Döring, and Miriam Athmann. 2020. "Vertical Root Distribution of Different Cover Crops Determined with the Profile Wall Method" Agriculture 10, no. 11: 503. https://doi.org/10.3390/agriculture10110503
APA StyleKemper, R., Bublitz, T. A., Müller, P., Kautz, T., Döring, T. F., & Athmann, M. (2020). Vertical Root Distribution of Different Cover Crops Determined with the Profile Wall Method. Agriculture, 10(11), 503. https://doi.org/10.3390/agriculture10110503