Mechanism of Plant Growth Promotion and Disease Suppression by Chitosan Biopolymer

 ,
,  ,
, 


 and
and 
Abstract
:1. Introduction
2. Chitosan and Its Structural Diversity
3. Effect of Chitosan Biopolymer on Plant Growth
4. Suppression of Plant Disease by Chitosan
4.1. Antifungal Activity of CHT
4.2. Antibacterial Activity of CHT
4.3. Antiviral Activity of CHT
4.4. Nematicidal Activity of CHT
5. Mechanism of Actions of Chitosan Biopolymer
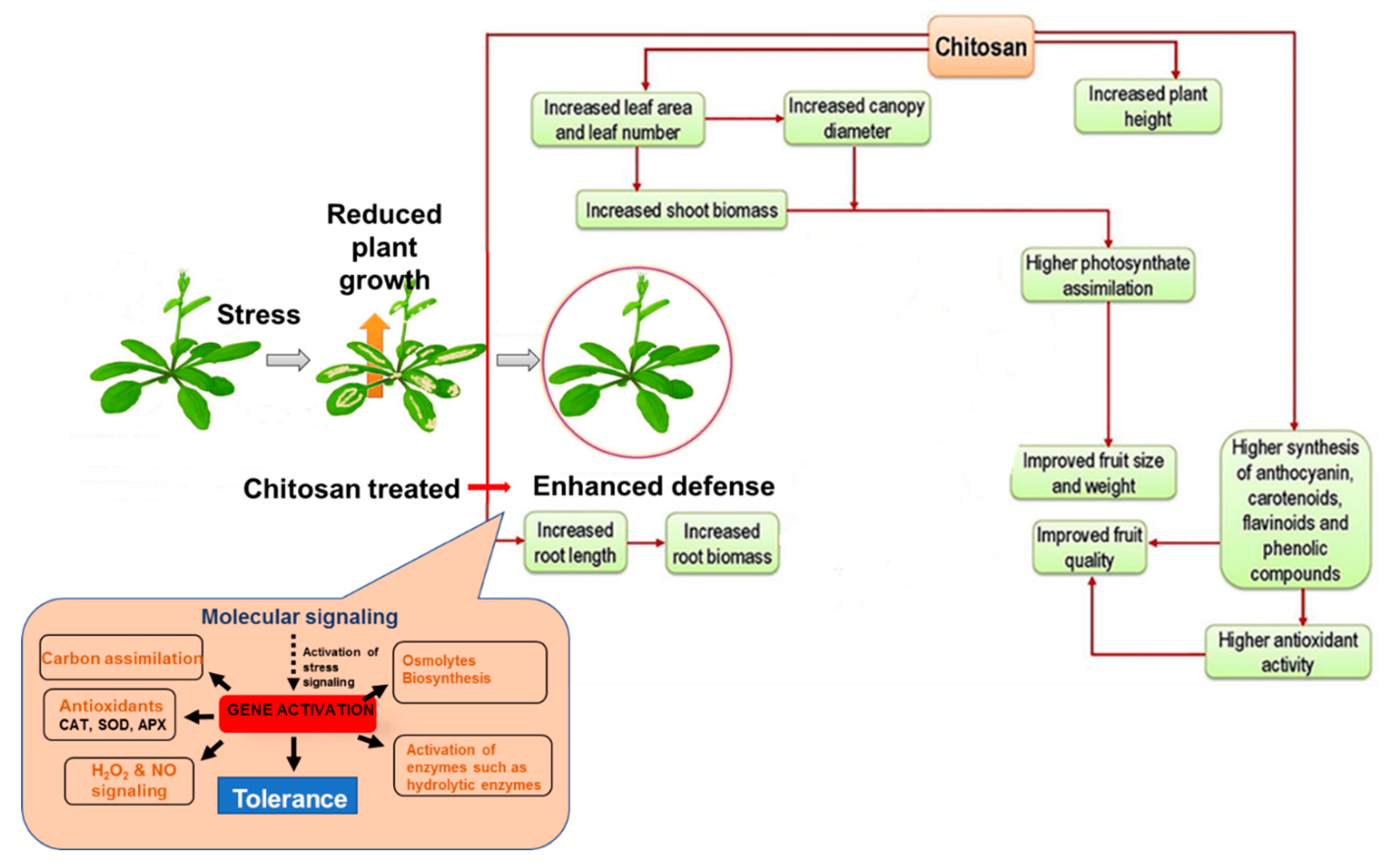
5.1. CHT as a Plant Growth Promoter
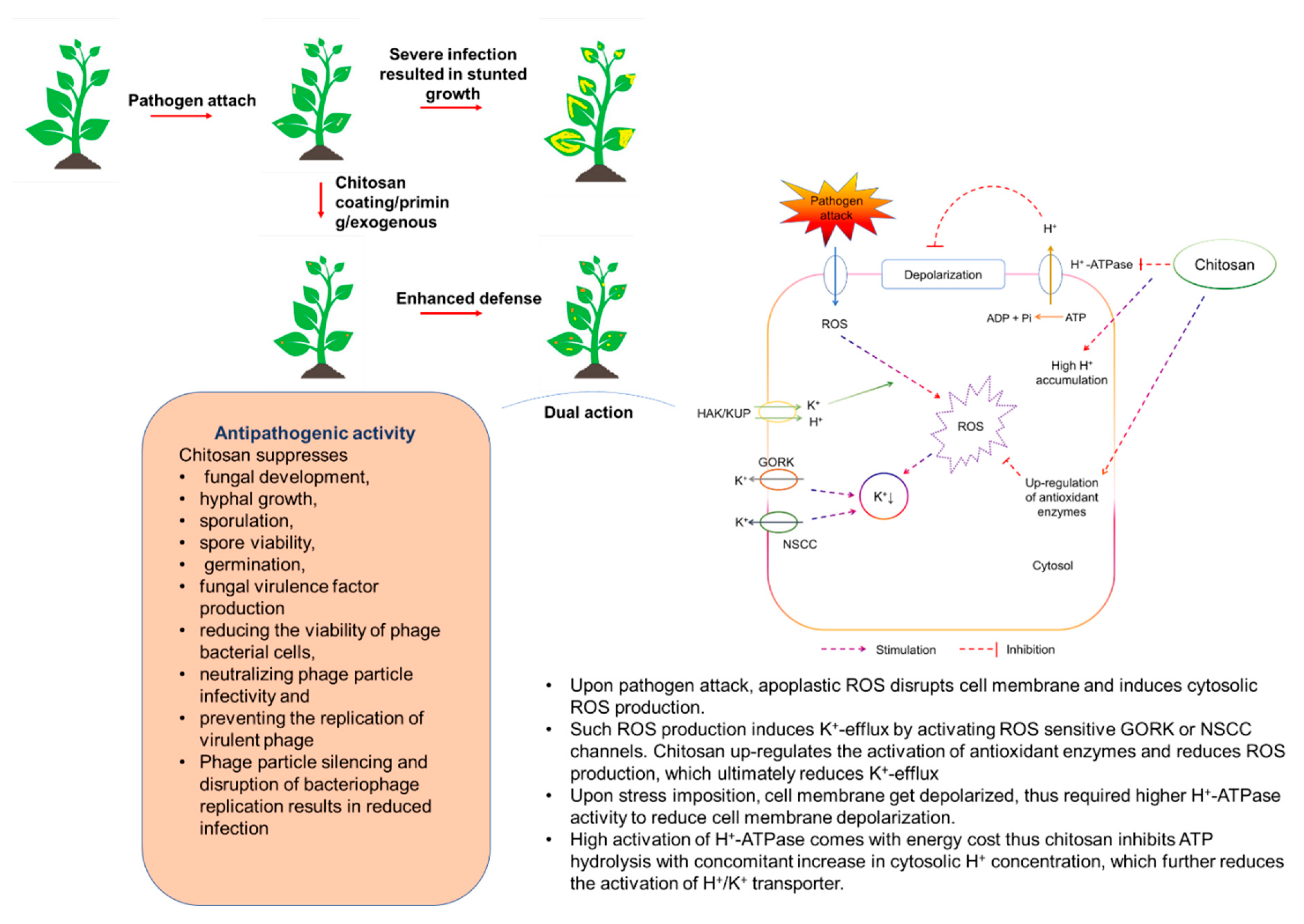
5.2. CHT as Plant Disease Suppressor
6. Concluding Remarks and Future Perspectives
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Kurita, K. Chitin and chitosan: Functional biopolymers from marine crustaceans. Mar. Biotechnol. 2006, 8, 203–226. [Google Scholar] [CrossRef]
- Braconnot, H. Sur la nature des champignons. Annu. Chem. 1811, 79, 265–304. [Google Scholar]
- Labrude, P.; Becq, C. Pharmacist and chemist Henri BraconnotLe pharmacien et chimiste Henri Braconnot. Rev. Hist. Pharm. 2003, 51, 61–78. [Google Scholar] [CrossRef]
- Hoppe-Seiler, F. Ueber chitosan und zellulose. Ber. Dtsch. Chem. Ges. 1894, 27, 3329–3331. [Google Scholar] [CrossRef]
- Anitha, A.; Sowmya, S.; Kumar, P.T.S.; Deepthi, S.; Chennazhi, K.P.; Ehrlich, H.; Tsurkan, M.; Jayakumar, R. Chitin and chitosan in selected biomedical applications. Prog. Polym. Sci. 2014, 39, 1644–1667. [Google Scholar] [CrossRef]
- Malerba, M.; Cerana, R. Chitosan effects on plant systems-A review. Int. J. Mol. Sci. 2016, 17, 996. [Google Scholar] [CrossRef] [PubMed]
- Park, Y.; Kim, M.H.; Park, S.C.; Cheong, H.; Jang, M.K.; Nah, J.W.; Hahm, K.S. Investigation of the antifungal activity and mechanism of action of LMWS-chitosan. J. Microbiol. Biotechnol. 2008, 18, 1729–1734. [Google Scholar]
- Wang, B.; Zhang, S.; Wang, X.; Yang, S.; Jiang, Q.; Xu, Y.; Xia, W. Transcriptome analysis of the effects of chitosan on the hyperlipidemia and oxidative stress in high-fat diet fed mice. Int. J. Biol. Macromol. 2017, 102, 104–110. [Google Scholar] [CrossRef]
- Sathiyabama, M.; Charles, R.E. Fungal cell wall polymer based nanoparticles in protection of tomato plants from wilt disease caused by Fusarium oxysporum f. sp. lycopersici. Carbohydr. Polym. 2015, 133, 400–407. [Google Scholar] [CrossRef]
- Sathiyabama, M.G.; Akila, R.; Einstein, C. Chitosan-induced defence responses in tomato plants against early blight disease caused by Alternaria solani (Ellis and Martin) Sorauer. Arch. Phytopathol. Plant Prot. 2014, 47, 1777–1787. [Google Scholar] [CrossRef]
- Kulikov, S.N.; Chirkov, S.N.; Ilina, A.V.; Lopatin, S.A.; Varlamov, V.P. Effect of the molecular weight of chitosan on its antiviral activity in plants. Appl. Biochem. Microbiol. 2006, 42, 200–203. [Google Scholar] [CrossRef]
- Silva, M.; Nunes, D.; Cardoso1, A.R.; Ferreiral, D.; Britol, M.; Pintadol, M.E.; Vasconcelos, M.W. Chitosan as a biocontrol agent against the pinewood nematode (Bursaphelenchus xylophilus). For. Pathol. 2014, 44, 420–423. [Google Scholar] [CrossRef]
- Sun, D.; Liang, G.; Xie, J.; Lei, X.; Mo, Y. Improved preservation effects of litchi fruit by combining chitosan coating with ascorbic acid treatment during postharvest storage. Afr. J. Biotechnol. 2010, 9, 3272–3279. [Google Scholar]
- Rahman, M.; Mukta, J.A.; Sabir, A.A.; Gupta, D.R.; Mohi-ud-din, M.; Hasanuzzaman, M.; Miah, M.G.; Rahman, M.; Islam, M.T. Chitosan biopolymer promotes yield and stimulates accumulation of antioxidants in strawberry fruit. PLoS ONE 2018, 13, e0203769. [Google Scholar] [CrossRef] [PubMed]
- Emami Bistgani, Z.; Siadat, S.A.; Bakhshandeh, A.; Ghasemi Pirbalouti, A.; Hashemi, M. Interactive effects of drought stress and chitosan application on physiological characteristics and essential oil yield of Thymus daenensis Celak. Crop J. 2017, 5, 407–415. [Google Scholar] [CrossRef]
- Xoca-Orozco, L.-Á.; Cuellar-Torres, E.A.; González-Morales, S.; Gutiérrez-Martínez, P.; López-García, U.; Herrera-Estrella, L.; Vega-Arreguín, J.; Chacón-López, A. Transcriptomic analysis of avocado hass (Persea americana Mill) in the interaction system fruit-chitosan-Colletotrichum. Front. Plant Sci. 2017, 8, 956. [Google Scholar] [CrossRef] [Green Version]
- Guan, Y.; Hu, J.; Wang, X.; Shao, C. Seed priming with chitosan improves maize germination and seedling growth in relation to physiological changes under low temperature stress. J. Zhejiang Univ. Sci. B 2009, 10, 427–433. [Google Scholar] [CrossRef] [Green Version]
- Muzzarelli, R.A. Natural Chelating Polymers; Alginic Acid, Chitin and Chitosan. In Natural Chelating Polymers; Alginic Acid, Chitin and Chitosan; Pergamon Press: Oxford, UK, 1973. [Google Scholar]
- Rinaudo, M. Chitin and chitosan: Properties and applications. Prog. Polym. Sci. 2006, 31, 603–632. [Google Scholar] [CrossRef]
- Shahidi, F.; Abuzaytoun, R. Chitin, chitosan, and co-products: Chemistry, production, applications, and health effects. Adv. Food Nutr. Res. 2005, 49, 93–135. [Google Scholar]
- Sarmento, B.; Francisco, M.; Goycoolea, F.M.; Sosnik, A.; das Neves, J. Chitosan and chitosan derivatives for biological applications: Chemistry and functionalization. Int. J. Carbohy. Chem. 2011, 802693. [Google Scholar] [CrossRef] [Green Version]
- Aljbour, N.D.; Beg, M.D.H.; Gimbun, J. Acid hydrolysis of chitosan to oligomers using hydrochloric acid. Chem. Eng. Technol. 2019, 42, 1741–1746. [Google Scholar] [CrossRef]
- Ma, Z.; Wang, W.; Wu, Y.; He, Y.; Wu, T. Oxidative degradation of chitosan to the low molecular water-soluble chitosan over peroxotungstate as chemical scissors. PLoS ONE 2014, 9, e100743. [Google Scholar] [CrossRef] [PubMed]
- Kaczmarek, M.B.; Struszczyk-Swita, K.; Li, X.; Szczęsna-Antczak, M.; Daroch, M. Enzymatic modifications of chitin, chitosan, and chitooligosaccharides. Front. Bioeng. Biotechnol. 2019, 7, 243. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kasaai, M.R.; Arul, J.; Charlet, G. Fragmentation of chitosan by ultrasonic irradiation. Ultrason. Sonochem. 2008, 15, 1001–1008. [Google Scholar] [CrossRef] [PubMed]
- Dowling, M.B.; Kumar, R.; Keibler, M.A.; Hess, J.R.; Bochicchio, G.V.; Raghavan, S.R. A self-assembling hydrophobically modified chitosan capable of reversible hemostatic action. Biomaterials 2011, 32, 3351–3357. [Google Scholar] [CrossRef] [PubMed]
- Park, J.H.; Kwon, S.; Lee, M.; Chung, H.; Kim, J.H.; Kim, Y.S.; Park, R.W.; Kim, I.S.; Seo, S.B.; Kwon, I.C.; et al. Self-assembled nanoparticles based on glycol chitosan bearing hydrophobic moieties as carriers for doxorubicin: In vivo biodistribution and antitumor activity. Biomaterials 2006, 27, 119–126. [Google Scholar] [CrossRef] [PubMed]
- Zargar, V.; Asghari, M.; Dashti, A. A review on chitin and chitosan polymers: Structure, chemistry, solubility, derivatives, and applications. ChemBioEng Rev. 2015, 2, 204–226. [Google Scholar] [CrossRef]
- Kafedjiiski, K.; Krauland, A.H.; Hoffer, M.H.A. Bernkop-Schnurch, synthesis and in vitro evaluation of a novel thiolated chitosan. Biomaterials 2005, 26, 819–826. [Google Scholar] [CrossRef]
- Khanjari, A.; Karabagias, I.; Kontominas, M. Combined effect of N, O-carboxymethyl chitosan and oregano essential oil to extend shelf life and control Listeria monocytogenes in raw chicken meat fillets. LWT Food Sci. Technol. 2013, 53, 94–99. [Google Scholar] [CrossRef]
- Yin, L.; Fei, L.; Cui, F.; Tang, C.; Yin, C. Superporous hydrogels containing poly (acrylic acidco-acrylamide)/O-carboxymethyl chitosan interpenetrating polymer networks. Biomaterials 2007, 28, 1258–1266. [Google Scholar] [CrossRef]
- Xiao, B.; Wan, Y.; Wang, X.; Zha, Q.; Liu, H.; Qiu, Z.; Zhang, S. Synthesis and characterization of N-(2-hydroxy)propyl-3-trimethyl ammonium chitosan chloride for potential application in gene delivery. Colloids Surf. B Biointerfaces 2012, 91, 168–174. [Google Scholar] [CrossRef] [PubMed]
- Sajomsang, W. Synthetic methods and applications of chitosan containing pyridylmethyl moiety and its quaternized derivatives: A review. Carbohydr. Polym. 2010, 80, 631–647. [Google Scholar] [CrossRef]
- Xing, R.; Yu, H.; Liu, S.; Zhang, W.; Zhang, Q.; Li, Z.; Li, P. Antioxidant activity of differently regioselective chitosan sulfates in vitro. Bioorg. Med. Chem. 2005, 13, 1387–1392. [Google Scholar] [CrossRef] [PubMed]
- Abd-Alla, M.; Wafaa, M. New safe methods for controlling anthracnose disease of mango (Mangifera indica L.) fruits caused by Colletotrichum gloeosporioides (Penz.). J. Am. Sci. 2010, 8, 361–367. [Google Scholar]
- Morimoto, M.; Saimoto, H.; Usui, H.; Okamoto, Y.; Minami, S.; Shigemasa, Y. Biological activities of carbohydrate-branched chitosan derivatives. Biomacromolecules 2001, 2, 1133–1136. [Google Scholar] [CrossRef]
- Prabaharan, M.; Mano, J.F. Chitosan derivatives bearing cyclodextrin cavities as novel adsorbent matrices. Carbohydr. Polym. 2006, 63, 153–166. [Google Scholar] [CrossRef]
- Kumar, G.; Smith, P.J.; Payne, G.F. Enzymatic grafting of a natural product onto chitosan to confer water solubility under basic conditions. Biotechnol. Bioeng. 1999, 63, 154–165. [Google Scholar] [CrossRef]
- Park, I.K.; Park, Y.H.; Shin, B.A.; Choi, E.S.; Kim, Y.R.; Akaike, T.; Cho, C.S. Galactosylated chitosan-graft-dextran as hepatocyte-targeting DNA carrier. J. Control. Release 2000, 69, 97–108. [Google Scholar] [CrossRef]
- Mahdavinia, G.R.; Pourjavadi, A.; Hosseinzadeh, H.; Zohuriaan, M.J. Modified chitosan 4. Superabsorbent hydrogels from poly (acrylic acid-co-acrylamide) grafted chitosan with salt-and pH-responsiveness properties. Eur. Polym. J. 2004, 40, 1399–1407. [Google Scholar] [CrossRef]
- Prashanth, K.V.H.; Tharanathan, R.N. Chitin/chitosan: Modifications and their unlimited application potential – An overview. Trends. Food Sci. Technol. 2007, 18, 117–131. [Google Scholar] [CrossRef]
- Chandrkrachang, S. The applications of chitin in agriculture in Thailand. Adv. Chitin Sci. 2002, 5, 458–462. [Google Scholar]
- Pornpeanpakdee, P.; Pichyangkura, R.; Chadchawan, S.; Limpanavech, P. Chitosan effects on Dendrobium ‘Eiskul’ Protocorm-like body production. In Proceedings of the 31st Congress on Science and Technology of Thailand, Nakornrachaseema, Thailand, 18–20 October 2005; pp. 1–3. [Google Scholar]
- Nahar, S.J.; Kazuhiko, S.; Haque, S.M. Effect of Polysaccharides Including Elicitors on Organogenesis in Protocorm-like Body (PLB) of Cymbidium insigne in vitro. J. Agric. Sci. Technol. 2012, 2, 1029–1033. [Google Scholar]
- Barka, E.A.; Eullaffroy, P.; Clément, C.; Vernet, G. Chitosan improves development, and protects Vitis vinifera L. against Botrytis cinerea. Plant Cell Rep. 2004, 22, 608–614. [Google Scholar] [CrossRef] [PubMed]
- Sauerwein, M.; Flores, H.M.; Yamazaki, T.; Shimomura, K. Lippia dulcis shoot cultures as a source of the sweet sesquiterpene hernandulcin. Plant Cell Rep. 1991, 9, 663–666. [Google Scholar] [CrossRef] [PubMed]
- Tsugita, T.; Takahashi, K.; Muraoka, T.; Fukui, H. The application of chitin/chitosan for agriculture (in Japanese). In Proceedings of the Special Session of the 7th Symposium on Chitin and Chitosan, Fukui, Japan, 6 March 1993; Japanese Society for Chitin and Chitosan: Fukui, Japan, 1993; pp. 21–22. [Google Scholar]
- Spiegel, Y.; Kafkafi, U.; Pressman, E. Evaluation of a protein-chitin derivative of crustacean shells as a slow-release nitrogen fertilizer on Chinese cabbage. J. Hortic. Sci. 1988, 63, 621–628. [Google Scholar] [CrossRef]
- Lee, Y.S.; Kim, Y.H.; Kim, S.B. Changes in the respiration, growth, and vitamin C content of soybean sprouts in response to chitosan of different molecular weights. Hortic. Sci. 2005, 40, 1333–1335. [Google Scholar] [CrossRef] [Green Version]
- Pirbalouti, A.G.; Malekpoor, F.; Salimi, A.; Golparvar, A. Exogenous application of chitosan on biochemical and physiological characteristics, phenolic content and antioxidant activity of two species of basil (Ocimum ciliatum and Ocimum basilicum) under reduced irrigation. Sci. Hortic. 2017, 217, 114–122. [Google Scholar] [CrossRef]
- Wanichpongpan, P.; Suriyachan, K.; Chandrkrachang, S. Effects of Chitosan on the growth of Gerbera flower plant (Gerbera jamesonii). In Chitin and Chitosan in Life Science, Proceedings of the Eighth International Chitin and Chitosan Conference and Fourth Asia Pacific Chitin and Chitosan Symposium, Yamaguchi, Japan, 21–23 September 2000; Uragami, T., Kurita, K., Fukamizo, T., Eds.; Kodansha Scientific: Tokyo, Japan, 2001; pp. 198–201. [Google Scholar]
- Choudhary, R.C.; Kumaraswamy, R.V.; Kumari, S.; Sharma, S.S.; Pal, A.; Raliya, R.; Biswas, P.; Saharan, V. Cu-chitosan nanoparticle boost defense responses and plant growth in maize (Zea mays L.). Sci. Rep. 2017, 7, 9754–9765. [Google Scholar] [CrossRef]
- Salachna, P.; Zawadzinska, A. Effect of chitosan on plant growth, flowering and corms yield of potted freesia. J. Ecol. Eng. 2014, 15, 93–102. [Google Scholar]
- Vasudevan, P.; Reddy, M.S.; Kavitha, S.; Velusamy, P.; Paul Raj, R.S.D.; Priyadarisini, V.B.; Bharathkumar, S.; Kloepper, J.W.; Gnanamanickam, S.S. Role of biological preparations in enhancement of rice seedling growth and seed yield. Curr. Sci. 2002, 83, 1140–1143. [Google Scholar]
- Phothi, R.; Theerakarunwong, C.D. Effect of chitosan on physiology, photosynthesis and biomass of rice (Oryza sativa L.) under elevated ozone. Aust. J. Crop Sci. 2017, 11, 624–630. [Google Scholar] [CrossRef]
- Chibu, H.; Shibayama, H.; Arima, S. Effects of chitosan application on the shoot growth of rice and soybean. Jpn. J. Crop Sci. 2002, 71, 206–211. [Google Scholar] [CrossRef] [Green Version]
- Zong, H.; Liu, S.; Xing, R.; Chen, X.; Li, P. Protective effect of chitosan on photosynthesis and antioxidative defense system in edible rape (Brassica rapa L.) in the presence of cadmium. Ecotoxicol. Environ. Saf. 2017, 138, 271–278. [Google Scholar] [CrossRef]
- Shao, C.X.; Hu, J.; Song, W.J.; Hu, W.M. Effects of seed priming with chitosan solutions of different acidity on seed germination and physiological characteristics of maize seedling. Agric. Life Sci. 2005, 31, 705–708. [Google Scholar]
- Saharan, V.; Kumaraswamy, R.; Choudhary, R.C.; Kumari, S.; Pal, A.; Raliya, R.; Biswas, P.J. Cu-Chitosan nanoparticle mediated sustainable approach to enhance seedling growth in maize by mobilizing reserved food. Agric. Food Chem. 2016, 64, 6148–6155. [Google Scholar] [CrossRef] [PubMed]
- Falcón-Rodríguez, A.B.; Costales, D.; Gónzalez-Peña, D.; Morales, D.; Mederos, Y.; Jerez, E.; Cabrera, J.C. Chitosans of different molecular weight enhance potato (Solanum tuberosum L.) yield in a field trial. Span. J. Agric. Res. 2017, 15, e0902. [Google Scholar]
- Kowalski, B.; Jimenez Terry, F.; Herrera, L.; Agramonte Peñalver, D. Application of soluble chitosan in vitro and in the greenhouse to increase yield and seed quality of potato minitubers. Potato Res. 2006, 49, 167–176. [Google Scholar] [CrossRef]
- Saharan, V.; Sharma, G.; Yadav, M.; Choudhary, M.K.; Sharma, S.; Pal, A.; Raliya, R.; Biswas, P. Synthesis and in vitro antifungal efficacy of Cu–chitosan nanoparticles against pathogenic fungi of tomato. Int. J. Biol. Macromol. 2015, 75, 346–353. [Google Scholar] [CrossRef]
- Mondal, M.M.A.; Malek, M.A.; Puteh, A.B.; Ismail, M.R.; Ashrafuzzaman, M.; Naher, L. Effect of foliar application of chitosan on growth and yield in okra. Aust. J. Crop Sci. 2012, 6, 918–921. [Google Scholar]
- Sultana, S.; Islam, M.; Khatun, M.A.; Hassain, M.A.; Huque, R. Effect of Foliar Application of Oligo-chitosan on growth, yield and quality of tomato and eggplant. Asian J. Agric. Res. 2017, 11, 36–42. [Google Scholar]
- Pereira, A.E.S.; Silva, P.M.; Oliveira, J.L.; Oliveira, H.C.; Fraceto, L.F. Chitosan nanoparticles as carrier systems for the plant growth hormone gibberellic acid. Colloids Surf. B Biointerfaces 2017, 150, 141–152. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Akter, J.; Jannat, R.; Hossain, M.M.; Ahmed, J.U.; Rubayet, M.T. Chitosan for plant growth promotion and disease suppression against anthracnose in chilli. Int. J. Agric. Environ. Biotechnol. 2018, 3, 806–817. [Google Scholar] [CrossRef] [Green Version]
- Chookhongkha, N.; Miyagawa, S.; Jirakiattikul, Y.; Photchanachai, S. Chili growth and seed productivity as affected by chitosan. In Proceedings of the International Conference on Agriculture Technology and Food Sciences (ICATFS’2012), Manila, Philippines, 17–18 November 2012; pp. 17–18. [Google Scholar]
- Mahmood, N.; Abbasi, N.A.; Hafiz, I.A.; Ali, I.; Zakia, S. Effect of biostimulants on growth, yield and quality of bell pepper cv. Yolo wonder. Pak. J. Agric. Sci. 2017, 54, 311–317. [Google Scholar]
- Anusuya, S.; Sathiyabama, M. Effect of chitosan on growth, yield and curcumin content in turmeric under field condition. Biocatal. Agric. Biotechnol. 2016, 6, 102–106. [Google Scholar] [CrossRef]
- Batool, M.; Asghar, R. Seed priming with chitosan improves the germination and growth performance of ajowan (Carum copticum) under salt stress. Eurasia J. Biosci. 2013, 7, 69–76. [Google Scholar]
- Ziani, K.; Ursúa, B.; Maté, J.I. Application of bioactive coatings based on chitosan for artichoke seed protection. Crop Prot. 2010, 29, 853–859. [Google Scholar] [CrossRef]
- Shehata, S.; Fawzy, Z.; El-Ramady, H. Response of cucumber plants to foliar application of chitosan and yeast under greenhouse conditions. Aust. J. Basic Appl. Sci. 2012, 6, 63–71. [Google Scholar]
- Mahdavi, B.; Safari, H. Effect of chitosan on growth and some physiological characteristics of chickpea under salinity stress condition. J. Plant Process Funct. 2015, 4, 117–127. [Google Scholar]
- Anusuya, S.; Nibiya Banu, K. Silver-chitosan nanoparticles induced biochemical variations of chickpea (Cicer arietinum L.). Biocatal. Agric. Biotechnol. 2016, 8, 39–44. [Google Scholar] [CrossRef]
- Van, S.N.; Minh, H.D.; Anh, D.N. Study on chitosan nanoparticles on biophysical characteristics and growth of Robusta coffee in green house. Biocatal. Agric. Biotechnol. 2013, 2, 289–294. [Google Scholar]
- Mutka, J.A.; Rahman, M.; Sabir, A.A.; Gupta, D.R.; Surovy, M.Z.; Rahman, M.; Islam, M.T. Chitosan and plant probiotics application enhance growth and yield of strawberry. Biocatal. Agric. Biotechnol. 2017, 11, 9–18. [Google Scholar]
- González Gómez, H.; Ramírez Godina, F.; Ortega Ortiz, H.; Benavides Mendoza, A.; Robledo Torres, V.; Cabrera De la Fuente, M. Use of chitosan-PVA hydrogels with copper nanoparticles to improve the growth of grafted watermelon. Molecules 2017, 22, 1031. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zagzog, O.A.; Gad, M.M.; Hafez, N.K. Effect of nano-chitosan on vegetative growth, fruiting and resistance of malformation of mango. Trends Hortic. Res. 2017, 6, 673–681. [Google Scholar]
- Sauerwein, M.; Yamazaki, T.; Shimomura, K. Hernandulcin in hairy root cultures of Lippia dulcis. Plant Cell Rep. 1991, 9, 579–581. [Google Scholar] [CrossRef]
- Irawati, E.B.; Sasmita, E.R.; Suryawati, A. Application of chitosan for vegetative growth of kemiri sunan plant in marginal land. IOP Conf. Ser. Earth Environ. Sci. 2019, 250, 012089. [Google Scholar] [CrossRef]
- Trzcinska, A.; Bogusiewic, A.; Szkop, M.; Drozdowski, S. Effect of chitosan on disease control and growth of scots pine (Pinus sylvestris L.) in a forest nursery. Forest 2015, 6, 3165–3176. [Google Scholar] [CrossRef] [Green Version]
- Orzali, L.; Corsi, B.; Forni, C.; Riccioni, L. Chitosan in agriculture: A new challenge for managing plant disease. In Biological Activities and Application of Marine Polysaccharides; Shalaby, E.A., Ed.; IntechOpen: London, UK, 2017. [Google Scholar]
- Andrews, J.M. Determination of minimum inhibitory concentrations. J. Antimicr. Chemoth. 2001, 48, 5. [Google Scholar] [CrossRef] [Green Version]
- Allan, C.R.; Hadwiger, L.A. The fungicidal effect of chitosan on fungi of varying cell wall composition. Exp. Mycol. 1979, 3, 285–287. [Google Scholar] [CrossRef]
- Muzzarelli, R.A.A.; Tarsi, R.; Filippini, O.; Giovanetti, E.; Biagini, G.; Varaldo, P.E. Antimicrobial properties of N-carboxybutyl chitosan. Antimicrob. Agents Chemother. 1990, 34, 2019–2023. [Google Scholar] [CrossRef] [Green Version]
- Vasyukova, N.I.; Chalenko, G.I.; Gerasimova, N.G.; Perekhod, E.A.; Ozeretskovskaya, O.L.; Irina, A.V.; Varlamov, V.P.; Albulov, A.I. Chitin and chitosan derivatives as elicitors of potato resistance to late blight. Appl. Biochem. Microbiol. 2005, 36, 372–376. [Google Scholar] [CrossRef]
- Badawy, M.E.I.; Rabea, E.I. A biopolymer chitosan and its derivatives as promising antimicrobial agents against plant pathogens and their applications in crop protection. Int. J. Carbohydr. Chem. 2011, 2011, 460381. [Google Scholar] [CrossRef]
- El Ghaouth, A.; Arul, J.; Wilson, C.; Benhamou, N. Ultrastructural and cytochemical aspects of the effect of chitosan on decay of bell pepper fruit. Physiol. Mol. Plant Pathol. 1994, 44, 417–432. [Google Scholar] [CrossRef]
- Xu, J.; Zhao, X.; Han, X.; Du, Y. Antifungal activity of oligochitosan against Phytophthora capsici and other plant pathogenic fungi in vitro. Pestic. Biochem. Physiol. 2007, 87, 220–228. [Google Scholar] [CrossRef]
- El-Mohamedya, R.S.R.; Abd El-Aziz, M.E.; Kamel, S. Antifungal activity of chitosan nanoparticles against some plant pathogenic fungi in vitro. Agric. Eng. Int. CIGR J. 2019, 21, 201–209. [Google Scholar]
- Berger, L.R.R.; Stanford, N.P.; Walladino, L.G.; Laranieira, D.; Barbosa de-Lima, M.A.; Malheiros, S.M.M.; Oliveria, W.J.D.; Stamford, T.C.M. Cowpea resistance induced against Fusarium oxysporum f. sp. tracheiphilum by crustaceous chitosan and by biomass and chitosan obtained from Cunninghamella elegans. Biol. Cont. 2016, 92, 45–54. [Google Scholar] [CrossRef]
- Hassni, M.; El Hadrami, A.; Daayf, F.; Barka, E.A.; El Hadrami, I. Chitosan, antifungal product against Fusarium oxysporum f. sp. albedinis and elicitor of defence reactions in date palm roots. Phytopathol. Mediterr. 2004, 43, 195–204. [Google Scholar]
- Dos Santos, N.S.T.; Aguiar, A.J.A.A.; de Oliveira, C.E.V.; de Sales, C.V.; de Melo e Silva, S.; da Silva, R.S.; Stamford, T.C.M.; de Souza, E.L. Efficacy of the application of a coating composed of chitosan and Origanum vulgare L. essential oil to control Rhizopus stolonifer and Aspergillus niger in grapes (Vitis labrusca L.). Food Microbiol. 2012, 32, 345–353. [Google Scholar] [CrossRef]
- Gutierrez-Martinez, P.; Ledezma-Morales, A.; Romero-Islas, L.d.L.; Ramos-Guerrero, A.; Romero-Islas, J.; Rodríguez-Pereida, C.; Casas-Junco, P.; Coronado-Partida, L.; González-Estrada, R. Antifungal activity of chitosan against postharvest fungi of tropical and subtropical fruits. In Chitin-Chitosan: Myriad Functionalities in Science and Technology; Dongre, R.S., Ed.; IntechOpen: London, UK, 2018. [Google Scholar]
- Kanetis, L.; Exarchou, V.; Charalambous, Z.; Goulas, V. Edible coating composed of chitosan and Salvia fruticosa Mill. extract for the control of grey mould of table grapes. J. Sci. Food Agric. 2017, 97, 452–460. [Google Scholar] [CrossRef]
- Falcon, A.B.; Cabrera, J.C.; Costales, D.; Ramirez, M.A.; Cabrera, G.; Toledo, V.; Martinez-Tellez, M.A. The effect of size and acetylation degree of chitosan derivatives on tobacco plant protection against Phytophthora parasitica nicotianae. World J. Microbiol. Biotechnol. 2008, 24, 103–112. [Google Scholar] [CrossRef] [Green Version]
- Kiprushkina, E.I.; Shestopalova, I.A.; Pekhotina, A.M.; Kuprina, E.E.; Nikitina, O.V. Protective-stimulating properties of chitosan in the vegetation and storing tomatoes. Prog. Chem. Appl. Chitin Deriv. 2017, 23, 77–81. [Google Scholar] [CrossRef] [Green Version]
- Guerra, I.C.D.; de Oliveira, P.D.L.; de Souza Pontes, A.L.; Lúcio, A.S.S.C.; Tavares, J.F.; Barbosa-Filho, J.M.; Madruga, M.S.; de Souza, E.L. Coatings comprising chitosan and Mentha piperita L. or Mentha×villosa Huds essential oils to prevent common postharvest mold infections and maintain the quality of cherry tomato fruit. Int. J. Food Microbiol. 2015, 214, 168–178. [Google Scholar] [CrossRef] [PubMed]
- Ing, L.Y.; Zin, N.M.; Sarwar, A.; Katas, H. Antifungal activity of chitosan nanoparticles and correlation with their physical properties. Int. J. Biomater. 2012, 2012, 632698. [Google Scholar] [CrossRef] [PubMed]
- Chowdappa, P.; Gowda, S.; Chethana, C.S.; Madhura, S. Antifungal activity of chitosan-silver nanoparticle composite against Colletotrichum gloeosporioides associated with mango anthracnose. Afr. J. Microbiol. Res. 2014, 8, 1803–1812. [Google Scholar]
- Cuero, R.G.; Duffus, E.; Osuji, G.; Pettit, R. Aflatoxin control in preharvest maize: Effects of chitosan and two microbial agents. J. Agric. Sci. 1991, 117, 165–169. [Google Scholar] [CrossRef]
- Meng, X.; Yang, L.; Kennedy, J.F.; Tian, S. Effects of chitosan and oligochitosan on growth of two fungal pathogens and physiological properties in pear fruit. Carbohydr. Polym. 2010, 81, 70–75. [Google Scholar] [CrossRef]
- Ma, Z.; Yang, L.; Yan, H.; Kennedy, J.F.; Meng, X. Chitosan and oligochitosan enhance the resistance of peach fruit to brown rot. Carbohydr. Polym. 2013, 94, 272–277. [Google Scholar] [CrossRef]
- Maia, A.J.; Botelho, R.V.; Faria, C.M.D.R.; Leite, C.D. Chitosan action on Plasmopara viticola and Elsinoe ampelina development in vitro and in grapevines cv. Isabel. Sum. Phytopathol. 2010, 36, 203–209. [Google Scholar] [CrossRef] [Green Version]
- Manjunatha, G.; Roopa, K.S.; Prashanth, G.N.; Shetty, H.S. Chitosan enhances disease resistance in pearl millet against downy mildew caused by Sclerospora graminicola and defence-related enzyme activation. Pest Manag. Sci. 2008, 64, 1250–1257. [Google Scholar] [CrossRef]
- Liu, H.; Tian, W.; Li, B.; Wu, G.; Ibrahim, M.; Tao, Z.; Wang, Y.; Xie, G.; Li, H.; Sun, G. Antifungal effect and mechanism of chitosan against the rice sheath blight pathogen, Rhizoctonia solani. Biotechnol. Lett. 2012, 34, 2291–2298. [Google Scholar] [CrossRef]
- Hadwiger, L.A.; McBride, P.O. Low-level copper plus chitosan applications provide protection against late blight of potato. Plant Health Prog. 2006, 7, 22. [Google Scholar] [CrossRef] [Green Version]
- Benhamou, N.; Theriault, G. Treatment with chitosan enhances resistance of tomato plants to the crown and root rot pathogen Fusarium oxysporum f. sp. radicis-lycopersici. Physiol. Mol. Plant Pathol. 1992, 41, 33–52. [Google Scholar] [CrossRef]
- Reglinski, T.; Elmer, P.A.G.; Taylor, J.T.; Wood, P.N.; Hoyte, S.M. Inhibition of Botrytis cinerea growth and suppression of botrytis bunch rot in grapes using chitosan. Plant Pathol. 2010, 59, 882–890. [Google Scholar] [CrossRef] [Green Version]
- Feliziani, E.; Landi, L.; Romanazzi, G. Preharvest treatments with chitosan and other alternatives to conventional fungicides to control postharvest decay of strawberry. Carbohydr. Polym. 2015, 132, 111–117. [Google Scholar] [CrossRef] [PubMed]
- Zahid, N.; Maqbool, M.; Siddiqui, Y.; Manickam, S.; Ali, A. Regulation of inducible enzymes and suppression of anthracnose using submicron chitosan dispersions. Sci. Hortic. 2015, 193, 381–388. [Google Scholar] [CrossRef]
- Sathiyabama, M.; Balasubramanian, R. Chitosan induces resistance components in Arachis hypogaea against leaf rust caused by Puccinia arachidis Speg. Crop Protec. 1998, 17, 307–313. [Google Scholar] [CrossRef]
- Dodgson, J.L.A.; Dodgson, W. Comparison of effects of chitin and chitosan for control of Colletotrichum sp. on cucumbers. J. Pure Appl. Microbiol. 2017, 11, 87–93. [Google Scholar] [CrossRef]
- Lopez-Moya, F.; Martin-Urdiroz, M.; Oses-Ruiz, M.; Fricker, M.D.; Littlejohn, G.R.; Lopez-Llorca, L.V.; Talbot, N.J. Chitosan inhibits septin-mediated plant infection by the rice blast fungus Magnaporthe oryzae in a protein kinase C and Nox1 NADPH oxidase-dependent manner. bioRxiv 2020. [Google Scholar] [CrossRef]
- Boonlertnirun, S.; Boonraung, C.; Suvanasara, R. Application of chitosan in rice production. J. Met. Mater. Miner. 2008, 18, 47–52. [Google Scholar]
- Chatterjee, S.; Chatterjee, B.P.; Guha, A.K. A study an antifungal activity of water-soluble chitosan against Macrophomina phaseolina. Int. J. Biol. Macromol. 2014, 67, 452–457. [Google Scholar] [CrossRef]
- Kheiri, A.; Moosawi Jorf, S.A.; Malihipour, A.; Saremi, H.; Nikkhah, A. Synthesis and characterization of chitosan nanoparticles and their effect on Fusarium head blight and oxidative activity in wheat. Int. J. Biol. Macromol. 2017, 102, 526–538. [Google Scholar] [CrossRef]
- Bhaskara Reddy, M.V.; Arul, J.; Angers, P.; Couture, L. Chitosan treatment of wheat seeds induces resistance to Fusarium graminearum and improves seed quality. J. Agric. Food Chem. 1999, 47, 1208–1216. [Google Scholar] [CrossRef] [PubMed]
- Hassan, G.M.; Xiang, M.; Liu, X. Synergetic suppression of soybean cyst nematodes by chitosan and Hirsutella minnesotensis via the assembly of the soybean rhizosphere microbial communities. Biol. Control 2017, 115, 86–94. [Google Scholar]
- Benhamou, N.; Lafontaine, P.J.; Nicole, M. Induction of systemic resistance to Fusarium crown and root rot in tomato plants by seed treatment with chitosan. Phytopathology 1994, 84, 1432–1444. [Google Scholar] [CrossRef]
- Lafontaine, J.P.; Benhamou, N. Chitosan treatment: An emerging strategy for enhancing resistance of greenhouse tomato plants to infection by Fusarium oxysporum f. sp. radicislycopersici. Biocontrol Sci. Technol. 1996, 6, 111–124. [Google Scholar] [CrossRef]
- O’Herlihy, E.A.; Duffy, E.M.; Cassells, A.C. The effects of arbuscular mycorrhizal fungi band chitosan sprays on yield and late blight resistance in potato crops from microplants. Folia Geobot. 2003, 38, 201–208. [Google Scholar] [CrossRef]
- El-Mohamedy, R.S.R.; Shafeek, M.R.; Abd El-Samad, E.E.-D.H.; Salama, D.M.; Rizk, F.A. Field application of plant resistance inducers (PRIs) to control important root rot diseases and improvement growth and yield of green bean (Phaseolus vulgaris L.). Aust. J. Crop Sci. 2017, 11, 496–505. [Google Scholar] [CrossRef]
- Ben-Shalom, N.; Fallik, E. Further suppression of Botrytis cinerea disease in cucumber seedlings by chitosan-copper complex as compared with chitosan alone. Phytoparasitica 2003, 31, 99–102. [Google Scholar] [CrossRef]
- Moret, A.; Muñoz, Z.; Garcés, S. Control of powdery mildew on cucumber cotyledons by chitosan. J. Plant Pathol. 2009, 1, 375–380. [Google Scholar]
- Zohara, F.; Surovy, M.Z.; Khatun, A.; Prince, M.F.R.K.; Akanda, M.A.M.; Rahman, M.; Islam, M.T. Chitosan biostimulant controls infection of cucumber by Phytophthora capsici through suppression of asexual reproduction of the pathogen. Acta Agrobot. 2019, 72, 1763. [Google Scholar] [CrossRef]
- Long, L.T.; Tan, L.V.; Boi, V.N.; Trung, T.S. Antifungal activity of water-soluble chitosan against Colletotrichum capsici in post-harvest chili pepper. J. Food Process. Preserv. 2018, 42, e13339. [Google Scholar] [CrossRef]
- Sid Ahmed, A.; Ezziyyani, M.; Pérez Sánchez, C.; Candela, M.E. Effect of chitin on biological control activity of Bacillus spp. and Trichoderma harzianum against root rot disease in pepper (Capsicum annuum) plants. Eur. J. Plant Pathol. 2003, 109, 633–637. [Google Scholar] [CrossRef]
- Mandal, S. Induction of phenolics, lignin and key defense enzymes in eggplant (Solanum melongena L.) roots in response to elicitors. Afr. J. Biotechnol. 2010, 9, 8038–8047. [Google Scholar]
- Bautista-Baños, S.; Hernández-López, M.; Bosquez-Molina, E.; Wilson, C. Effects of chitosan and plant extracts on growth of Colletotrichum gloeosporioides, anthracnose levels and quality of papaya fruit. Crop Prot. 2003, 22, 1087–1092. [Google Scholar] [CrossRef]
- Sivakumar, D.; Sultanbawa, Y.; Ranasingh, N.; Wijesundera, R.L.C. Effect of the combined application of chitosan and carbonate salts on the incidence of anthracnose and on the quality of papaya during storage. J. Hortic. Sci. Biotechnol. 2005, 80, 447–452. [Google Scholar] [CrossRef]
- Cheah, L.; Page, B.; Shepherd, R. Chitosan coating for inhibition of sclerotinia rot of carrots. N. Z. J. Crop Hortic. Sci. 1997, 25, 89–92. [Google Scholar] [CrossRef]
- Aziz, A.; Trotel-Aziz, P.; Dhuicq, L.; Jeandet, P.; Couderchet, M.; Vernet, G. Chitosan oligomers and copper sulphate induce grapevine defense reaction and resistance to grey mould and down mildew. Phytopathology 2006, 96, 1188–1194. [Google Scholar] [CrossRef] [Green Version]
- Bell, A.A.; Hubbard, J.C.; Liu, L.; Davis, R.M.; Subbarao, K.V. Effects of chitin and chitosan on the incidence and severity of Fusarium yellows in celery. Plant Dis. 1998, 82, 322–328. [Google Scholar] [CrossRef] [Green Version]
- El Ghaouth, A.; Arul, J.; Grenier, J.; Asselin, A. Antifungal activity of chitosan on two postharvest pathogens of strawberry fruits. Phytopathology 1992, 82, 398–402. [Google Scholar] [CrossRef]
- Lowe, A.; Rafferty-McArdle, S.M.; Cassells, A.C. Effects of AMF- and PGPR-root inoculation and a foliar chitosan spray in single and combined treatments on powdery mildew disease in strawberry. Agric. Food Sci. 2012, 21, 28–38. [Google Scholar] [CrossRef]
- Du, J.; Gemma, H.; Iwahori, S. Effects of chitosan coating on the storage of peach Japanese pear and kiwifruit. J. Jpn. Soc. Hort. Sci. 1997, 66, 15–22. [Google Scholar] [CrossRef]
- Jinasena, D.; Pathirathna, P.; Wickramarachchi, S.; Marasinghe, E. Use of chitosan to control anthracnose on “Embul” banana. In Proceedings of the 2011 International Conference on Asia Agriculture and Animal IPCBEE, Hong Kong, China, 2 July 2011; pp. 56–60. [Google Scholar]
- Gutiérrez-Martínez, P.; Ramos-Guerrero, A.; Rodríguez-Pereida, C.; Coronado-Partida, L.; Angulo-Parra, J.; González-Estrada, R. Chitosan for postharvest disinfection of fruits and vegetables. Postharv. Disinfect. Fruits Veget. 2018, 1, 231–241. [Google Scholar]
- Jitareerat, P.; Paumchai, S.; Kanlayanarat, S.; Sangchote, S. Effect of chitosan on ripening, enzymatic activity, and disease development in mango (Mangifera indica) fruit. N. Z. J. Crop Hortic. Sci. 2007, 35, 211–218. [Google Scholar] [CrossRef]
- Jongsri, P.; Wangsomboondee, T.; Rojsitthisak, P.; Seraypheap, K. Effect of molecular weights of chitosan coating on postharvest quality and physicochemical characteristics of mango fruit. LWT Food Sci. Technol. 2016, 73, 28–36. [Google Scholar] [CrossRef]
- De Oliveira, K.Á.R.; Berger, L.R.R.; de Araújo, S.A.; Câmara, M.P.S.; de Souza, E.L. Synergistic mixtures of chitosan and Mentha piperita L. essential oil to inhibit Colletotrichum species and anthracnose development in mango cultivar Tommy Atkins. Food Microbiol. 2017, 66, 96–103. [Google Scholar] [CrossRef] [PubMed]
- Zeng, K.; Deng, Y.; Ming, J.; Deng, L. Induction of disease resistance and ROS metabolism in navel oranges by chitosan. Sci. Hortic. 2010, 126, 223–228. [Google Scholar] [CrossRef]
- Romanazzi, G.; Mlikota Gabler, F.; Smilanick, J.L. Preharvest chitosan and postharvest UV-C irradiation treatments suppress gray mold of table grapes. Plant Dis. 2006, 90, 445–450. [Google Scholar] [CrossRef] [Green Version]
- Romanazzi, G.; Nigro, F.; Ippolito, A. Short hypobaric treatments potentiate the effect of chitosan in reducing storage decay of sweet cherries. Postharvest Biol. Technol. 2003, 29, 73–80. [Google Scholar] [CrossRef]
- Chandra, S.; Chakraborty, N.; Panda, K.; Acharya, K. Chitosan-induced immunity in Camellia sinensis (L.) O. Kuntze against blister blight disease is mediated by nitric-oxide. Plant Physiol. Biochem. 2017, 115, 298–307. [Google Scholar] [CrossRef]
- Maluin, F.N.; Hussein, M.Z.; Yusof, N.A.; Fakurazi, S.; Idris, A.S.; Hilmi, N.H.Z.; Daim, L.D.J. Chitosan-based agronanofungicides as a sustainable alternative in the basal stem rot disease management. J. Agric. Food Chem. 2020, 68, 4305–4314. [Google Scholar] [CrossRef]
- Kishore, G.K.; Pande, S.; Podile, A.R. Chitin-supplemented foliar application of Serratia marcescens GPS 5 improves control of late leaf spot disease of groundnut by activating defense-related enzymes. J. Phytopathol. 2005, 153, 169–173. [Google Scholar] [CrossRef] [Green Version]
- Li, B.; Liu, B.; Shan, C.; Ibrahim, M.; Lou, Y.; Wang, Y.; Xie, G.; Li, H.Y.; Sun, G.C. Antibacterial activity of two chitosan solutions and their effect on rice bacterial leaf blight and leaf streak. Pest Manag. Sci. 2013, 69, 312–320. [Google Scholar] [CrossRef] [PubMed]
- Ramkissoon, A.; Francis, J.; Bowrin, V.; Ramjegathesh, R.; Ramsubhag, A.; Jayaraman, J. Bio efficacy of a chitosan based elicitor on Alternaria solani and Xanthomonas vesicatoria infections in tomato under tropical conditions. Ann. Appl. Biol. 2016, 169, 274–283. [Google Scholar] [CrossRef]
- Li, B.; Liu, B.; Su, T.; Fang, Y.; Xie, G.; Wang, G.; Wang, Y.; Sun, G. Effect of chitosan solution on the inhibition of Pseudomonas fluorescens causing bacterial head rot of broccoli. Plant Pathol. J. 2010, 26, 189–193. [Google Scholar] [CrossRef] [Green Version]
- Toan, N.V.; Hanh, T.T.; Thien, P.V.M. Antibacterial activity of chitosan on some common food contaminating microbes. Open Biomat. J. 2013, 4, 1–5. [Google Scholar] [CrossRef] [Green Version]
- Beausejour, J.; Clermont, N.; Beaulieu, C. Effect of Streptomyces melanosporofaciens strain EF-76 and of chitosan on common scab of potato. Plant Soil 2003, 256, 463–468. [Google Scholar] [CrossRef]
- Lou, M.M.; Zhu, B.; Muhammad, I.; Li, B.; Xie, G.L.; Wang, Y.L.; Li, H.Y.; Sun, G.C. Antibacterial activity and mechanism of action of chitosan solutions against apricot fruit rot pathogen Burkholderia seminalis. Carbohydr. Res. 2011, 346, 1294–1301. [Google Scholar] [CrossRef]
- Yang, C.; Li, B.; Ge, M.; Zhou, K.; Wang, Y.; Luo, J.; Ibrahim, M.; Xie, G.; Sun, G. Inhibitory effect and mode of action of chitosan solution against rice bacterial brown stripe pathogen Acidovorax avenae subsp. avenae RS-1. Carbohydr. Res. 2014, 391, 48–54. [Google Scholar] [CrossRef]
- Li, B.; Shi, S.; Shan, C.; Zhou, Q.; Ibrahim, M.; Wang, Y.; Wu, G.; Li, H. Effect of chitosan solution on the inhibition of Acidovorax citrulli causing bacterial fruit blotch of watermelon. J. Sci. Food Agric. 2013, 93, 1010–1015. [Google Scholar] [CrossRef]
- Farag, S.M.A.; Elhalag, K.M.A.; Mohamed, H.; Hagag, M.H.; Khairy, A.S.M.; Ibrahim, H.M.; Saker, M.T.; Messiha, N.A.S. Potato bacterial wilt suppression and plant health improvement after application of different antioxidants. J. Phytopathol. 2017, 65, 522–537. [Google Scholar] [CrossRef]
- Acar, O.; Aki, C.; Erdugan, H. Fungal and bacterial diseases control with ElexaTM plant booster. Fresenius Environ. Bull. 2008, 17, 797–802. [Google Scholar]
- Liu, N.; Chen, X.G.; Park, H.J.; Liu, C.G.; Liu, C.S.; Meng, X.H.; Yu, L.J. Effect of MW and concentration of chitosan on antibacterial activity of Escherichia coli. Carbohydr. Polym. 2006, 64, 60–65. [Google Scholar] [CrossRef]
- Shanmugam, A.; Kathiresan, K.; Nayak, L. Preparation, characterization and antibacterial activity of chitosan and phosphorylated chitosan from cuttlebone of Sepia kobiensis (Hoyle, 1885). Biotechnol. Rep. 2016, 9, 25–30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sapers, G.M. Chitosan enhances control of enzymatic browning in apple and pear juice by filtration. J. Food Sci. 1992, 57, 1192–1193. [Google Scholar] [CrossRef]
- Rabea, E.I.; Steurbaut, W. Chemically modified chitosans as antimicrobial agents against some plant pathogenic bacteria and fungi. Plant Protect. Sci. 2010, 46, 149–158. [Google Scholar] [CrossRef]
- Chung, Y.C.; Wang, H.L.; Chen, Y.M.; Li, S.L. Effect of abiotic factors on the antibacterial activity of chitosan against waterborne pathogens. Bioresour. Technol. 2003, 88, 179–184. [Google Scholar] [CrossRef]
- Algam, S.; Xie, G.; Li, B.; Yu, S.; Su, T.; Larsen, J. Effects of Paenibacillus strains and chitosan on plant growth promotion and control of Ralstonia wilt in tomato. J. Plant Pathol. 2010, 92, 593–600. [Google Scholar]
- Coqueiro, D.S.O.; di Piero, M.R. Antibiotic activity against Xanthomonas gardneri and protection of tomato plants by chitosan. J. Plant Pathol. 2011, 93, 337–344. [Google Scholar]
- Ferrante, P.; Scortichini, M. Molecular and phenotypic features of Pseudomonas syringae pv. actinidiae isolated during recent epidemics of bacterial canker on yellow kiwifruit (Actinidia chinensis) in central Italy. Plant Pathol. 2010, 59, 954–962. [Google Scholar] [CrossRef]
- Li, B.; Wang, X.; Chen, R.; Huangfu, W.; Xie, G.L. Antibacterial activity of chitosan solution against Xanthomonas pathogenic bacteria isolated from Euphorbia pulcherrima. Carbohydr. Polym. 2008, 72, 287–292. [Google Scholar] [CrossRef]
- Chirkov, S.N. The antiviral activity of chitosan (review). Appl. Biochem. Microbiol. 2002, 38, 5–13. [Google Scholar] [CrossRef]
- Chirkov, S.N.; Il’ina, A.V.; Surgucheva, N.; Letunova, E.; Varitsev, Y.A.; Tatarinova, N.Y.; Varlamov, V. Effect of chitosan on systemic viral infection and some defense responses in potato plants. Russ. J. Plant Physiol. 2001, 48, 774–779. [Google Scholar] [CrossRef]
- Bondok, A. Response of tomato plants to salicylic acid and chitosan under infection with tomato mosaic virus. Am.-Eur. J. Agric. Environ. Sci. 2015, 15, 1520–1529. [Google Scholar]
- Mishra, S.; Jagadeesh, K.S.; Krishnaraj, P.U.; Prem, S. Biocontrol of tomato leaf curl virus (ToLCV) in tomato with chitosan supplemented formulations of Pseudomonas sp. under field conditions. Aust. J. Crop Sci. 2014, 8, 347–355. [Google Scholar]
- Firmansyah, D. Use of chitosan and plant growth promoting rhizobacteria to control squash mosaic virus on cucumber plants. Asian J. Plant Pathol. 2017, 11, 148–155. [Google Scholar] [CrossRef]
- Pospieszny, H.; Chirkov, S.; Atabekov, J. Induction of antiviral resistance in plants by chitosan. Plant Sci. 1991, 79, 63–68. [Google Scholar] [CrossRef]
- Nagorskaya, V.; Reunov, A.; Lapshina, L.; Davydova, V.; Yermak, I. Effect of chitosan on tobacco mosaic virus (TMV) accumulation, hydrolase activity, and morphological abnormalities of the viral particles in leaves of N. tabacum L. cv. Samsun. Virol. Sin. 2014, 29, 250–256. [Google Scholar] [CrossRef]
- Iriti, M.; Sironi, M.; Gomarasca, S.; Casazza, A.P.; Soave, C.; Faoro, F. Cell death-mediated antiviral effect of chitosan in tobacco. Plant Physiol. Biochem. 2006, 44, 893–900. [Google Scholar] [CrossRef]
- Scorza, R.; Callahan, A.; Levy, L.; Damsteegt, V.; Webb, K.; Ravelonandro, M. Post-transcriptional gene silencing in plum pox virus resistant transgenic European plum containing the plum pox potyvirus coat protein gene. Transgenic Res. 2001, 10, 201–209. [Google Scholar] [CrossRef]
- Kalaiarasan, P.; Lakshmanan, P.; Rajendran, G.; Samiyappan, R. Chitin and chitinolytic biocontrol agents for the management of root knot nematode, Meloidogyne arenaria in groundnut (Arachis hypogaea L.) cv. Co3. Indian J. Nematol. 2006, 36, 181–186. [Google Scholar]
- El-Sayed, S.M.; Mahdy, M.E. Effect of chitosan on root-knot nematode, Meloidogyne javanica on tomato plants. Int. J. ChemTech Res. 2015, 7, 1985–1992. [Google Scholar]
- Westerdahl, B.B.; Carlson, H.L.; Grant, J.; Radewald, J.D.; Welch, N.; Anderson, C.A.; Darso, J.; Kirby, D.; Shibuya, F. Management of plant-parasitic nematodes with a chitin-urea soil amendment and other materials. J. Nematol. 1992, 24, 669–680. [Google Scholar] [PubMed]
- Fan, Z.; Qin, Y.; Liu, S.; Xing, R.; Yu, H.; Li, P. Chitosan oligosaccharide fluorinated derivative control root-knot nematode (Meloidogyne incognita) disease based on the multi-efficacy strategy. Mar. Drugs 2020, 18, 273. [Google Scholar] [CrossRef] [PubMed]
- Liang, W.; Yu, A.; Wang, G.; Zheng, F.; Jia, J.; Xu, H. Chitosan-based nanoparticles of avermectin to control pine wood nematodes. Int. J. Biol. Macromol. 2018, 112, 258–263. [Google Scholar] [CrossRef] [PubMed]
- Ibrahim, A.Y.; Kurniawati, F. The efficacy of chitosan to control nematode Aphelenchoides besseyi Christie through seed treatment. IOP Conf. Ser. Earth Environ. Sci. 2020, 468, 012025. [Google Scholar] [CrossRef]
- Khalil, M.S.; Badawy, M.E. Nematicidal activity of a biopolymer chitosan at different molecular weights against root-knot nematode, Meloidogyne incognita. Plant Protect. Sci. 2012, 48, 170–178. [Google Scholar] [CrossRef] [Green Version]
- Escudero, N.; Lopez-Moya, F.; Ghahremani, Z.; Zavala-Gonzalez, E.A.; Alaguero-Cordovilla, A.; Ros-Ibañez, C.; Lacasa, A.; Sorribas, F.J.; Lopez-Llorca, L.V. Chitosan increases tomato root colonization by Pochonia chlamydosporia and their combination reduces root-knot nematode damage. Front. Plant Sci. 2017, 8, 1415. [Google Scholar] [CrossRef] [PubMed]
- Belair, G.; Tremblay, N. The influence of chitin-urea amendments applied to an organic soil on a Meloidogyne hapla population and on the growth of greenhouse tomato. Phytoprotection 1995, 76, 75–80. [Google Scholar] [CrossRef] [Green Version]
- Asif, M.; Ahmad, F.; Tariq, M.; Khan, A.; Ansari, T.; Khan, F.; Siddiqui, A.M. Potential of chitosan alone and in combination with agricultural wastes against the root-knot nematode, Meloidogyne incognita infesting eggplant. J. Plant Prot. Res. 2017, 57, 288–295. [Google Scholar] [CrossRef] [Green Version]
- El-Saedy, M.A.M.; Hammad, S.E.; Awd Allah, S.F.A. Nematicidal effect of abamectin, boron, chitosan, hydrogen peroxide and Bacillus thuringiensis against citrus nematode on Valencia orange trees. J. Plant Sci. Phytopathol. 2019, 3, 111–117. [Google Scholar]
- Hameed, A.; Sheikh, M.; Hameed, A.; Farooq, T.; Basra, S.; Jamil, A. Chitosan priming enhances the seed germination, antioxidants, hydrolytic enzymes, soluble proteins and sugars in wheat seeds. Agrochimica 2013, 67, 32–46. [Google Scholar]
- John, M.; Röhrig, H.; Schmidt, J.; Walden, R.; Schell, J. Cell signalling by oligosaccharides. Trends Plant Sci. 1997, 2, 111–115. [Google Scholar] [CrossRef]
- Dzung, N.A.; Khanh, V.T.P.; Dzung, T.T. Research on impact of chitosan oligomers on biophysical characteristics, growth, development and drought resistance of coffee. Carbohydr. Polym. 2011, 84, 751–755. [Google Scholar] [CrossRef]
- Amin, A.A.; Rashad EL-Sh, M.; EL-Abagy, H.M.H. Physiological effect of indole-3-butyric acid and salicylic acid on growth, yield and chemical constituents of onion plants. J. Appl. Sci. Res. 2007, 3, 1554–1563. [Google Scholar]
- Zeng, D.; Luo, X. Physiological effects of chitosan coating on wheat growth and activities of protective enzyme with drought tolerance. J. Soil Sci. 2012, 2, 282–288. [Google Scholar] [CrossRef] [Green Version]
- Dzung, N.A.; Thang, N.T. Effect of oligoglucosamine prepared by enzyme degradation on the growth of soybean. Adv. Chitin Sci. 2002, 5, 463–467. [Google Scholar]
- Xing, K.; Zhu, X.; Peng, X.; Qin, S. Chitosan antimicrobial and eliciting properties for pest control in agriculture: A review. Agron. Sustain. Dev. 2015, 35, 569–588. [Google Scholar] [CrossRef] [Green Version]
- Zhang, D.; Wang, H.; Hu, Y.; Liu, Y. Chitosan controls postharvest decay on cherry tomato fruit possibly via the mitogen-activated protein kinase signaling pathway. J. Agric. Food Chem. 2015, 63, 7399–7404. [Google Scholar] [CrossRef]
- Li, P.; Cao, Z.; Wu, Z.; Wang, X.; Li, X. The effect and action mechanisms of oligochitosan on control of stem dry rot of Zanthoxylum bungeanum. Int. J. Mol. Sci. 2016, 17, 1044. [Google Scholar] [CrossRef]
- Shahidi, F.; Arachchi, J.K.V.; Jeon, Y.J. Food applications of chitin and chitosan-review. Trends Food Sci. Technol. 1999, 10, 37–51. [Google Scholar] [CrossRef]
- Helander, I.M.; Nurmiaho-Lassila, E.L.; Ahvenainen, R.; Rhoades, J.; Roller, S. Chitosan disrupts the barrier properties of the outer membrane of Gram-negative bacteria. Int. J. Food Microbiol. 2001, 71, 235–244. [Google Scholar] [CrossRef]
- Xing, K.; Chen, X.G.; Liu, C.S.; Cha, D.S.; Park, H.J. Oleoyl-chitosan nanoparticles inhibits Escherichia coli and Staphylococcus aureus by damaging the cellmembrane and putative binding to extracellular or intracellular targets. Int. J. Food Microbiol. 2009, 132, 127–133. [Google Scholar] [CrossRef] [PubMed]
- García-Rincóna, J.; Vega-Pérez, J.; Guerra-Sánchez, M.G.; Hernández-Lauzardo, A.N.; Peña-Díaz, A.; Velázquez-Del Valle, M.G. Effect of chitosan on growth and plasma membrane properties of Rhizopus stolonifera (Ehrenb.:Fr.) Vuill. Pestic. Biochem. Phys. 2010, 97, 275–278. [Google Scholar]
- Rabea, E.I.; Badawy, M.E.-T.; Stevens, C.V.; Smagghe, G.; Steurbaut, W. Chitosan as antimicrobial agent: Applications and mode of action. Biomacromolecules 2003, 4, 1457–1465. [Google Scholar] [CrossRef] [PubMed]
- Chung, Y.C.; Su, Y.P.; Chen, C.C.; Jia, G.; Wang, H.L.; Wu, J.C.; Lin, J.G. Relationship between antibacterial activity of chitosan and surface characteristics of cell wall. Acta Pharmacol. Sin. 2004, 25, 932–936. [Google Scholar]
- Liu, H.; Du, Y.M.; Wang, X.H.; Sun, L.P. Chitosan kills bacteria through cell membrane damage. Int. J. Food Microbiol. 2004, 95, 147–155. [Google Scholar] [CrossRef]
- Gow, N.A.R.; Latge, J.-P.; Munro, C.A. The fungal cell wall: Structure, biosynthesis, and function. Microbiol. Spectr. 2017, 5, 267–292. [Google Scholar]
- Chung, Y.C.; Chen, C.Y. Antibacterial characteristics and activity of acid-soluble chitosan. Bioresour. Technol. 2008, 99, 2806–2814. [Google Scholar] [CrossRef]
- Hadwiger, L.A. Multiple effects of chitosan on plant systems: Solid science or hype. Plant Sci. 2013, 208, 42–49. [Google Scholar] [CrossRef]
- Sudarshan, N.R.; Hoover, D.G.; Knorr, D. Antibacterial action of chitosan. Food Biotechnol. 1992, 6, 257–272. [Google Scholar] [CrossRef]
- Goy, R.C.; Britto, D.; Assis, O.B.G. A review of the antimicrobial activity of chitosan. Polímeros 2009, 19, 241–247. [Google Scholar] [CrossRef]
- Come, V.; Deschamps, A.; Mertial, A. Bioactive packaging materials from edible chitosan polymer-antimicrobial activity assessment on dairy-related contaminants. J. Food Sci. 2003, 68, 2788–2792. [Google Scholar] [CrossRef]
- Issam, S.T.; Adele, M.G.; Adele, C.P.; Stephane, G.; Veronique, C. Chitosan polymer as bioactive coating and film against Aspergillus niger contamination. J. Food Sci. 2005, 70, 100–104. [Google Scholar]
- Qin, Y. The chelating properties of chitosan fibers. J. Appl. Polym. Sci. 1993, 49, 727–731. [Google Scholar] [CrossRef]
- Freeman, B.C.; Beattie, G.A. An overview of plant defenses against pathogens and herbivores. Plant Health Instr. 2008, 10, 1094. [Google Scholar] [CrossRef] [Green Version]
- Martins, A.; Facchi, S.; Follmann, H.; Pereira, A.; Rubira, A.; Muniz, E. Antimicrobial activity of chitosan derivatives containing N-quaternized moieties in its backbone: A review. Int. J. Mol. Sci. 2014, 15, 20800–20832. [Google Scholar] [CrossRef]
- El Ghaouth, A.; Ponnampalam, R.; Castaigne, F.; Arul, J. Chitosan coating to extend the storage life of tomatoes. HortScience 1992, 27, 1016–1018. [Google Scholar] [CrossRef] [Green Version]
- Dornenburg, H.; Knorr, D. Evaluation of elicitor- and high-pressure-induced enzymatic browning utilizing potato (Solanum tuberosum) suspension cultures as a model system for plant tissues. J. Agric. Food Chem. 1997, 45, 4173. [Google Scholar] [CrossRef]
- Bhaskara, M.V.; Arul, J.; Essaid, A.B.; Anger, P.; Richard, C.; Castaigne, F. Effect of chitosan on growth and toxin production by Alternaria alternata f. sp. lycopersici. Biocontrol Sci. Technol. 1998, 8, 33. [Google Scholar]
- El Ghaouth, A.; Arul, J.; Asselin, A.; Benhamou, N. Antifungal activity of chitosan on post-harvest pathogens: Induction of morphological and cytological alterations in Rhizopus stolonifera. Mycol. Res. 1992, 96, 769. [Google Scholar] [CrossRef]
- Lin, W.; Hu, X.; Zhang, W.; Rogers, W.J.; Cai, W. Hydrogen peroxide mediates defence responses induced by chitosans of different molecular weights in rice. J. Plant Physiol. 2005, 162, 937–944. [Google Scholar] [CrossRef]
- Burkhanova, G.F.; Yarullina, L.G.; Maksimov, I.V. The control of wheat defense responses during infection with Bipolaris sorokiniana by chitooligosaccharides. Russ. J. Plant Physiol. 2007, 54, 104–110. [Google Scholar] [CrossRef]
- Eilenberg, H.; Pnini-Cohen, S.; Rahamim, Y.; Sionov, E.; Segal, E.; Carmeli, S.; Zilberstein, A. Induced production of antifungal naphthoquinones in the pitchers of the carnivorous plant Nepenthes khasiana. J. Exp. Bot. 2009, 61, 911–922. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yin, H.; Zhao, X.; Du, Y. Oligochitosan: A plant diseases vaccine-a review. Carbohydr. Polym. 2010, 82, 1–8. [Google Scholar] [CrossRef]
- Romanazzi, G. Chitosan treatment for the control of postharvest decay of table grapes, strawberries and sweet cherries. Fresh Prod. 2010, 4, 111–115. [Google Scholar]
- Chang, T.; Kim, B.S. Application of chitosan preparations for eco-friendly control of potato late blight. Res. Plant Dis. 2012, 18, 338–348. [Google Scholar] [CrossRef] [Green Version]
- Rahman, M.; Punja, Z.K. Biochemistry of rusty root on American ginseng (Panax quinquefolius L.). Plant Physiol. Biochem. 2005, 43, 1103–1114. [Google Scholar] [CrossRef] [PubMed]
- Orzali, L.; Forni, C.; Riccioni, L. Effect of chitosan seed treatment as elicitor of resistance to Fusarium graminearum in wheat. Seed Sci. Technol. 2014, 42, 132–149. [Google Scholar] [CrossRef]
- Hoat, T.X.; Nakayashiki, H.; Yang, Q.; Tosa, Y.; Mayama, S. Molecular cloning of the apoptosis-related calcium-binding protein AsALG-2 in Avena sativa. Mol. Plant Pathol. 2013, 14, 222–229. [Google Scholar] [CrossRef]
- Hadwiger, L.A.; Beckman, J.M. Chitosan as a component of pea-Fusarium solani interactions. Plant Physiol. 1980, 66, 205–211. [Google Scholar] [CrossRef] [Green Version]
- Klessig, D.F.; Durner, J.; Shah, J.; Yang, Y. Salicylic Acid-Mediated Signal Transduction in Plant Disease Resistance. In Phytochemical Signals and Plant-Microbe Interactions; Romeo, T.J., Downum, R.K., Verpoorte, R., Eds.; Springer: New York, NY, USA, 1998; pp. 119–137. [Google Scholar]
- Lienart, Y.; Gautier, C.; Domard, A. Isolation from Rubus cell-suspension cultures of a lectin specific for glucosamine oligomers. Planta 1991, 184, 8–13. [Google Scholar] [CrossRef]
- Petutschnig, E.K.; Jones, A.M.; Serazetdinova, L.; Lipka, U.; Lipka, V. The Lysin Motif Receptor-Like Kinase (LysM-RLK) CERK1 is a major chitin-binding protein in Arabidopsis thaliana and subject to chitin-induced phosphorylation. J. Biol. Chem. 2010, 285, 28902–28911. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rahman, M.; Rahman, M.; Islam, T. Improving yield and antioxidant properties of strawberries by utilizing microbes and natural products. In Strawberry: Pre- and Post-Harvest Management Techniques for Higher Fruit Quality; Asao, T., Asaduzzaman, M., Eds.; IntechOpen: London, UK, 2019. [Google Scholar] [CrossRef] [Green Version]
- Galván, M.I.; Akuaku, J.; Cruz, I.; Cheetham, J.; Golshani, A.; Smith, M.L. Disruption of protein synthesis as antifungal mode of action by chitosan. Int. J. Food Microbiol. 2013, 164, 108–112. [Google Scholar] [CrossRef] [PubMed]
- Geisberger, G.; Gyenge, E.B.; Hinger, D.; Käch, A.; Maake, C.; Patzke, G.R. Chitosan-thioglycolic acid as a versatile antimicrobial agent. Biomacromolecules 2013, 14, 1010–1017. [Google Scholar] [CrossRef] [PubMed]
- Palma-Guerrero, J.; Lopez-Jimenez, J.A.; Pérez-Berná, A.J.; Huang, I.C.; Jansson, H.B.; Salinas, J.; Villalaín, J.; Read, N.D.; Lopez-Llorca, L.V. Membrane fluidity determines sensitivity of filamentous fungi to chitosan. Mol. Microbiol. 2010, 75, 1021–1032. [Google Scholar] [CrossRef] [PubMed]
- Palma-Guerrero, J.; Huang, I.C.; Jansson, H.B.; Salinas, J.; Lopez-Llorca, L.V.; Read, N.D. Chitosan permeabilizes the plasma membrane and kills cells of Neurospora crassa in an energy dependent manner. Fungal Genet. Biol. 2009, 46, 585–594. [Google Scholar] [CrossRef] [PubMed]
- Vishu, K.A.B.; Varadaraj, M.C.; Gowda, L.R.; Tharanathan, R.N. Characterization of chito-oligosaccharides prepared by chitosanolysis with the aid of papain and pronase, and their bactericidal action against Bacillus cereus and Escherichia coli. Biochem. J. 2005, 391, 167–175. [Google Scholar]
- Liang, C.; Yuan, F.; Liu, F.; Wang, Y.; Gao, Y. Structure and antimicrobial mechanism of ε-polylysine-chitosan conjugates through Maillard reaction. Int. J. Biol. Macromol. 2014, 70, 427–434. [Google Scholar] [CrossRef]
- Kong, M.; Chen, X.G.; Liu, C.S.; Liu, C.G.; Meng, X.H.; Yu, L.J. Antibacterial mechanism of chitosan microspheres in a solid dispersing system against E. coli. Colloid Surf. B 2008, 65, 197–202. [Google Scholar] [CrossRef]
- Tang, H.; Zhang, P.; Kieft, T.L.; Ryan, S.J.; Baker, S.M.; Wiesmann, W.P.; Rogelj, S. Antibacterial action of a novel functionalized chitosanarginine against gram-negative bacteria. Acta Biomater. 2010, 6, 2562–2571. [Google Scholar] [CrossRef] [Green Version]
- Mansilla, A.Y.; Albertengo, L.; Rodríguez, M.S.; Debbaudt, A.; Zúñiga, A.; Casalongué, C.A. Evidence on antimicrobial properties and mode of action of a chitosan obtained from crustacean exoskeletons on Pseudomonas syringae pv. tomato DC3000. Appl. Microbiol. Biotechnol. 2013, 97, 6957–6966. [Google Scholar] [CrossRef]
- Raafat, D.; Bargen, K.V.; Haas, A.; Sahl, H.-G. Insights into the mode of action of chitosan as an antibacterial compound. Appl. Environ. Microbiol. 2008, 74, 3764–3773. [Google Scholar] [CrossRef] [PubMed] [Green Version]



{kind=link}
{kind=link}
| Plant Species | CHT Effects | Mode of Application | References |
|---|---|---|---|
| Rice (Oryza sativa L.) | Increased plant growth, higher photosynthesis rate | In vivo | [55] |
| Soybean (Glycine max) | Increased plant growth | Soil application | [56] |
| Rape (Brassica rapa L.) | Increased plant growth and content of leaf chlorophyll | Hydroponic pot application | [57] |
| Maize (Zea mays L.) | Increased plant growth and grain weight | Biofertilization | [52] |
| Improved seed germination | In vivo | [58] | |
| Improved seed germination and vigor index | In vivo | [59] | |
| Potato (Solanum tuberosum L.) | Increased of tuber size | In vivo | [60] |
| Increased plant growth and yield | In vitro and in vivo | [61] | |
| Tomato (Solanum lycopersicum) | Improved fruit quality and productivity | In vivo | [9,10] |
| Increased seed germination and vigor index | In vivo | [62] | |
| Daikon radishes (Raphanus sativus) | Increased plant growth | In vivo | [47] |
| Cabbage (Brassica oleracea) | Increased plant growth | In vivo | [48] |
| Soybean sprouts (Glycine max) | Increased plant growth | In vivo | [49] |
| Okra (Hibiscus esculentus L.) | Increased plant growth, and yield | In vivo | [63] |
| Eggplant (Solanum melongena) | Increased plant growth, and yield | In vivo | [64] |
| Bean (Phaseolus vulgaris) | Increased leaf area, and carotenoids and chlorophylls levels | In vitro | [65] |
| Chili (Capsicum frutescence L.) | Increased plant growth, yield, and thousand seed weight | In vivo | [66] |
| Increased leaf area, canopy diameter, and plant height | In vivo | [67] | |
| Bell pepper (Capsicum annuum) | Increased fruit weight, diameter, and yield | In vivo | [68] |
| Turmeric (Curcuma longa) | Increased plant growth, and yield | In vivo | [69] |
| Ajowan (Carum copticum) | Increased seed germination, vigor index, dry weight and radical length | In vivo | [70] |
| Artichoke (Cynara scolymus) | Improved seed germination and plant growth | In vivo | [71] |
| Cucumber (Cucumis sativus) | Increased plant growth and improved quality | In vivo | [72] |
| Chickpea (Cicer arietinum) | Increased plant growth | In vivo | [73] |
| Increased seed germination and vigor index | In vivo | [74] | |
| Coffee (Coffea arabica) | Increased plant height and leaf area | In vivo | [75] |
| Strawberry (Fragaria × annanasa) | Increased fruit yield and total antioxidant activities | In vivo | [14] |
| Increased fruit yield | In vivo | [76] | |
| Watermelon (Citrullus lanatus) | Increased plant growth | In vivo | [77] |
| Mango (Mangifera indica) | Increased plant growth, fruit size and weight | In vivo | [78] |
| Grapevine (Vitis vinifera L.) | Increased plant growth | In vivo | [45] |
| Basil (Ocimum ciliatum and Ocimum basilicum) | Increased plant growth and phenol content | In vivo | [50] |
| Phyla dulcis | Increased plant growth | In vitro | [79] |
| Freesia (Freesia corymbosa) | Increased plant growth | In vivo | [53] |
| Gerbera jamesonii | Increased plant growth | In vivo | [51] |
| Dendrobium aggregatum | Increased plant growth | In vitro | [42] |
| Cymbidium insigne | Increased plant growth | In vitro | [44] |
| Kemiri sunan (Reutealis trisperma) | Increased plant growth | In vivo | [80] |
| Scots pine (Pinus sylvestris L.) | Increased plant growth | In vivo | [81] |
| Plant Species | Fungi | Mode of Application | References |
|---|---|---|---|
| Rice (Oryza sativa) | Magnaporthe oryzae | In vitro | [114] |
| M. oryzae | In vivo | [115] | |
| Rhizoctonia solani | In vitro and In vivo | [106] | |
| Jute (Corchorus olitorius) | Macrophomina phaseolina | In vivo | [116] |
| Maize (Zea mays) | Aspergillus flavus | Pre-harvest treatment | [101] |
| Wheat (Triticum aestivum) | Bipolaris sorokiniana | In vivo | [87] |
| Fusarium graminearum | In vivo | [117] | |
| F. graminearum | In vivo | [118] | |
| Pearl millet (Pennisetum glaucum) | Sclerospora graminicola | Post-harvest treatment | [105] |
| Soybean (Glycine max L.) | Heterodera glycines | In vivo | [119] |
| Cowpea (Vigna unguiculata) | F. oxysporum f. sp. tracheiphilum | In vivo | [91] |
| Tobacco (Nicotiana tabacum) | Phytophthora parasitica | In vitro and In vivo | [96] |
| Cherry tomato (Solanum lycopersicum var. cerasiforme) | Botrytis cinerea | Post-harvest treatment | [98] |
| Tomato (S. lycopersicum) | F. oxysporum | In vivo | [108] |
| F. oxysporum f. sp. radicis-lycopersici | In vivo | [120] | |
| F. oxysporum f. sp. radicislycopersici | In vivo | [121] | |
| F. oxysporum f. sp. lycopersici | In vivo | [9] | |
| Alternaria solani | In vivo | [10] | |
| P. infestans | In vivo | [97] | |
| F. ox f. sp. lycopersici | In vitro | [90] | |
| F. solani | |||
| F. oxf. radicis. lycopersici | |||
| A. solani | |||
| P. infestance | |||
| R. solani | |||
| Sclerotium rolfsii | |||
| Potato (S. tuberosum) | F. sembaticum | In vitro | [90] |
| P. infestans | |||
| A. solani, | |||
| R. solani | |||
| P. infestans | In vivo | [122] | |
| P. infestans | In vivo | [107] | |
| Green bean (Phaseolus vulgaris L.) | F. oxysporium | In vitro | [90] |
| F. solani | |||
| R. solani | |||
| S. rolfsii | |||
| Sclerotina sclerotiorum | |||
| B. cienera | |||
| Macrophomina phaseolina | |||
| F. solani | In vivo | [123] | |
| R. solani | |||
| Cucumber (Cucumis sativus L.) | Colletotrichum spp. | Foliar spraying | [113] |
| B. cinerea | Foliar spray | [124] | |
| Sphaerotheca fuliginea | In vitro | [125] | |
| Phytophthora capsici | In vivo | [126] | |
| Pepper (Piper nigrum) | P. capsici | In vivo | [89] |
| Chili pepper (Capsicum annuum) | Colletotrichum capsici | In vivo | [127] |
| P. capsici | In vivo | [128] | |
| Chilli (Capsicum frutescence L.) | C. capsici | In vivo | [66] |
| Eggplant (Solanum melongena) | Ralstonia solanacearum | In vitro | [129] |
| Papaya (Carica papaya) | C. gloeosporioides | In situ | [130] |
| C. gloeosporioides | Post-harvest treatment | [131] | |
| Carrot (Daucus carota) | S. sclerotiorum | In vitro | [132] |
| Grapevine (Vitis vinifera) | Elsinoe ampelina | Post-harvest treatment | [104] |
| Plasmopara viticola | |||
| B. cinerea | In vitro and In vivo | [109] | |
| B. cinerea | In vivo | [133] | |
| P. viticola | |||
| Celery (Apium graveolens) | Fusarium oxysporum f. sp. apii | In vivo | [134] |
| Strawberry (Fragaria × ananassa) | Rhizopus stolonifer | Post-harvest treatment | [135] |
| B. cinerea | |||
| B. cinerea | Pre-harvest treatment | [110] | |
| Sphaerotheca macularis | In vivo | [136] | |
| Dragon fruit (Hylocereus undatus) | C. gleosporoides | In vivo | [111] |
| Kiwifruit (Actinidia deliciosa) | B. cinerea | Post-harvest treatment | [137] |
| Pear (Pyrus communis) | A. kikuchiana | In vitro and Post-harvest treatment | [102] |
| P. piricola | |||
| B. cinerea | Post-harvest treatment | [137] | |
| Peach (Prunus persica) | Monilinia fructicola | Post-harvest treatment | [103] |
| B. cinerea | Post-harvest treatment | [137] | |
| Banana (Musa acuminate) | P. viticola | In vivo | [138] |
| Colletotrichum sp. | Post-harvest treatment | [139] | |
| Fusarium sp. | |||
| Mango (Mangifera indica) | C. gloeosporioides | Post-harvest coating | [140] |
| C. gloeosporioides | Post-harvest coating | [141] | |
| C. asianum | Post-harvest coating | [142] | |
| C. dianesei | |||
| C. fructicola | |||
| C. tropicale | |||
| C. karstii | |||
| Orange (Citrus sinensis) | Penicillium italicum | Post-harvest coating | [143] |
| P. digitatum | |||
| Grape (Vitis vinifera) | Rhizopus stolonifer | Post-harvest treatment | [93] |
| Aspergillus niger | |||
| B. cinerea | Pre-harvest treatment | [144] | |
| Pomegranate (Punica granatum L.) | Botrytis spp. | Post-harvest coating | [95] |
| Penicillium spp. | |||
| Pilidiella granati | |||
| Avocado (Persea americana mill.) | C. gloeosporioides | Post-harvest treatment | [16] |
| Soursop (Annona muricata L.) | C. gloeosporioides | Post-harvest treatment | [139] |
| Jackfruit (Artocarpus heterophyllus L.) | Rhizopus sp. | Post-harvest treatment | [139] |
| Sweet cherry (Prunus avium) | Monilinia fructicola | Post-harvest treatment | [145] |
| B. cinerea | |||
| Tea (Camellia sinensis L.) | Exobasidium vexans | Foliar spraying | [146] |
| Scots Pine (Pinus sylvestris L.) | Fusarium spp. | In vivo | [81] |
| Date palm (Phoenix dactylifera) | F. oxysporum | In vivo | [92] |
| Oil palm (Elaeis guineensis) | Ganoderma boninense | In vivo | [147] |
| Peanut (Arachis hypogaea) | Puccinia arachidis | In vivo | [112] |
| Groundnut (Arachis hypogaea) | Phaeoisariopsis personata | Foliar spraying | [148] |
| Plant Species | Bacteria | Mode of Application | References |
|---|---|---|---|
| Rice (Oryza sativa) | Acidovorax avenae subsp. avenae | In vitro | [155] |
| Xanthomonas oryzae pv. oryzae | In vitro and In vivo | [149] | |
| X. oryzae pv. oryzicola | |||
| Tomato (Solanum lycopersicum) | Ralstoniasolanacearum | In vivo | [164] |
| X. vesicatoria | In vivo | [165] | |
| X. vesicatoria | In vitro and in vivo | [150] | |
| Potato (S. tuberosum) | Streptomyces scabies | In vivo | [153] |
| Ralstonia solanacearum | In vitro and in vivo | [157] | |
| Broccoli (Brassica oleracea var. italic) | Pseudomonas fluorescens | In vitro and In vivo | [151] |
| Cucumber (Cucumis sativus) | P. syringae pv. lachrymans | In vivo | [158] |
| Kiwifruit (Actinidia chinensis) | P. syringae pv. actinidiae | In vitro | [166] |
| Apricot (Prunus armeniaca) | Burkholderia seminalis | In vitro | [154] |
| Watermelon (Citrullus lanatus) | A. citrulli | In vitro | [156] |
| Banana (Musa acuminate) | P. aeruginosa | In vitro | [152] |
| Staphylococcus aureus | |||
| Apple (Malus domestica) | P. aeruginosa | ||
| S. aureus | |||
| Poinsettia (Euphorbia pulcherrima) | Xanthomonas spp. | In vitro | [167] |
| Plant Species | Viruses | References |
|---|---|---|
| Tomato (Lycopersicum esculentum) | PVX, TYLCV | [173] |
| ToLCV | [171] | |
| PSTV, TMV | [168] | |
| Potato (Solanum tuberosum) | PVX | [169] |
| PVY | [168] | |
| Bean (Phuseolus vulgaris) | AMV | [173] |
| AMV, BGMV, PSV, TNV, BYMV, TMV, BCMV | [168] | |
| BCMV | [11] | |
| Cucumber (Cucumis sativus) | SMV | [172] |
| Globe amaranth (Gomphrena globosa L.) | PVX | [168] |
| Pea (Pisum sativum) | AMV, PSV | [173] |
| Quinoa (Chenopodium quinoa) | TNV | [173] |
| CMV, TNV, AMV | [168] | |
| Tobacco (Nicotiana tabacum) | TMV | [174] |
| TNV | [175] | |
| TMV | [170] | |
| PSV | [173] | |
| Stramony (Datura stramonium L.) | FMV, TMV | [168] |
| Plum (Prunus domestica) | PPV | [176] |
| Plant Species | Nematodes | Mode of Application | References |
|---|---|---|---|
| Rice (Oryza sativa) | Aphelenchoides besseyi | In vivo | [182] |
| Tomato (Solanum lycopersicum) | Meloidogyne incognita | In vivo | [180] |
| M. incognita | In vitro and in vivo | [178] | |
| M. incognita | In vitro and Fertigation | [183] | |
| M. incognita | Fertigation | [179] | |
| Meloidogyne spp. | In vivo | [184] | |
| M. javanica | Fertigation | [178] | |
| M. hapla | Fertigation | [185] | |
| Potato (S. tuberosum) | M. chitwood | Fertigation | [179] |
| Eggplant (S. melongena) | M. incognita | Fertigation | [186] |
| Brussels sprouts (Brassica oleracea) | Heterodera schachtii | Fertigation | [179] |
| Valencia orange (Citrus sinensis Valencia) | Tylenchulus semipenetrans | Fertigation | [187] |
| Walnut (Juglans regia) | Pratylenchus vulnus | Fertigation | [179] |
| Groundnut (Arachis hypogaea L.) | M. arenaria | Fertigation | [177] |
| Pinewood (Pinus sp.) | Bursaphelen chusxylophilus | Fertigation | [12] |
| B. chusxylophilus | Fertigation | [181] |
| Pathogens | Mechanisms | References |
|---|---|---|
| Magnaporthe oryzae | Permeabilized the cell plasma membrane and affected the NADPH oxidase-dependent synthesis of ROS | [114] |
| Rhizoctonia solani | Disrupted cell plasma membrane, destroyed cell structures and induced defense-associated enzymes activity in plants | [106] |
| Saccharomyces cerevisiae | Destroyed synthesis of protein, and integrity of cell membrane | [232] |
| Candida albicans | Disrupted the integrity of cell wall and intra-cellular ultrastructure | [233] |
| Beauveria bassiana | Fluidity of the cell membrane determines the vulnerability of fungi to CHT | [234] |
| Pochonia chlamydosporia | ||
| Fusarium oxysporum f. sp. radicis-lycopersici | ||
| F. oxysporum | Had an attraction for lipids in the plasma membrane | [7] |
| F. solani | ||
| Neurospora crassa | Fluidity of the cell membrane determines the vulnerability of fungi to CHT | [234] |
| Permeabilized the cell membrane and destroyed cells | [235] | |
| Rhizopus stolonifer | Triggered K+ efflux and inhibited the activity of H+-ATPase | [200] |
| Aspergillus fumigatus | Had an attraction for lipids in the plasma membrane | [7] |
| Botrytis cinerea | ||
| Aspergillus parasiticus | ||
| Penicillium verrusosum var. verrucosum | ||
| Alternaria alternata | Chelation of metals | [216] |
| Bacillus cereus | Blockage of nutrient flow | [236] |
| Escherichia coli | Disrupted cell membranes and leaked cellular cytoplasm | [237] |
| Destroyed plasma membrane structure of cells, and bind to intracellular or extracellular targets | [199] | |
| Destroyed structure of cells, electrostatic interactions, induced enzyme and nucleotide leakages | [205] | |
| Disrupted cell membrane structures, and leaked cellular cytoplasm | [238] | |
| Destroyed cell membrane | [198,203] | |
| Blockage of nutrient flow | [236] | |
| Increased cell plasma membrane permeability by CHT-membrane interaction | [239] | |
| Neisseria subflava | Destroyed the integrity of cell wall, and intra-cellular ultrastructure | [233] |
| Pseudomonas syringae | Electrostatic interactions, disrupted bacterial cell surface and induced morphological alterations | [240] |
| P. fluorescens | Increased cell plasma membrane permeability by CHT-membrane interaction | [239] |
| P. aeruginosa | Disrupted outer cell membrane | [198] |
| Streptococcus sobrinus | Destroyed the integrity of cell wall, and intra-cellular ultrastructure | [233] |
| Staphylococcus simulans | Electrostatic interactions | [241] |
| S. aureus | Electrostatic interactions | [241] |
| Destroyed cell plasma membrane | [203] | |
| Disrupted cell plasma membranes and leaked cellular cytoplasm | [237] | |
| Damaged structures of cell membrane, and bind to intracellular or extracellular targets | [199] | |
| Salmonella typhimurium | Destroyed the outer cell membrane | [198] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chakraborty, M.; Hasanuzzaman, M.; Rahman, M.; Khan, M.A.R.; Bhowmik, P.; Mahmud, N.U.; Tanveer, M.; Islam, T. Mechanism of Plant Growth Promotion and Disease Suppression by Chitosan Biopolymer. Agriculture 2020, 10, 624. https://doi.org/10.3390/agriculture10120624
Chakraborty M, Hasanuzzaman M, Rahman M, Khan MAR, Bhowmik P, Mahmud NU, Tanveer M, Islam T. Mechanism of Plant Growth Promotion and Disease Suppression by Chitosan Biopolymer. Agriculture. 2020; 10(12):624. https://doi.org/10.3390/agriculture10120624
Chicago/Turabian StyleChakraborty, Moutoshi, Mirza Hasanuzzaman, Mahfuzur Rahman, Md. Arifur Rahman Khan, Pankaj Bhowmik, Nur Uddin Mahmud, Mohsin Tanveer, and Tofazzal Islam. 2020. "Mechanism of Plant Growth Promotion and Disease Suppression by Chitosan Biopolymer" Agriculture 10, no. 12: 624. https://doi.org/10.3390/agriculture10120624
APA StyleChakraborty, M., Hasanuzzaman, M., Rahman, M., Khan, M. A. R., Bhowmik, P., Mahmud, N. U., Tanveer, M., & Islam, T. (2020). Mechanism of Plant Growth Promotion and Disease Suppression by Chitosan Biopolymer. Agriculture, 10(12), 624. https://doi.org/10.3390/agriculture10120624