Root Growth of Hordeum vulgare and Vicia faba in the Biopore Sheath



Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Setup
2.2. Statistical Data Analysis
3. Results
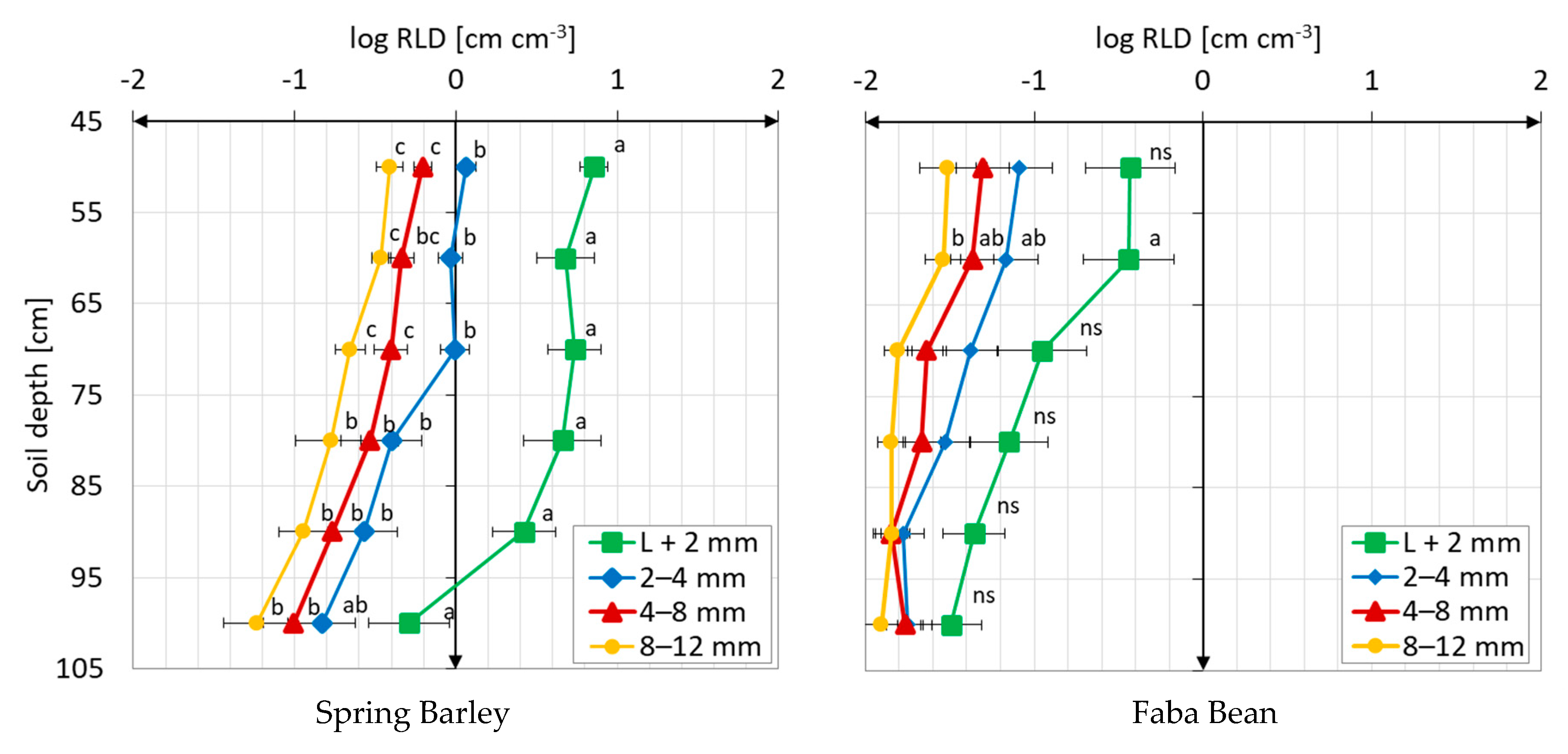
3.1. Root Distribution
3.2. Nt-, Ct-Content and CN-Ratio
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Hodge, A.; Berta, G.; Doussan, C.; Merchan, F.; Crespi, M. Plant root growth, architecture and function. Plant Soil 2009, 321, 153–187. [Google Scholar] [CrossRef]
- Kutschera, L.; Lichtenegger, E.; Sobotik, M. Hordeum vulgare L., vielzeilige gerste. In Wurzelatlas der Kulturpflanzen Gemäßigter Gebiete Mit Arten des Feldgemüseanbaus; DLG-Verlags-GmbH: Frankfurt am Main, Germany, 2009; Volume 7, p. 212. [Google Scholar]
- Amelung, W.; Blume, H.-P.; Fleige, H.; Horn, R.; Kandeler, E.; Kögel-Knabner, I.; Kretzschmar, R.; Stahr, K.; Wilke, B.-M. Böden als pflanzenstandorte. In Scheffer/Schachtschabel Lehrbuch der Bodenkunde, 17th ed.; Springer-Verlag GmbH: Berlin/Heidelberg, Germany, 2018; pp. 491–581. [Google Scholar] [CrossRef]
- Gaiser, T.; Perkons, U.; Küpper, P.M.; Uteau Puschmann, D.; Peth, S.; Kautz, T.; Pfeifer, J.; Ewert, F.; Horn, R.; Köpke, U. Evidence of improved water uptake from subsoil by spring wheat following lucerne in a temperate humid climate. Field Crops Res. 2012, 126, 56–62. [Google Scholar] [CrossRef]
- Fleige, H.; Grimme, H.; Renger, M.; Strebel, O. Zur erfassung der nährstoffanlieferung durch diffusion im effektiven wurzelraum. Mitt. Dtsch. Bodenkd. Ges. 1983, 38, 381–386. [Google Scholar]
- Kuhlmann, H.; Baumgärtel, G. Potential importance of the subsoil for the P and Mg nutrition of wheat. Plant Soil 1991, 137, 259–266. [Google Scholar] [CrossRef]
- Ehlers, W.; Köpke, U.; Hesse, F.; Böhm, W. Penetration resistance and root growth of oats in tilled and untilled loess soil. Soil Tillage Res. 1983, 3, 261–275. [Google Scholar] [CrossRef]
- Kautz, T.; Amelung, W.; Ewert, F.; Gaiser, T.; Horn, R.; Jahn, R.; Javaux, M.; Kemna, A.; Kuzyakov, Y.; Munch, J.C.; et al. Nutrient acquisition from arable subsoils in temperate climates: A review. Soil Biol. Biochem. 2013, 57, 1003–1022. [Google Scholar] [CrossRef]
- Kuzyakov, Y.; Blagodatskaya, E. Microbial hotspots and hot moments in soil: Concept & review. Soil Biol. Biochem. 2015, 83, 184–199. [Google Scholar] [CrossRef]
- Tiunov, A.V.; Scheu, S. Microbial respiration, biomass, biovolume and nutrient status in burrow walls of Lumbricus terrestris L. (Lumbricidae). Soil Biol. Biochem. 1999, 31, 2039–2048. [Google Scholar] [CrossRef]
- Uksa, M.; Schloter, M.; Kautz, T.; Athmann, M.; Köpke, U.; Fischer, D. Spatial variability of hydrolytic and oxidative potential enzyme activities in different subsoil compartments. Biol. Fertil. Soils 2015, 51, 517–521. [Google Scholar] [CrossRef]
- Hagedorn, F.; Bundt, M. The age of preferential flow paths. Geoderma 2002, 108, 119–132. [Google Scholar] [CrossRef]
- Perkons, U.; Kautz, T.; Uteau, D.; Peth, S.; Geier, V.; Thomas, K.; Lütke Holz, K.; Athmann, M.; Pude, R.; Köpke, U. Root-length densities of various annual crops following crops with contrasting root systems. Soil Tillage Res. 2014, 137, 50–57. [Google Scholar] [CrossRef]
- Kautz, T.; Perkons, U.; Athmann, M.; Pude, R.; Köpke, U. Barley roots are not constrained to large-sized biopores in the subsoil of a deep Haplic Luvisol. Biol. Fertil. Soils 2013, 49, 959–963. [Google Scholar] [CrossRef]
- Stewart, J.B.; Moran, C.J.; Wood, J.T. Macropore sheath: Quantification of plant root and soil macropore association. Plant Soil 1999, 211, 59–67. [Google Scholar] [CrossRef]
- Pierret, A.; Moran, C.J.; Pankhurst, C.E. Differentiation of soil properties related to the spatial association of wheat roots and soil macropores. Plant Soil 1999, 211, 51–58. [Google Scholar] [CrossRef]
- White, R.G.; Kirkegaard, J.A. The distribution and abundance of wheat roots in a dense, structured subsoil – implications for water uptake. Plant Cell Environ. 2010, 33, 133–148. [Google Scholar] [CrossRef]
- Stirzaker, R.J.; Passioura, J.B.; Wilms, Y. Soil structure and plant growth: Impact of bulk density and biopores. Plant Soil 1996, 185, 151–162. [Google Scholar] [CrossRef]
- Passioura, J.B. Soil structure and plant growth. Aust. J. Soil Res. 1991, 29, 717–728. [Google Scholar] [CrossRef]
- Athmann, M.; Kautz, T.; Pude, R.; Köpke, U. Root growth in biopores—Evaluation with in situ endoscopy. Plant Soil 2013, 371, 179–190. [Google Scholar] [CrossRef]
- Bouché, M.B. Action de la faune sur les états de la matière organique dans les ecosystèmes. In Humification et Biodégradation; Kilbertius, G., Reisinger, O., Mourey, A., Cancela da Fonseca, J.A., Eds.; Pierron: Sarreguemines, France, 1975; pp. 157–168. [Google Scholar]
- Brown, G.G.; Baroisa, I.; Lavelle, P. Regulation of soil organic matter dynamics and microbial activity in the drilosphere and the role of interactions with other edaphic functional domains. Eur. J. Soil Biol. 2000, 36, 177–198. [Google Scholar] [CrossRef]
- Lavelle, P. Earthworm activities and the soil system. Biol. Fertil. Soils 1988, 6, 237–251. [Google Scholar] [CrossRef]
- Pankhurst, C.E.; Pierret, A.; Hawke, B.G.; Kirby, J.M. Microbiological and chemical properties of soil associated with macropores at different depths in a red-duplex soil in NSW Australia. Plant Soil 2002, 238, 11–20. [Google Scholar] [CrossRef]
- Athmann, M.; Kautz, T.; Banfield, C.; Bauke, S.; Hoang, D.T.T.; Lüsebrink, M.; Pausch, J.; Amelung, W.; Kuzyakov, Y.; Köpke, U. Six months of L. terrestris L. activity in root-formed biopores increases nutrient availability, microbial biomass and enzyme activity. Appl. Soil Ecol. 2017, 120, 135–142. [Google Scholar] [CrossRef]
- Don, A.; Steinberg, B.; Schöning, I.; Pritsch, K.; Joschko, M.; Gleixner, G.; Schulze, E.-D. Organic carbon sequestration in earthworm burrows. Soil Biol. Biochem. 2008, 40, 1803–1812. [Google Scholar] [CrossRef]
- Tiunov, A.V.; Bonkowski, M.; Alphei, J.; Scheu, S. Microflora, Protozoa and Nematoda in Lumbricus terrestris burrow walls: A laboratory experiment. Pedobiologia 2001, 45, 46–60. [Google Scholar] [CrossRef]
- Jégou, D.; Cluzeaua, D.; Hallaireb, V.; Balesdentc, J.; Tréhen, P. Burrowing activity of the earthworms Lumbricus terrestris and Aporrectodea giardi and consequences on C transfers in soil. Eur. J. Soil Biol. 2000, 36, 27–34. [Google Scholar] [CrossRef]
- Andriuzzi, W.; Bolger, T.; Schmidt, O. The drilosphere concept: Fine-scale incorporation of surface residue-derived N and C around natural Lumbricus terrestris burrows. Soil Biol. Biochem. 2013, 64, 136–138. [Google Scholar] [CrossRef]
- Bengough, A.G. Root Growth and function in relation to soil structure, composition, and strength. In Root Ecology; de Kroon, H., Visser, E.J.W., Eds.; Springer: Berlin/Heidelberg, Germany, 2003; pp. 151–171. [Google Scholar]
- Uteau, D.; Pagenkemper, S.K.; Peth, S.; Horn, R. Root and time dependent soil structure formation and its influence on gas transport in the subsoil. Soil Tillage Res. 2013, 132, 69–76. [Google Scholar] [CrossRef]
- Lee, K.E. Earthworms: Their Ecology and Relationship with Soil and Land Use; Academic Press: Sydney, Australia, 1985. [Google Scholar]
- Hinsinger, P.; Bengough, A.G.; Vetterlein, D.; Young, I.M. Rhizosphere: Biophysics, biogeochemistry and ecological relevance. Plant Soil 2009, 321, 117–152. [Google Scholar] [CrossRef]
- Helliwell, J.R.; Sturrock, C.J.; Mairhofer, S.; Craigon, J.; Ashton, R.W.; Miller, A.J.; Whalley, W.R.; Mooney, S.J. The emergent rhizosphere: Imaging the development of the porous architecture at the root-soil interface. Sci. Rep. 2017, 7, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Schrader, S.; Rogasik, H.; Onasch, I.; Jégou, D. Assessment of soil structural differentiation around earthworm burrows by means of X-ray computed tomography and scanning electron microscopy. Geoderma 2007, 137, 378–387. [Google Scholar] [CrossRef]
- Pagenkemper, S.K.; Athmann, M.; Uteau, D.; Kautz, T.; Peth, S.; Horn, R. The effect of earthworm activity on soil bioporosity—Investigated with X-ray computed tomography and endoscopy. Soil Tillage Res. 2015, 146, 79–88. [Google Scholar] [CrossRef]
- Bengough, A.G. Root elongation is restricted by axial but not by radial pressures: So what happens in field soil? Plant Soil 2012, 360, 15–18. [Google Scholar] [CrossRef]
- Kolb, E.; Hartmann, C.; Genet, P. Radial force development during root growth measured by photoelasticity. Plant Soil 2012, 360, 19–35. [Google Scholar] [CrossRef]
- Pätzold, S.; Vetterlein, D.; Jahn, R. DFG Research Unit 1320. Crop Sequence and the Nutrient Acquisition from the Subsoil. Description of the Reference Soil Profile. Available online: https://www.cka.uni-bonn.de/standort/bodenprofilbeschreibung-cka (accessed on 12 November 2020).
- Patterson, H.D.; Thompson, R. Recovery of inter-block information when block sizes are unequal. Biometrika 1971, 58, 545–554. [Google Scholar] [CrossRef]
- Littell, R.C.; Milliken, G.A.; Stroup, W.W.; Wolfinger, R.D.; Schabenberger, O. SAS for Mixed Model, 2nd ed.; SAS Institute Inc.: Cary, NC, USA, 2006. [Google Scholar]
- Kenward, M.G.; Roger, J.R. Small sample inference for fixed effects from restricted maximum likelihood. Biometrics 1997, 53, 983–997. [Google Scholar] [CrossRef] [Green Version]
- Kackar, R.N.; Harville, D.A. Approximations for standard errors of estimators of fixed and random effects in mixed linear models. J. Am. Stat. Assoc. 1984, 388, 853–862. [Google Scholar]
- Piepho, H. A SAS macro for generating letter displays of pairwise mean comparisons. Commun. Biometry Crop Sci. 2012, 1, 4–13. [Google Scholar]
- Athmann, M.; Sondermann, J.; Kautz, T.; Köpke, U. Comparing macropore exploration by Faba bean, wheat, barley and oilseed rape roots using in situ endoscopy. J. Soil Sci. Plant Nutr. 2019, 19, 689–700. [Google Scholar] [CrossRef]
- Popova, L.; Van Dusschoten, D.; Nagel, K.A.; Fiorani, F.; Mazzolai, B. Plant root tortuosity: An indicator of root path formation in soil with different composition and density. Ann. Bot. 2016, 118, 685–698. [Google Scholar] [CrossRef] [Green Version]
- Lynch, J.; Marschner, P.; Rengel, Z. Effect of internal and external factors on root growth and development. In Marschner’s Mineral Nutrition of Higher Plants, 3rd ed.; Marschner, P., Ed.; Academic Press: San Diego, CA, USA, 2012; pp. 331–346. [Google Scholar]
- Perkons, U. Bioporengenese Durch Homo- und Allorhize Kulturpflanzen: Einfluss auf das Wurzelwachstum der Nachfrüchte. Ph.D. Thesis, University of Bonn, Bonn, Germany, 2018. [Google Scholar]
- Han, E.; Kautz, T.; Köpke, U. Precrop root system determines root diameter of subsequent crop. Biol. Fertil. Soils 2016, 52, 113–118. [Google Scholar] [CrossRef]
- Atkinson, J.A.; Hawkesford, M.J.; Whalley, W.R.; Zhou, H.; Mooney, S.J. Soil strength influences wheat root interactions with soil macropores. Plant Cell Environ. 2020, 43, 235–245. [Google Scholar] [CrossRef] [PubMed]
- Materechera, S.A.; Dexter, A.R.; Alston, A.M. Penetration of very strong soils by seedling roots of different species. Plant Soil 1991, 135, 31–41. [Google Scholar] [CrossRef]
- Li, H.B.; Ma, Q.H.; Li, H.G.; Zhan, F.S.; Rengel, Z.; Shen, J.B. Root morphological responses to localized nutrient supply differ among crop species with contrasting root traits. Plant Soil 2014, 376, 151–163. [Google Scholar] [CrossRef]
- Zhan, A.; Lynch, J.P. Reduced frequency of lateral root branching improves N capture from low-N soils in maize. J. Exp. Bot. 2015, 66, 2055–2065. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Colombi, T.; Walter, A. Genetic diversity under soil compaction in wheat: Root number as a promising trait for early plant vigor. Front. Plant Sci. 2017, 8, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Correa, J.; Postma, J.A.; Watt, M.; Wojciechowski, T. Soil compaction and the architectural plasticity of root systems. J. Exp. Bot. 2019, 70, 6019–6034. [Google Scholar] [CrossRef]
- Watt, M.; Hugenholtz, P.; White, R.; Vinall, K. Numbers and locations of native bacteria on field-grown wheat roots quantified by fluorescence in situ hybridization (FISH). Environ. Microbiol. 2006, 8, 871–884. [Google Scholar] [CrossRef]
- Watt, M.; Silk, W.K.; Passioura, J.B. Rates of root and organism growth, soil conditions, and temporal and spatial development of the rhizosphere. Ann. Bot. 2006, 97, 839–855. [Google Scholar] [CrossRef] [Green Version]
- Jones, D.L.; Magthab, E.A.; Gleeson, D.B.; Hill, P.W.; Sánchez-Rodríguez, A.R.; Roberts, P.; Ge, T.; Murphy, D.V. Microbial competition for nitrogen and carbon is as intense in the subsoil as in the topsoil. Soil Biol. Biochem. 2018, 117, 72–82. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Soil Type | Crop | Share of Roots [%] | Reference |
|---|---|---|---|
| Haplic Luvisol (silty loam) | Winter barley, Oilseed rape | 21 | Perkons et al. [13] |
| Haplic Luvisol (silty loam) | Winter barley | <25 | Kautz et al. [14] |
| Black Vertosol | Pasture, dominated by Queens-land blue grass and Tall oat grass | 11–26 | Stewart et al. [15] |
| Typic and Haplic Palexeralf (hard setting clay) | Wheat | 80 * | Pierret et al. [16] |
| Red Kondosol (acidic loam) | Wheat | 44–95 ** | White and Kirkegaard [17] |
| Fine | Small | Medium | Coarse | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Lateral | 0–0.2 mm | 0.2–0.4 mm | 0.4–0.6 mm | 0.6–5 mm | |||||
| Distance | (%) | (%) | (%) | (%) | |||||
| (mm) | Mean | SE | Mean | SE | Mean | SE | Mean | SE | |
| Spring Barley | L + 2 | 43.7 b | ±4.1 | 34.9 a | ±2.9 | 13.4 a | ±2.0 | 7.9 a | ±1.7 |
| 2–4 | 53.1 a | ±5.3 | 38.2 a | ±4.2 | 7.0 b | ±2.1 | 1.8 b | ±1.0 | |
| 4–8 | 51.3 a | ±4.9 | 41.8 a | ±4.3 | 5.8 b | ±1.5 | 1.1 b | ±0.6 | |
| 8–12 | 52.5 a | ±4.6 | 39.5 a | ±3.8 | 7.0 b | ±1.6 | 1.0 b | ±0.5 | |
| Faba Bean | L + 2 | 43.9 a | ±11.9 | 31.3 a | ±7.7 | 11.8 a | ±4.6 | 13.0 a | ±5.9 |
| 2–4 | 62.8 a | ±12.3 | 26.6 a | ±8.3 | 6.6 a | ±4.7 | 4.0 ab | ±3.1 | |
| 4–8 | 56.0 a | ±10.9 | 32.7 a | ±9.1 | 6.2 a | ±2.9 | 5.1 ab | ±4.0 | |
| 8–12 | 53.1 a | ±13.5 | 38.6 a | ±11.7 | 6.1 a | ±3.9 | 2.2 b | ±1.5 | |
| Spring Barley | Faba Bean | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Soil Depth | Lateral Distance | Nt (%) | Ct (%) | C/N | Nt (%) | Ct (%) | C/N | ||||||
| (cm) | (mm) | Mean | SE | Mean | SE | Mean | SE | Mean | SE | Mean | SE | Mean | SE |
| 45–55 | 0–2 | 0.070 a | ±0.003 | 0.65 a | ±0.02 | 9.3 a | ±0.2 | 0.062 a | ±0.003 | 0.55 a | ±0.03 | 8.8 a | ±0.2 |
| 2–4 | 0.060 b | ±0.003 | 0.52 b | ±0.02 | 8.8 ab | ±0.2 | 0.051 b | ±0.001 | 0.43 ab | ±0.01 | 8.4 ab | ±0.2 | |
| 4–8 | 0.056 c | ±0.002 | 0.47 b | ±0.01 | 8.5 b | ±0.3 | 0.047 b | ±0.001 | 0.39 b | ±0.01 | 8.2 b | ±0.2 | |
| 8–12 | 0.055 c | ±0.002 | 0.46 b | ±0.01 | 8.5 b | ±0.3 | 0.046 b | ±0.001 | 0.38 b | ±0.01 | 8.1 b | ±0.1 | |
| 55–65 | 0–2 | 0.068 a | ±0.003 | 0.61 a | ±0.02 | 8.9 a | ±0.2 | 0.060 a | ±0.002 | 0.52 a | ±0.03 | 8.6 a | ±0.2 |
| 2–4 | 0.058 b | ±0.002 | 0.50 b | ±0.02 | 8.5 ab | ±0.3 | 0.052 b | ±0.002 | 0.44 ab | ±0.03 | 8.5 ab | ±0.4 | |
| 4–8 | 0.056 c | ±0.001 | 0.46 b | ±0.01 | 8.2 b | ±0.3 | 0.049 b | ±0.001 | 0.39 b | ±0.01 | 8.0 b | ±0.1 | |
| 8–12 | 0.054 c | ±0.001 | 0.44 b | ±0.01 | 8.2 b | ±0.3 | 0.048 b | ±0.001 | 0.38 b | ±0.01 | 7.9 b | ±0.1 | |
| 65–75 | 0–2 | 0.068 a | ±0.002 | 0.61 a | ±0.02 | 9.1 a | ±0.2 | 0.059 a | ±0.002 | 0.50 a | ±0.03 | 8.4 a | ±0.2 |
| 2–4 | 0.056 b | ±0.001 | 0.45 b | ±0.01 | 8.2 b | ±0.2 | 0.053 ab | ±0.002 | 0.43 ab | ±0.02 | 8.1 a | ±0.2 | |
| 4–8 | 0.052 bc | ±0.001 | 0.40 c | ±0.01 | 7.7 bc | ±0.2 | 0.050 b | ±0.001 | 0.38 b | ±0.01 | 7.8 a | ±0.2 | |
| 8–12 | 0.050 c | ±0.001 | 0.38 c | ±0.01 | 7.6 c | ±0.2 | 0.048 b | ±0.002 | 0.37 b | ±0.01 | 7.5 a | ±0.2 | |
| 75–85 | 0–2 | 0.068 a | ±0.002 | 0.59 a | ±0.03 | 8.6 a | ±0.2 | 0.058 a | ±0.002 | 0.50 a | ±0.03 | 8.5 a | ±0.2 |
| 2–4 | 0.054 b | ±0.001 | 0.41 b | ±0.01 | 7.7 b | ±0.1 | 0.051 ab | ±0.002 | 0.41 ab | ±0.02 | 8.0 ab | ±0.3 | |
| 4–8 | 0.051 c | ±0.001 | 0.38 b | ±0.01 | 7.4 b | ±0.2 | 0.048 bc | ±0.001 | 0.37 b | ±0.01 | 7.7 b | ±0.3 | |
| 8–12 | 0.050 c | ±0.001 | 0.37 b | ±0.01 | 7.3 b | ±0.2 | 0.046 c | ±0.001 | 0.35 b | ±0.01 | 7.7 b | ±0.3 | |
| 85–95 | 0–2 | 0.064 a | ±0.002 | 0.57 a | ±0.03 | 9.0 a | ±0.5 | 0.056 a | ±0.002 | 0.48 a | ±0.03 | 8.4 a | ±0.3 |
| 2–4 | 0.051 b | ±0.001 | 0.43 ab | ±0.03 | 8.3 b | ±0.6 | 0.048 ab | ±0.001 | 0.38 ab | ±0.02 | 7.9 b | ±0.3 | |
| 4–8 | 0.048 b | ±0.001 | 0.37 b | ±0.02 | 7.6 b | ±0.4 | 0.045 bc | ±0.001 | 0.35 b | ±0.01 | 7.7 b | ±0.2 | |
| 8–12 | 0.045 c | ±0.001 | 0.35 b | ±0.02 | 7.7 b | ±0.4 | 0.044 c | ±0.001 | 0.33 b | ±0.01 | 7.6 b | ±0.3 | |
| 95–105 | 0–2 | 0.060 a | ±0.002 | 0.53 a | ±0.04 | 8.7 a | ±0.3 | 0.056 a | ±0.002 | 0.48 a | ±0.03 | 8.6 a | ±0.3 |
| 2–4 | 0.047 b | ±0.002 | 0.39 ab | ±0.02 | 8.1 b | ±0.4 | 0.047 b | ±0.001 | 0.37 ab | ±0.01 | 7.8 b | ±0.3 | |
| 4–8 | 0.042 c | ±0.002 | 0.33 b | ±0.01 | 7.2 c | ±0.1 | 0.045 b | ±0.001 | 0.34 b | ±0.01 | 7.6 bc | ±0.3 | |
| 8–12 | 0.040 c | ±0.002 | 0.32 b | ±0.01 | 7.3 c | ±0.2 | 0.044 b | ±0.001 | 0.32 b | ±0.01 | 7.5 c | ±0.3 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Petzoldt, L.; Athmann, M.; Buechse, A.; Kautz, T. Root Growth of Hordeum vulgare and Vicia faba in the Biopore Sheath. Agriculture 2020, 10, 650. https://doi.org/10.3390/agriculture10120650
Petzoldt L, Athmann M, Buechse A, Kautz T. Root Growth of Hordeum vulgare and Vicia faba in the Biopore Sheath. Agriculture. 2020; 10(12):650. https://doi.org/10.3390/agriculture10120650
Chicago/Turabian StylePetzoldt, Lisa, Miriam Athmann, Andreas Buechse, and Timo Kautz. 2020. "Root Growth of Hordeum vulgare and Vicia faba in the Biopore Sheath" Agriculture 10, no. 12: 650. https://doi.org/10.3390/agriculture10120650
APA StylePetzoldt, L., Athmann, M., Buechse, A., & Kautz, T. (2020). Root Growth of Hordeum vulgare and Vicia faba in the Biopore Sheath. Agriculture, 10(12), 650. https://doi.org/10.3390/agriculture10120650