Isolation, Identification and Characterization of Rhizobacteria Strains for Biological Control of Bacterial Wilt (Ralstonia solanacearum) of Eggplant in China




Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Collection and Bacterial Isolation
2.2. In Vitro Assessment of Antagonistic Activity of Bacterial Strains against R. solanacearum
2.3. Identification of Bacterial Strains
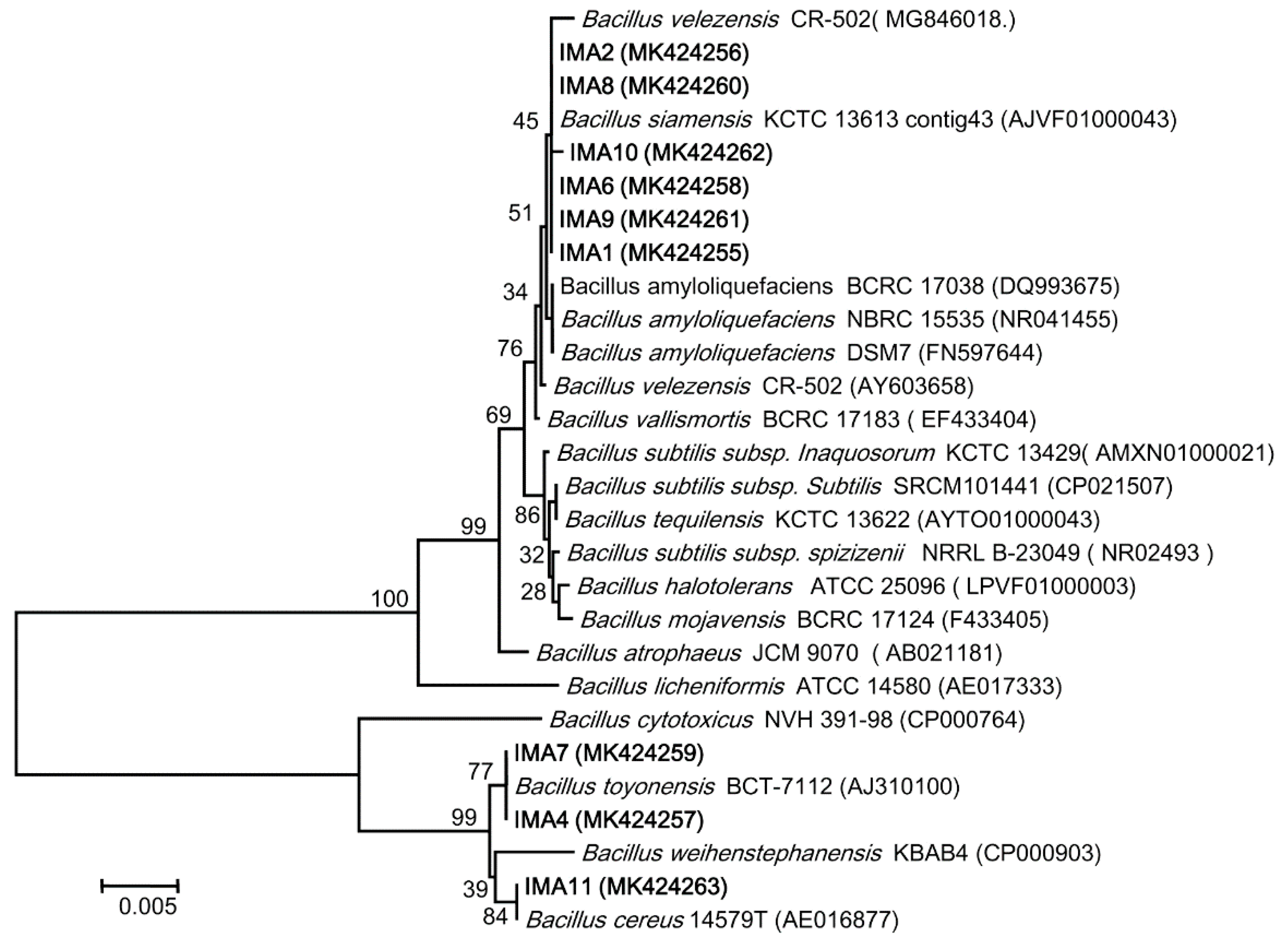
2.4. Phylogenetic Analyses of gyrA and rpoB Gene Sequences
2.5. Lipopeptides Detection by MALDI-TOF MS Analysis
2.6. Greenhouse Experiment of Biocontrol Effect of Selected Bacteria Strain on Severity Index of Eggplant Bacterial Wilt and Plant Biomass
2.7. Plant Growth Promotion
2.8. Statistical Analysis
3. Results
3.1. Isolation, Screening and Assessment of Bacterial Strains
3.2. Identification of Bacterial Strains
3.3. Lipopeptides Detection by MALDI-TOF MS Analyses
3.4. Effect of Different Strains on Eggplant Wilt and Plant Biomass
3.5. Eggplant Growth Promotion
4. Discussion
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Jung, E.-J.; Bae, M.-S.; Jo, E.-K.; Jo, Y.-H.; Lee, S.-C. Antioxidant activity of different parts of eggplant. J. Med. Plant Res. 2011, 5, 4610–4615. [Google Scholar]
- Gholizadeh, S.; Valipour-Chahardahcharic, S.; Sazegar, H. The effect of oral administration of eggplant hydroalcoholic extract on avoidance memory retrieval in streptozotocin-induced diabetic male rats. J. HerbMed Pharmacol. 2016, 4, 137–142. [Google Scholar]
- Lebeau, A.; Daunay, M.-C.; Frary, A.; Palloix, A.; Wang, J.-F.; Dintinger, J.; Chiroleu, F.; Wicker, E.; Prior, P. Bacterial wilt resistance in tomato, pepper, and eggplant: Genetic resources respond to diverse strains in the ralstonia solanacearum species complex. Phytopathology 2011, 101, 154–165. [Google Scholar] [CrossRef] [Green Version]
- Hayward, A.C. Characteristics of Pseudomonas solanacearum. J. Appl. Bacteriol. 1964, 27, 265–277. [Google Scholar]
- Hayward, C. Ralstonia solanacearum. Encycl. Microbiol. 2000, 4, 32–42. [Google Scholar]
- Chandrasekaran, J.; Brumin, M.; Wolf, D.; Leibman, D.; Klap, C.; Pearlsman, M.; Sherman, A.; Arazi, T.; Gal-On, A. Development of broad virus resistance in non-transgenic cucumber using CRISPR/Cas9 technology. Mol. Plant Pathol. 2016, 7, 1140–1153. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Almoneafy, A.A.; Kakar, K.U.; Nawaz, Z.; Li, B.; saand, M.A.; Chun-lan, Y.; Xie, G.-L. Tomato plant growth promotion and antibacterial related-mechanisms of four rhizobacterial Bacillus strains against Ralstonia solanacearum. Symbiosis 2014, 2, 59–70. [Google Scholar] [CrossRef]
- Li, B.; Su, T.; Yu, R.; Tao, Z.; Wu, Z.; Algam, S.A.E.; Xie, G.; Wang, Y.; Sun, G. Inhibitory activity of Paenibacillus macerans and Paenibacillus polymyxa against Ralstonia solanacearum. Afr. J. Microbiol. 2010, 4, 2048–2054. [Google Scholar]
- Hyakumachi, M.; Nishimura, M.; Arakawa, T.; Asano, S.; Yoshida, S.; Tsushima, S.; Takahashi, H. Bacillus thuringiensis suppresses bacterial wilt disease caused by Ralstonia solanacearum with systemic induction of defense-related gene expression in tomato. Microbes Environ. 2012, 28, 128–134. [Google Scholar] [CrossRef] [Green Version]
- Nakahara, H.; Mori, T.; Sadakari, N.; Matsusaki, H.; Matsuzoe, N. Selection of effective non-pathogenic Ralstonia solanacearum as biocontrol agents against bacterial wilt in eggplant. J. Plant Dis. Protect. 2016, 3, 119–124. [Google Scholar] [CrossRef]
- Price, N.P.J.; Rooney, A.P.; Swezey, J.L.; Perry, E.; Cohan, F.M. Mass spectrometric analysis of lipopeptides from Bacillus strains isolated from diverse geographical locations. FEMS Microbiol. Lett. 2007, 271, 83–89. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, W.; Wu, X.; Bai, X.; Zhang, H.; Dong, D.; Zhang, T.; Wu, H. Fusaricidins in Paenibacillus polymyxa A21 and their antagonistic activity against Botrytis cinerea on tomato. Front. Agric. Sci. Eng. 2018, 2, 262–270. [Google Scholar] [CrossRef] [Green Version]
- Guo, J.; Qi, H.; Guo, Y.; Ge, H.; Gong, L.; Zhang, L.; Sun, P. Biocontrol of tomato wilt by plant growth-promoting rhizobacteria. Biol. Control 2004, 29, 66–72. [Google Scholar] [CrossRef]
- Lemessa, F.; Zeller, W. Screening rhizobacteria for biological control of Ralstonia solanacearum in Ethiopia. Biol. Control 2007, 3, 336–344. [Google Scholar] [CrossRef]
- Ramesh, R.; Phadke, G.S. Rhizosphere and endophytic bacteria for the suppression of eggplant wilt caused by Ralstonia solanacearum. Crop Prot. 2012, 37, 35–41. [Google Scholar] [CrossRef]
- Kabaluk, T.; Gazdik, K. Directory of Microbial Pesticides for Agricultural Crops in OECD Countries; Agriculture and Agri-Food Canada: Ottawa, ON, Canada, 2007. [Google Scholar]
- Zhang, F.; Li, X.-L.; Zhu, S.-J.; Ojaghian, M.R.; Zhang, J.-Z. Biocontrol potential of Paenibacillus polymyxa against Verticillium dahliae infecting cotton plants. Biol. Control 2018, 127, 70–77. [Google Scholar] [CrossRef]
- Silva, M.; Cavalett, A.; Spinner, A.; Rosa, D.; Beltrame Jasper, R.; Quecine, M.; Letícia Bonatelli, M.; Pizzirani-Kleiner, A.; Corção, G.; Lima, A. Phylogenetic identification of marine bacteria isolated from deep-sea sediments of the eastern South Atlantic Ocean. SpringerPlus 2013, 2, 127. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7:molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 7, 1870–1874. [Google Scholar] [CrossRef] [Green Version]
- Jeon, Y.H.; Chang, S.P.; Hwang, I.Y.; Kim, Y.H. Involvement of growth-promoting rhizobacterium Paenibacillus polymyxa in root rot of stored Korean Ginseng. J. Microbiol. Biotechnol. 2003, 13, 881–891. [Google Scholar]
- Chun, J.; Bae, K.S. Phylogenetic analysis of Bacillus subtilis and related taxa based on partial gyrA gene sequences. Antonie Leeuwenhoek. 2000, 2, 123–127. [Google Scholar] [CrossRef]
- Qiu, Z.; Lu, X.; Li, N.; Zhang, M.; Qiao, X. Characterization of garlic endophytes isolated from the black garlic processing. Microbiol. Open 2017, 7, e00547. [Google Scholar] [CrossRef] [PubMed]
- Hanen, B.A.; Hmidet, N.; Béchet, M.; Chollet, M.; Chataigné, G.; Leclère, V.; Jacques, P.; Nasri, M. Identification and biochemical characteristics of lipopeptides from Bacillus mojavensis A21. Process Biochem. 2014, 10, 1699–1707. [Google Scholar]
- Winstead, N.N.; Kelman, A. Inoculation techniques for evaluating resistance to Pseudomonas solanacearum. PhytoPath 1952, 11, 628–634. [Google Scholar]
- Mrozik, A.; Łabużek, S.; Piotrowska-Seget, Z. Changes in fatty acid composition in Pseudomonas putida and Pseudomonas stutzeri during naphthalene degradation. Microbiol. Res. 2005, 2, 149–157. [Google Scholar] [CrossRef] [PubMed]
- Jiménez, G.; Urdiain, M.; Cifuentes, A.; López-López, A.; Blanch, A.R.; Tamames, J.; Kämpfer, P.; Kolstø, A.-B.; Ramón, D.; Martínez, J.F.; et al. Description of Bacillus toyonensis sp. nov., a novel species of the Bacillus cereus group, and pairwise genome comparisons of the species of the group by means of ANI calculations. Appl. Microbiol. 2013, 6, 383–391. [Google Scholar]
- Kämpfer, P. Limits and possibilities of total fatty acid analysis for classification and identification of bacillus species. Syst. Appl. Microbiol. 1994, 17, 86–98. [Google Scholar] [CrossRef]
- Berti, A.D.; Greve, N.J.; Christensen, Q.H.; Thomas, M.G. Identification of a biosynthetic gene cluster and the six associated lipopeptides involved in swarming motility of Pseudomonas syringae pv. tomato DC3000. J. Bacteriol. 2007, 17, 6312–6323. [Google Scholar] [CrossRef] [Green Version]
- Michelsen, C.F.; Watrous, J.; Glaring, M.A.; Kersten, R.; Koyama, N.; Dorrestein, P.C.; Stougaard, P. Nonribosomal peptides, key biocontrol components for Pseudomonas fluorescens In5, isolated from a Greenlandic suppressive soil. Am. Soc. Microbiol. 2015, 2, e00079. [Google Scholar] [CrossRef] [Green Version]
- Kuiper, I.; Lagendijk, E.L.; Pickford, R.; Derrick, J.P.; Lamers, G.E.M.; Thomas-Oates, J.E.; Lugtenberg, B.J.J.; Bloemberg, G.V. Characterization of two Pseudomonas putida lipopeptide biosurfactants, putisolvin I and II, which inhibit biofilm formation and break down existing biofilms. Mol. Microbiol. 2004, 51, 97–113. [Google Scholar] [CrossRef]
- Moskowitz, S.M.; Brannon, M.K.; Dasgupta, N.; Pier, M.; Sgambati, N.; Miller, A.K.; Selgrade, S.E.; Miller, S.I.; Denton, M.; Conway, S.P.; et al. PmrB mutations promote polymyxin resistance of Pseudomonas aeruginosa isolated from colistin-treated cystic fibrosis patients. Antimicrob. Agents Chemother. 2012, 2, 1019–1030. [Google Scholar] [CrossRef] [Green Version]
- Li, W.; Rokni-Zadeh, H.; De Vleeschouwer, M.; Ghequire, M.G.K.; Sinnaeve, D.; Xie, G.-L.; Rozenski, J.; Madder, A.; Martins, J.C.; De Mot, R. The antimicrobial compound xantholysin defines a new group of Pseudomonas cyclic lipopeptides. PLoS ONE 2013, 8, e62946. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Niu, D.; Wang, X.; Wang, Y.; Song, X.; Wang, J.; Guo, J.; Zhao, H. Bacillus cereus AR156 activates PAMP-triggered immunity and induces a systemic acquired resistance through a NPR1-and SA-dependent signaling pathway. Biochem. Biophys. Res. Commun. 2016, 1, 120–125. [Google Scholar] [CrossRef] [PubMed]
- Vater, J.; Herfort, S.; Doellinger, J.; Weydmann, M.; Borriss, R.; Lasch, P. Genome mining of the lipopeptide biosynthesis of paenibacillus polymyxa E681 in combination with mass spectrometry: Discovery of the Lipoheptapeptide Paenilipoheptin. Chem. BioChem. 2018, 7, 744–753. [Google Scholar] [CrossRef] [PubMed]
- Vater, J.; Niu, B.; Dietel, K.; Borriss, R. Characterization of novel fusaricidins produced by paenibacillus polymyxa-M1 using MALDI-TOF mass spectrometry. J. Am. Soc. 2015, 9, 1548–1558. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- AlMasoud, N.; Xu, Y.; Trivedi, D.K.; Salivo, S.; Abban, T.; Rattray, N.J.W.; Szula, E.; AlRabiah, H.; Sayqal, A.; Goodacre, R. Classification of Bacillus and Brevibacillus species using rapid analysis of lipids by mass spectrometry. Anal. Bioanal. Chem. 2016, 27, 7865–7878. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hayes, J.M.; Murray, K.K. Ambient laser ablation sample transfer with nanostructure-assisted laser desorption ionization mass spectrometry for bacteria analysis. Rapid Commun. Mass Spectrom. 2014, 21, 2382–2384. [Google Scholar] [CrossRef]
- Masum, M.M.I.; Liu, L.; Yang, M.; Hossain, M.M.; Siddiqa, M.M.; Supty, M.E.; Ogunyemi, S.O.; Hossain, A.; An, Q.; Li, B. Halotolerant bacteria belonging to operational group Bacillus amyloliquefaciens in biocontrol of the rice brown stripe pathogen Acidovorax oryzae. J. Appl. Microbiol. 2018, 6, 1852–1867. [Google Scholar] [CrossRef]
- Arwiyanto, T. Biological control of plant diseases caused by bacteria. J. Perlindungan Tanam. Indones. 2014, 18, 1–12. [Google Scholar]
- Essghaier, B.; Rouaissi, M.; Boudabous, A.; Jijakli, H.; Sadfi-Zouaoui, N. Production and partial characterization of chitinase from a halotolerant Planococcus rifitoensis strain M2-26. J. Microbiol. Biotechnol. 2010, 6, 977–984. [Google Scholar] [CrossRef]
- Sharma, A.; Meena, K.; Kanwar, S. Microbial lipopeptides and their medical applications. J. Pharm. Pharmacol. 2017, 2, 1126–1132. [Google Scholar]
- Cochrane, S.A.; Lohans, C.T.; van Belkum, M.J.; Bels, M.A.; Vederas, J.C. Studies on tridecaptin B1, a lipopeptide with activity against multidrug resistant Gram-negative bacteria. Org. Biomol. Chem. 2015, 13, 6073–6081. [Google Scholar] [CrossRef] [PubMed]
- Cochrane, S.A.; Vederas, J.C. Lipopeptides from Bacillus and Paenibacillus spp.: A gold mine of antibiotic candidates. Med. Res. Rev. 2016, 36, 4–31. [Google Scholar] [CrossRef] [PubMed]
- Velkov, T.; Thompson, P.; Nation, R.; Li, J. Structure-Activity relationships of polymyxin antibiotics. J. Med. Chem. 2009, 53, 1898–1916. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kurusu, K.; Ohba, K.; Arai, T.; Fukushima, K. New peptide antibiotics LI-F03, F04, F05, F07, and F08, produced by Bacillus polymyxa. I. Isolation and characterization. J. Antibiot. 1987, 40, 1506–1514. [Google Scholar]
- Chen, M.C.; Wang, J.P.; Zhu, Y.J.; Liu, B.; Yang, W.J.; Ruan, C.Q. Antibacterial activity against Ralstonia solanacearum of the lipopeptides secreted from the Bacillus amyloliquefaciens strain FJAT-2349. J. Appl. Microbiol. 2019, 5, 1519–1529. [Google Scholar] [CrossRef]
- Villegas-Escobar, V.; González-Jaramillo, L.M.; Ramírez, M.; Moncada, R.N.; Sierra-Zapata, L.; Orduz, S.; Romero-Tabarez, M. Lipopeptides from Bacillus sp. EA-CB0959: Active metabolites responsible for in vitro and in vivo control of Ralstonia solanacearum. Biol. Control 2018, 125, 20–28. [Google Scholar]
- Wang, N.; Wang, L.; Zhu, K.; Hou, S.; Chen, L.; Mi, D.; Gui, Y.; Qi, Y.; Jiang, C.; Guo, J.-H. Plant root exudates are involved in bacillus cereus AR156 mediated biocontrol against Ralstonia solanacearum. Front. Microbiol. 2019, 98, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Raza, S.; Ameen, A. Comparison of 16S rRNA gene of bacillus cereus with different bacterial species. J. Adv. Biol. Biotechnol. 2016, 7, 1–6. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Soil Samples | Origin | No. of Strains | Antagonistic Strains | Diameters of Inhibition Zones (mm) |
|---|---|---|---|---|
| 1 | Shangyu district, Shaoxing city | 42 | IMA7 | 17.33 ± 0.06 b |
| 2 | Shangyu district, Shaoxing city | 21 | IMA5 | 20.70 ± 0.85 a |
| 3 | Shangyu district, Shaoxing city | 27 | IMA11 | 17.93 ± 0.45 b |
| 4 | Shangyu district, Shaoxing city | 30 | IMA6 | 20.13 ± 0.13 a |
| 5 | Qliangfeeng town, Linan city | 20 | IMA8 | 20.20 ± 1.13 a |
| 6 | Qliangfeeng town, Linan city | 23 | IMA3 | 20.50 ± 0.04 a |
| 7 | Qliangfeeng town, Linan city | 25 | IMA10 | 20.00 ± 1.49 a |
| 8 | Yuhang district, Hangzhou city | 14 | IMA2 | 20.29 ± 0.11 a |
| 9 | Yuhang district, Hangzhou city | 23 | IMA4 | 18.00 ± 0.56 b |
| IMA9 | 15.00 ± 1.13 d | |||
| 10 | Yuhang district, Hangzhou city | 20 | IMA1 | 16.33 ± 1.30 c |
| Fatty Acid | Fatty Acid Content (%) | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| IMA 1 | IMA 2 | IMA 3 * | IMA 4 | IMA 5 | IMA 6 | IMA 7 | IMA 8 | IMA 9 | IMA 10 | IMA 11 | |
| Saturated straight-chain fatty acid | |||||||||||
| 12:0 | 0.72 | - | 2.27 | 0.56 | - | - | 1.42 | - | 1.12 | - | 0.68 |
| 14:0 | 3.50 | 3.51 | 0.33 | 3.40 | 1.31 | 1.33 | 3.45 | 2.14 | 3.70 | 1.65 | 3.72 |
| 16:0 | 6.87 | 19.70 | 24.58 | 6.86 | 9.98 | 8.94 | 7.22 | 15.84 | 7.69 | 12.33 | 7.31 |
| 18:0 | 0.69 | 0.82 | 0.74 | 0.48 | - | 0.68 | 0.53 | 0.62 | 0.72 | 0.61 | 0.69 |
| Saturated terminally branched fatty acid | |||||||||||
| 13:0 iso | 11.53 | 0.60 | - | 9.42 | - | 0.53 | 8.75 | 0.66 | 8.59 | 0.45 | 10.59 |
| 14:0 iso | 5.09 | 1.50 | - | 5.47 | 1.48 | 0.99 | 5.54 | 1.04 | 5.84 | 1.17 | 6.33 |
| 15:0 iso | 21.53 | 13.81 | 0.14 | 23.04 | 8.43 | 21.07 | 19.76 | 38.23 | 17.42 | 17.30 | 20.14 |
| 16:0 iso | 6.75 | 2.03 | - | 8.34 | 7.96 | 1.13 | 8.27 | 1.56 | 7.88 | 1.55 | 8.11 |
| 17:0 iso | 7.87 | 6.01 | 1.35 | 8.60 | 4.25 | 5.26 | 8.18 | 7.67 | 6.63 | 5.81 | 7.04 |
| 15:0 anteiso | 4.36 | 38.73 | - | 4.73 | 56.54 | 40.46 | 5.05 | 17.15 | 5.35 | 41.25 | 5.75 |
| 17:0 anteiso | 11.53 | 8.44 | - | 9.42 | 10.06 | 8.80 | 8.75 | 8.74 | 8.59 | 9.58 | 10.59 |
| Monounsaturated fatty acid | |||||||||||
| 16:1 w11c | 1.18 | 3.47 | - | 1.08 | - | 6.60 | 1.25 | 4.63 | 1.39 | 5.61 | 1.49 |
| 17:1ω10c iso | 4.87 | 0.64 | - | 4.27 | - | 1.53 | 4.33 | 1.11 | 4.46 | 1.09 | 4.01 |
| Inoculated Strains | a Disease Severity (%) | b Disease Incidence (%) | c Control Efficacy (%) |
|---|---|---|---|
| IMA2+YY06 | 30.0 ± 2.3 c | 46.7 ± 2.6 c | 46.2 c |
| IMA3+YY06 | 26.4 ± 1.0 d | 36.7 ± 2.6 d | 57.7 b |
| IMA4+YY06 | 40.7 ± 1.9 b | 60.0 ± 5.3 b | 30. 8 d |
| IMA5+YY06 | 21.7 ± 1.3 e | 26.7 ± 2.6 e | 69.2 a |
| IMA8+YY06 | 30.8 ± 1.6 c | 50.0 ± 5.3 c | 42.3 c |
| YY06 | 77.5 ± 2.3 a | 86.7 ± 5.3 a |
| Strains | Seedling Length (cm) | GPE (%) | Root Length (cm) | GPE (%) | Seedling Fresh Weight (g) | GPE (%) | Seedling Dry Weight (g) | GPE (%) | Root Fresh Weight (g) | GPE (%) | Root Dry Weight (g) | GPE (%) |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| IMA2+YY06 | 11.80 ± 0.01 | 26.4 c | 9.11 ± 0.03 | 47.0 c | 7.80 ± 0.00 | 67.7 b | 0.73 ± 0.00 | 84.0 c | 0.83 ± 0.01 | 59.6 c | 0.13 ± 0.00 | 45.4 c |
| IMA3+YY06 | 12.70 ± 0.01 | 36.1 b | 10.01 ± 0.22 | 61.6 b | 8.90 ± 0.00 | 89.5 a | 0.89 ± 0.02 | 122.5 b | 0.90 ± 0.01 | 73.0 b | 0.16 ± 0.00 | 77.7 b |
| IMA4+YY06 | 10.51 ± 0.01 | 12.6 d | 8.00 ± 0.27 | 29.0 d | 5.50 ± 0.11 | 17.0 c | 0.50 ± 0.01 | 25.0 d | 0.67 ± 0.01 | 28.8 d | 0.10 ± 0.00 | 11.1 d |
| IMA5+YY06 | 13.79 ± 0.00 | 44.2 a | 10.49 ± 0.01 | 69.2 a | 9.01 ± 0.01 | 91.7 a | 0.99 ± 0.01 | 147.3 a | 0.98 ± 0.01 | 88.5 a | 0.1 ± 0.00 | 87.8 a |
| IMA8+YY06 | 11.78 ± 0.00 | 26.3 c | 9.03 ± 0.01 | 45.6 c | 7.76 ± 0.02 | 68.5 b | 0.72 ± 0.01 | 80.00 c | 0.81 ± 0.00 | 56.7 c | 0.1 ± 0.00 | 43.4 c |
| YY06 | 9.33 | 6.20 | 4.70 | 0.40 | 0.52 | 0.09 | 9.3 |
| Strains | Seedling Length (cm) | GPE (%) | Root Length (cm) | GPE (%) | Seedling Fresh Weight (g) | GPE (%) | Seedling Dry Weight (g) | GPE (%) | Root Fresh Weight (g) | GPE (%) | Root Dry Weight (g) | GPE (%) |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| IMA2 | 14.31 ± 0.09 | 21.9 c | 11.31 ± 0.01 | 15.9 c | 9.00 ± 0.87 | 47.7 c | 0.89 ± 0.02 | 36.9 c | 0.97 ± 0.02 | 29.3 c | 0.17 ± 0.00 | 30.8 c |
| IMA3 | 16.33 ± 0.01 | 39.6 b | 12.00 ± 0.43 | 22.9 b | 10.22 ± 0.01 | 67.8 b | 1.07 ± 0.01 | 64.6 b | 1.08 ± 0.02 | 42.7 b | 0.19 ± 0.00 | 46.2 b |
| IMA4 | 13.01 ± 0.51 | 11.3 d | 10.58± 0.39 | 8.4 d | 7.50 ± 0.01 | 23.1 d | 0.70 ± 0.01 | 7.6 d | 0.87 ± 0.03 | 15.9 d | 0.14 ± 0.01 | 7.7 d |
| IMA5 | 18.81 ± 0.00 | 60.8 a | 12.98 ± 0.04 | 32.9 a | 10.90 ± 0.04 | 78.9 a | 1.35 ± 0.05 | 107.6 a | 1.13 ± 0.01 | 50.6 a | 0.22 ± 0.01 | 69.2 a |
| IMA8 | 14.20 ± 0.11 | 21.4 c | 11.20 ± 0.11 | 14.7 c | 8.89 ± 0.04 | 45.9 c | 0.85 ± 0.02 | 30.7 c | 0.94 ± 0.02 | 25.3 c | 0.17 ± 0.00 | 30.8 c |
| water | 11.69 | 9.76 | 6.0 | 0.65 | 0.75 | 0.13 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Abd Alamer, I.S.; Tomah, A.A.; Li, B.; Zhang, J.-Z. Isolation, Identification and Characterization of Rhizobacteria Strains for Biological Control of Bacterial Wilt (Ralstonia solanacearum) of Eggplant in China. Agriculture 2020, 10, 37. https://doi.org/10.3390/agriculture10020037
Abd Alamer IS, Tomah AA, Li B, Zhang J-Z. Isolation, Identification and Characterization of Rhizobacteria Strains for Biological Control of Bacterial Wilt (Ralstonia solanacearum) of Eggplant in China. Agriculture. 2020; 10(2):37. https://doi.org/10.3390/agriculture10020037
Chicago/Turabian StyleAbd Alamer, Iman Sabah, Ali Athafah Tomah, Bin Li, and Jing-Ze Zhang. 2020. "Isolation, Identification and Characterization of Rhizobacteria Strains for Biological Control of Bacterial Wilt (Ralstonia solanacearum) of Eggplant in China" Agriculture 10, no. 2: 37. https://doi.org/10.3390/agriculture10020037
APA StyleAbd Alamer, I. S., Tomah, A. A., Li, B., & Zhang, J. -Z. (2020). Isolation, Identification and Characterization of Rhizobacteria Strains for Biological Control of Bacterial Wilt (Ralstonia solanacearum) of Eggplant in China. Agriculture, 10(2), 37. https://doi.org/10.3390/agriculture10020037