3.1. Crop Performance and Physical Traits of Cultivated Plants
In both harvests, the interaction between species and density was not significant for yield or physical traits of cultivated plants (
Table 1 and
Table 2). In the first harvest, average leaf number, leaf weight and leaf area were 3.2 plant
−1, 660 mg leaf
−1 and 71.7 cm
2 plant
−1, respectively (
Table 1). At density D1 we obtained a yield 57% higher and dry weight 20% lower than at D2;
U. delechampii showed a higher SPAD index (
Table 1). In the second harvest the average yield was 1441 g m
−2, without differences in relation to species or density.
U. delechampii showed leaf number and SPAD that were 90% and 46% higher, respectively, than
U. picroides, while leaf weight and dry weight were 59% and 15% higher in
U. picroides than in
U. delechampii. At density D2, leaf area was 40% higher than at D1 (
Table 2).
In evaluating the two species for the ready-to-eat production chain, our results suggest the possibility of obtaining the same yield with both species, and highlight the possibility of using higher density (D1) to optimize yield in both harvests (
Table 1 and
Table 2). At the same time, the higher SPAD index of
U. delechampii, irrespective of plant density and harvest time, should be considered. We measured SPAD index to evaluate chlorophyll content indirectly, since this pigment is important not only for visual appearance but also for human health benefits (antioxidant and antimutagenic content) [
35]. From this point of view,
U. delechampii could be preferred to
U. picroides (
Table 1 and
Table 2).
From a crop performance point of view, crop cycle duration of the two species was shorter for
U. picroides, allowing earlier harvests than for
U. delechampii. The second harvest of
U. picroides was ready about 30 days before that of
U. delechampii (
Figure 1). The choice of
U. picroides could therefore have an economic advantage for the production chain, namely the same yield with a shorter crop cycle.
Finally, it is important to highlight that the leaves harvested from cultivated plants were clean (without soil particles) and, therefore, more suitable for the ready-to-eat production chain than wild plants (
Figure 2). This is because the floating system allows a high hygienic standard of vegetables, besides improving growth uniformity [
36].
3.2. Inorganic Ions Content
The interaction of species and density was not significant for ash or cations in the cultivated plants (
Table 3).
U. delechampii showed a sodium content 21% higher than
U. picroides, while potassium and calcium were higher in
U. picroides. At density D1, ash, sodium and potassium were 7.5%, 16% and 8% higher, respectively, than at density D2. Average magnesium content was 0.34 g 100 g
−1 without differences in relation to species and density (
Table 3).
Ash and magnesium contents of wild plants were 7.6 and 10.5% higher, respectively, in
U. picroides than
U. delechampii, while dry weight was 28% higher in
U. delechampii (
Table 4). Average contents of sodium, potassium and calcium were 1.06, 0.20 and 1.35 g 100 g
−1 d.w., respectively, without differences between species (
Table 4).
From the point of view of nutrition, we compared element intake per 100 g portion for cultivated and wild plants. For the cultivated plants, a 100 g portion of
U. delechampii provided an average of 48 mg sodium, 404 mg potassium, 20 mg magnesium and 91 mg calcium. The same serving of
U. picroides provided 37 mg sodium, 410 mg potassium, 20 mg magnesium and 88 mg calcium (
Table 1 and
Table 3). The wild plants provided 104 mg sodium, 293 mg potassium, 18 mg magnesium and 123 mg calcium per 100 g of
U. delechampii and 77 mg sodium, 234 mg potassium, 16 mg magnesium and 106 mg calcium per serving of
U. picroides (
Table 4). Potassium is an important element for humans, since it contributes to osmolarity and plays a major role in the distribution of fluids inside and outside cells. According to the European Food Safety Authority (EFSA) Panel on Dietetic Products, Nutrition and Allergies, the potassium Dietary Reference Value (DRV) is estimated at 3500 mg per day [
37]. Therefore, a serving of either cultivated
U. delechampii or
U. picroides supplies about 12% of the average daily requirement of this element, while a serving size of wild
U. delechampii or
U. picroides supplies 8.4% and 6.7%, respectively. It could therefore be interesting to compare the potassium content of the present species and other plants common in the ready-to-eat production chain. According to United States Department of Agriculture, 100 g of lettuce supplies 194 mg potassium [
38], while the same sized serving of arugula supplies 369 g potassium [
39]. Thus, our results suggest that the cultivated species of the present study can be considered potassium-rich vegetables. When cultivated at density D1, a 100 g portion of our plants provided 376 mg potassium, and 436 mg potassium when cultivated at density D2 (
Table 1 and
Table 3). Our results therefore suggest that potassium content can be modulated by choosing the appropriate plant density. Regarding sodium, magnesium and calcium, we found similar percentages of daily requirements for
U. delechampii and
U. picroides, which were intermediate between those of lettuce and arugula [
38,
39]. According to García-Herrera and de Cortes Sánchez-Mata [
40], our results, therefore, suggest that
U. delechampii and
U. picroides may be considered as a potential food source of mineral elements in the daily diet. At density D2, phosphate content was 25% higher than at density D1, while
U. picroides showed the highest sulphate content (
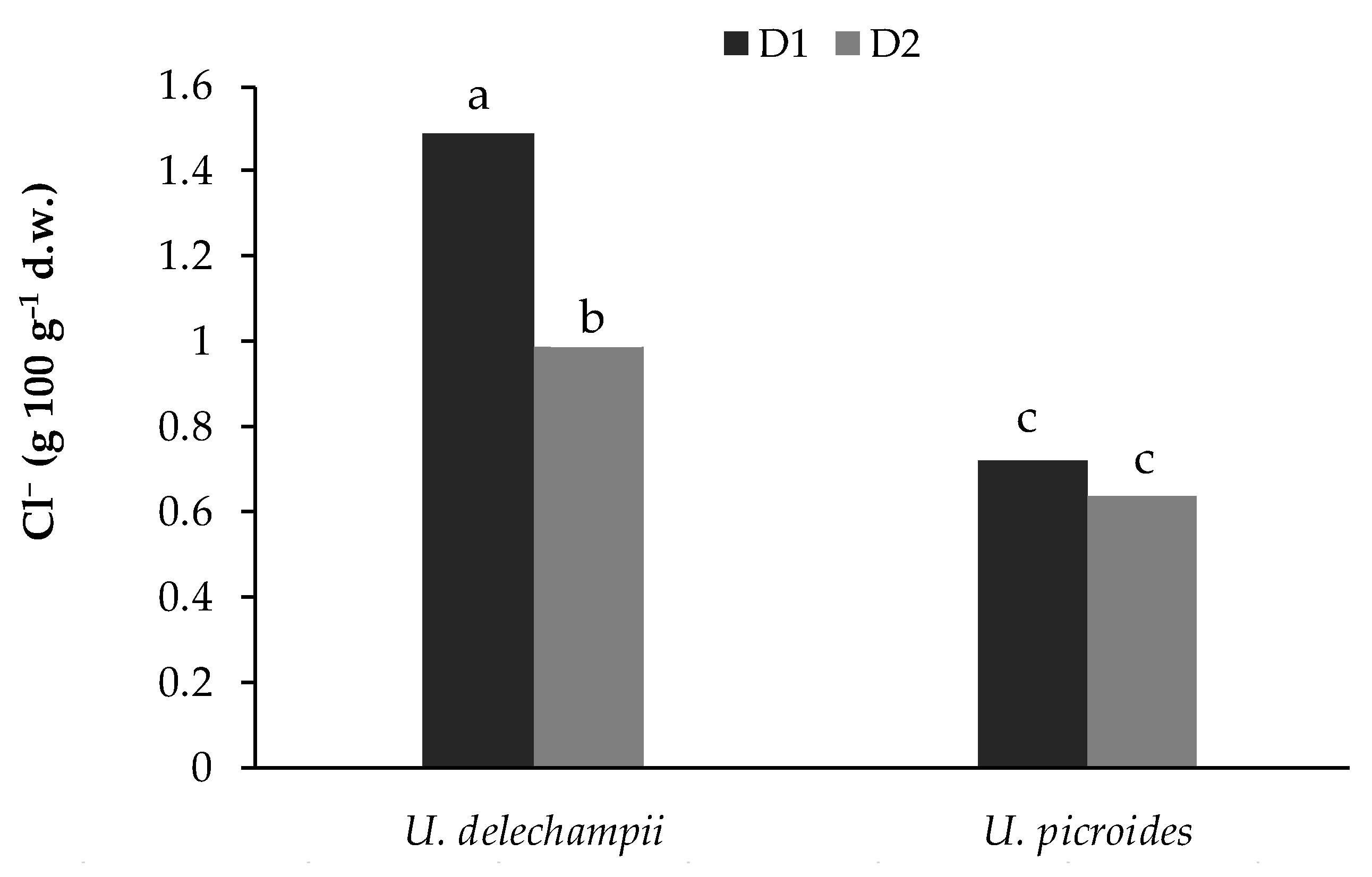
Table 5). The interaction between species and density was significant for nitrate and chlorine in the cultivated plants (
Table 5).
Cultivated at density D2,
U. delechampii had a nitrate content of about 7200 mg kg
−1 f.w., which is 38% higher than in plants of the same species grown at density D1 and in plants of
U. picroides (
Figure 3).
Grown at density D1,
U. delechampii showed a chloride content 50% higher than plants of the same species grown at density D2 and 119% higher than plants of
U. picroides (
Figure 4).
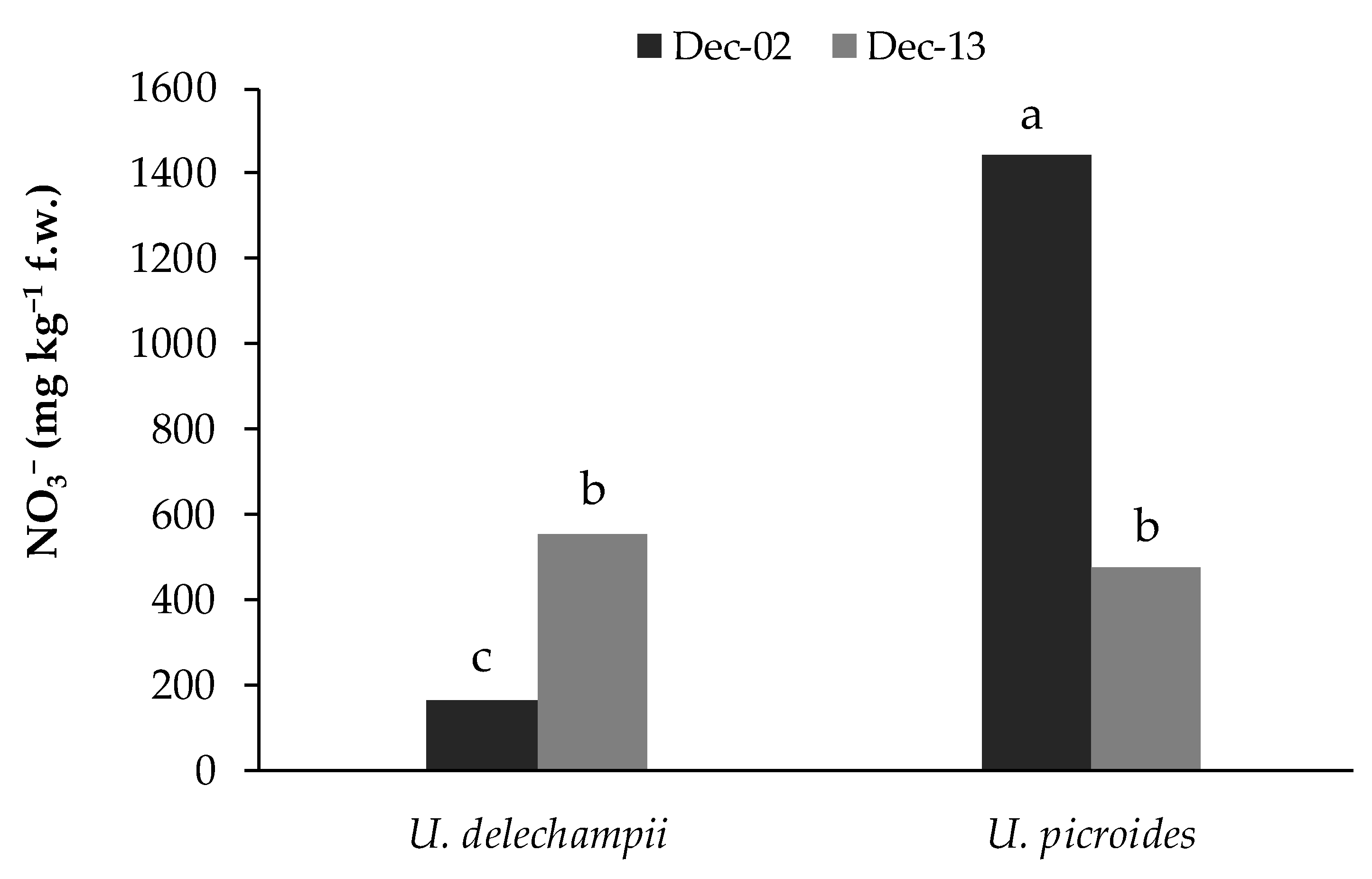
The interaction between species and harvest dates was significant for nitrate in wild plants (
Table 6). Average chloride, phosphate and sulphate content was 4.4, 0.45 and 0.48 g 100 g
−1 d.w., respectively, without differences between species or harvest dates (
Table 6).
U. picroides gathered on 2nd December showed a nitrate content of about 1450 mg kg
−1 f.w., 181% higher than that of plants gathered on 13th December and 8-fold higher than plants of
U. delechampii gathered on 2nd December (
Figure 5).
Nitrate content is an important quality parameter of vegetables, and among anions can be considered the most problematic due its possible implications for human health. Nitrate
per se is relatively non-toxic but on ingestion about 5% may be converted into nitrite and then N-nitroso compounds, which are linked to severe pathologies in humans [
41]. The European Commission adopted European Regulation (EU) no. 1258/2011 that sets maximum levels for nitrates in foodstuffs [
42]. For vegetables harvested from 1 October to 31 March (our study period), the Regulation set a maximum level of 7000 mg NO
3 kg
−1 f.w. for arugula and maximum levels of 5000 and 4000 mg NO
3 kg
−1 f.w. for lettuce grown under cover and in the open air, respectively [
42]. Our results show that wild
U. delechampii and
U. picroides have a substantially lower nitrate content than same species cultivated (
Figure 3 and
Figure 5). This is probably due to the higher availability of nitrate to plants grown in the floating system than to wild plants growing without exogenous fertilization. According to Santamaria [
41], nitrogen availability and plant genetic traits are two major factors that can affect nitrate content in vegetables. Indeed, in our study only
U. delechampii grown at density D2 showed a higher nitrate content as a consequence of nitrogen availability and it was twice as high as in plants grown at density D1 (
Figure 3). This is because for the same nitrogen supplied to the floating system, halving the number of plants increases nitrogen availability. Moreover, higher nitrate content in cultivated plants is also probably due to lower light intensity in the greenhouse. This is in line with Buttaro et al. [
23], who found a higher nitrate content in arugula grown in a greenhouse covered with photovoltaic modules (not transparent) than the same species grown in a greenhouse covered with transparent modules. In the interests of quality, strategies such as removing part of leaf petioles; removing part or all nitrate nitrogen from the nutrient solution a few days before harvesting; using nutrient solutions with NO
3-N and NH
4-N rather than nitrate nitrogen alone [
43] could therefore be used to reduce nitrate content in cultivated
U. delechampii and
U. picroides for the ready-to-eat production chain.
3.3. Total Phenols and Antioxidant Activity
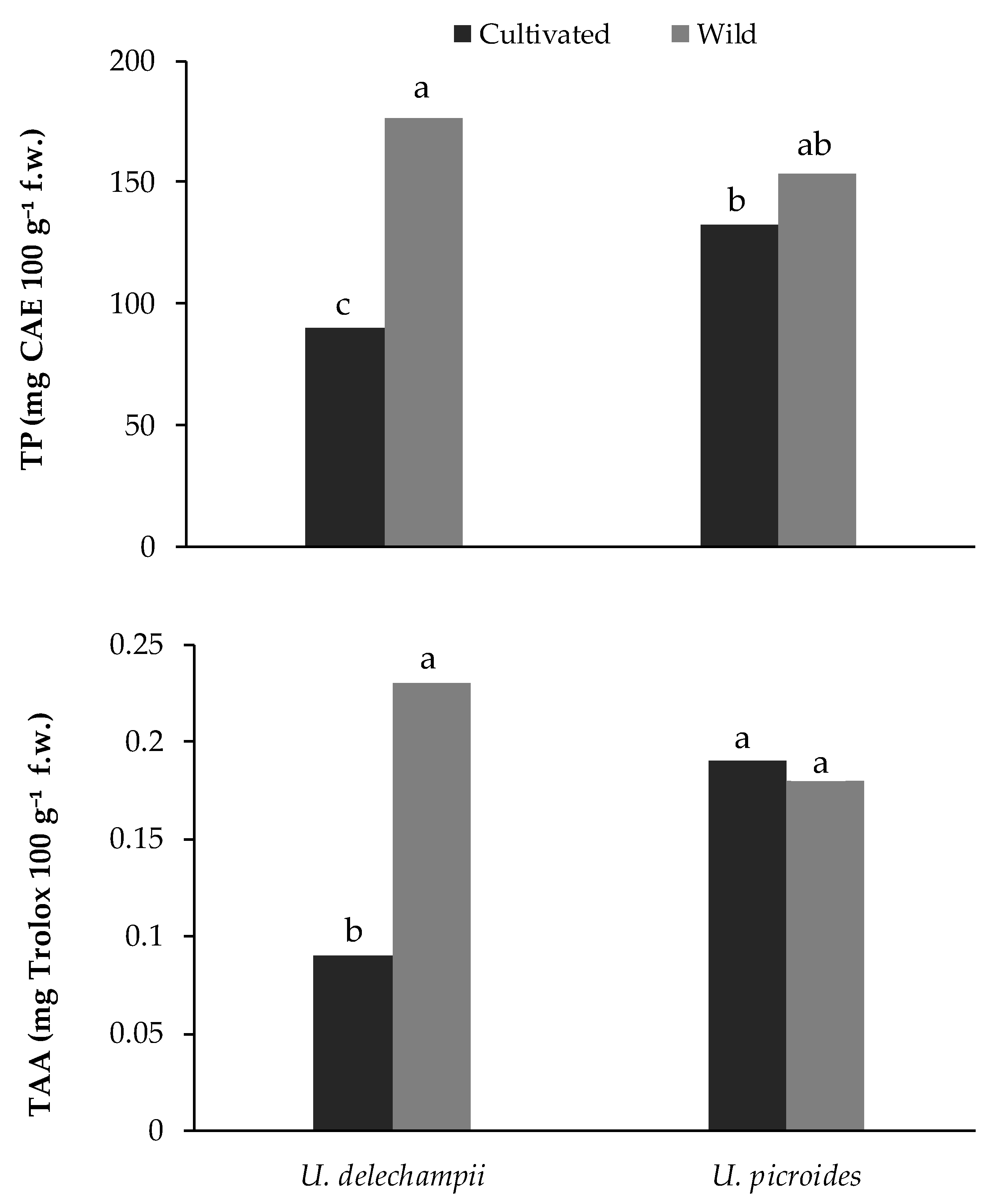
Wild plants of the two species gathered in grasslands did not show significantly different TP and TAA values (154 ± 20, 176 ± 22 mg CAE 100 g
−1 f.w. and 0.18 ± 0.03, 0.23 ± 0.05 g Trolox 100 g
−1 f.w. for
U. picroides and
U. dalechampii, respectively). As regards cultivated plants, the interaction between species and plant density was not significant (
Table 7).
U. dalechampii showed significantly lower TP (−32%) and TAA (−53%) than
U. picroides. At the same time, TP and TAA contents were 18 and 33% higher, respectively, in plants grown at density D2 than D1 (
Table 7).
Greenhouse-grown
U. dalechampii also showed significantly lower values than wild plants (TP: −49%; TAA: −61%), while wild and greenhouse specimens of
U. picroides did not show significant differences (
Figure 6).
Our data showed a positive correlation (
r2 = 0.91) between TP and TAA values, as already reported by many authors, who showed significant correlations between phenol content and antioxidant activity in herbs and vegetables [
21,
44,
45,
46,
47]. In our trials, lower phenol content and antioxidant activity were found in cultivated than in wild plants, and were sharper and more significant in
U. dalechhampii than in
U. picroides, thus demonstrating higher sensitivity of the former species regarding this aspect. The effect can probably be ascribed to lower luminosity in the greenhouse, in the absence of supplementary lighting, than in the open field, due to the sunlight filtering effect of the glass roof. Light is known to be a major environmental factor for the biosynthesis of phenols in plants. The physical characteristics of light used for supplementary lighting (e.g. intensity, wavelength, duration, photoperiod, etc.) play a fundamental role in eliciting as well as orientating plant metabolism, especially biosynthesis of flavonoids [
48], which are reported to be common in wild edible herbs [
20,
21,
45].
From a nutritional point of view, it is well known that phenolic compounds have an important positive role for humans. Effectively, these compounds can prevent malignancies by inhibiting the formation of nitrosamines or even decreasing their capacity for action, when they are formed. Moreover, they have antioxidant properties to be effective in preventing oxidation of the fraction low-density lipoprotein (LDL) cholesterol, thereby preventing atherosclerosis and other cardiovascular diseases [
49]. Most Mediterranean wild edible plants have an average total phenol content of less than or equal to 100 mg 100 g f.w. [
49]. Therefore, our results highlight that: (1) both
U. dalechhampii and
U. picroides can be considered a good source of phenols; (2) the total phenols content of cultivated plants in our study results similar or higher than other Mediterranean wild plants.


 ,
, 







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}