The Dynamic of Nitrogen Uptake from Different Sources by Pea (Pisum sativum L.)


Abstract
:1. Introduction
2. Materials and Methods
2.1. Field Experiment
2.2. Laboratory Analyses
- −
- dry matter (d.m.) content—70 °C
- −
- total nitrogen content—Kjeldahl method
- −
- enrichment with the 15N isotope—after wet mineralization by the Kiejdahl method and distillation to an acid solution (5% HCl), on the NOI-6e emission spectrometer (Leipzig, Germany).
2.3. Weather Conditions
2.4. Calculations of Nitrogen Sources
- the percentage of nitrogen derived from the atmosphere (%Ndfa):%Ndfa = 1 − (%15Nenrichment_fx/%15Nenrichemnt_nfx)
- −
- % 15N enrichment_fx-15N isotope enrichment in pea;
- −
- % 15N enrichment_nfx-15N isotope enrichment in the references plant—barley;
- the percentage of nitrogen derived from the fertilizer (%Ndff):%Ndff = %15Nenrichment_fx/%15Nenrichment_fertilizer
- −
- % 15N enrichment_fx-15N isotope enrichment in pea;
- −
- % 15N enrichment_fertilizer-15N isotope enrichment of fertilizer;
- the percentage of nitrogen derived from soil (%Ndfs):%Ndfs = 100 − (%Ndfa − %Ndff)
- −
- %Ndfa—% of nitrogen derived from the atmosphere;
- −
- %Ndff—% of nitrogen derived from the fertilizer.
2.5. Statistical Analysis
3. Results
3.1. Dry Weight of Pea Plants
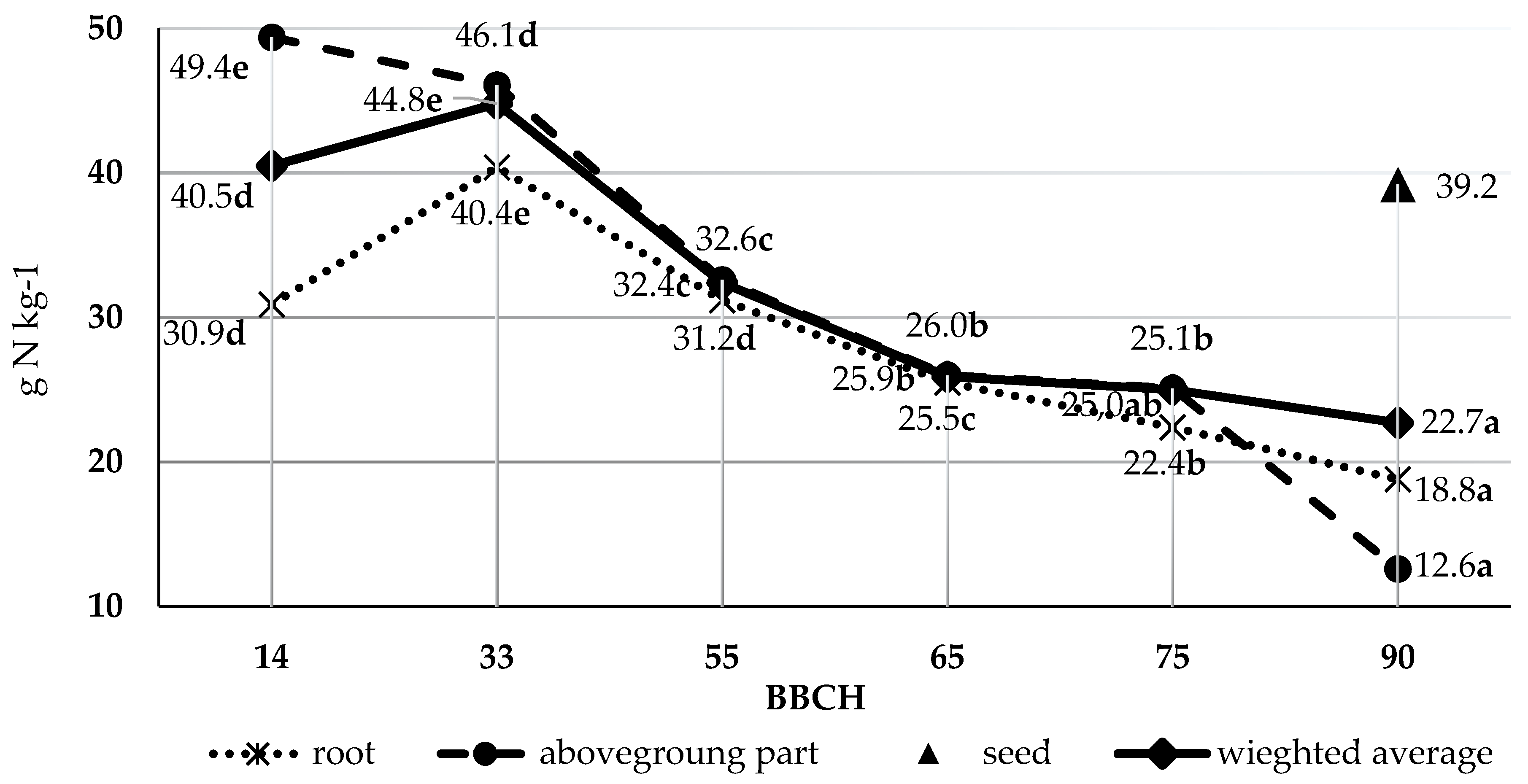
3.2. Nitrogen Content in Dry Wieght of Pea
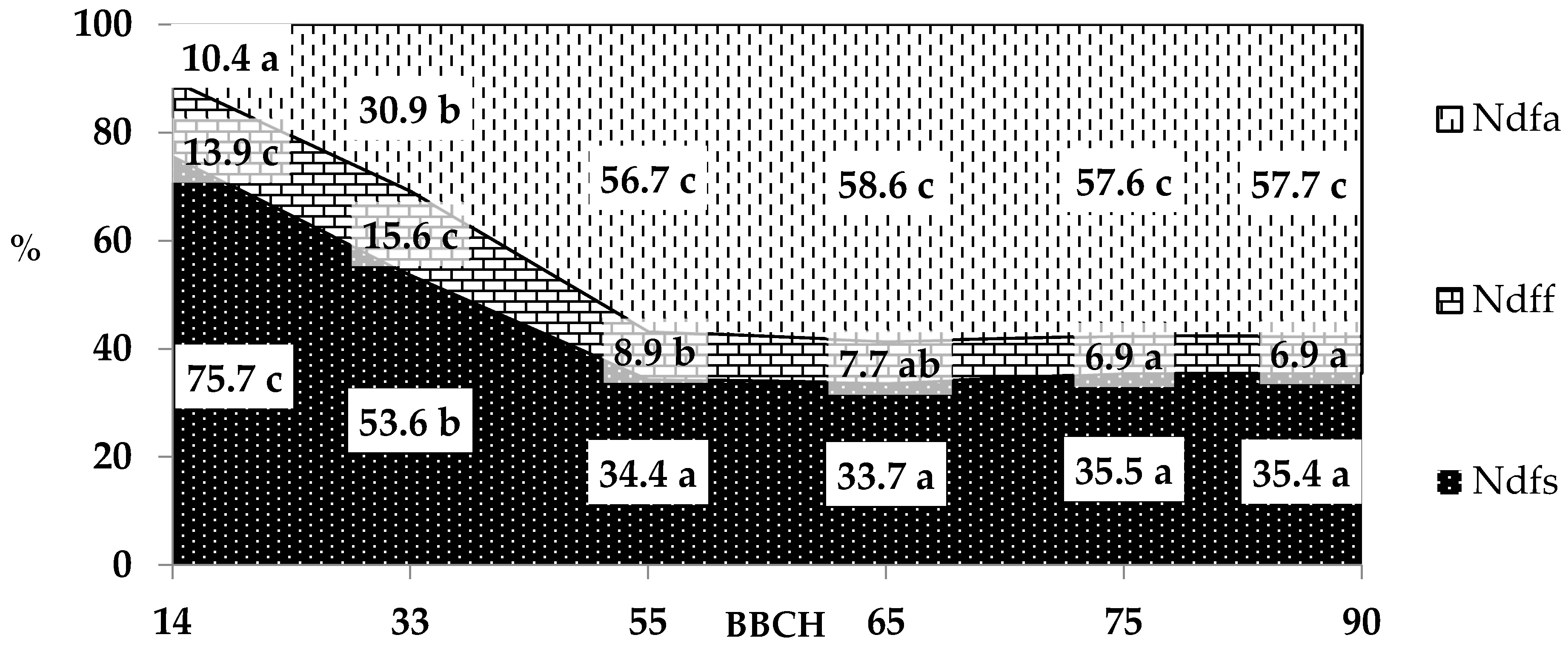
3.3. The Amount of Nitrogen Uptake from Different Sources
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Fowler, C.J.E.; Condron, L.M.; McLenaghen, R.D. Effects of green manures on nitrogen loss and availability in an organic cropping system. N. Z. J. Agric. Res. 2004, 47, 95–100. [Google Scholar] [CrossRef]
- Stagnari, F.; Maggio, A.; Galieni, A.; Pisante, M. Multiple benefits of legumes for agriculture sustainability: An overview. Chem. Biol. Technol. Agric. 2017, 4, 2. [Google Scholar] [CrossRef] [Green Version]
- Uzoh, I.; Igwe, C.; Okebalama, C.B.; Babalola, O.O. Legume-maize rotation effect on maize productivity and soil fertility parameters under selected agronomic practices in a sandy loam soil. Sci. Rep. 2019, 9, 8539. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, C.; Bueckert, R.; Schoenau, J.; Diederichsen, A.; Zakeri, H.; Warkentin, T.D. Evaluation of growth and nitrogen fixation of pea nodulation mutants in western Canada. Can. J. Plant Sci. 2017, 97, 1121–1129. [Google Scholar] [CrossRef] [Green Version]
- Hong, Y.; Ma, Y.; Wu, L.; Maki, M.; Qin, W.; Chen, S. Characterization and analysis of nifH genes from Paenibacillus sabinae T27. Microbiol. Res. 2012, 167, 596–601. [Google Scholar] [CrossRef]
- Ishizuka, J. Trends in biological nitrogen fixation research and application. Plant Soil 1992, 141, 197–209. [Google Scholar] [CrossRef]
- Kozłowski, S.; Swędrzyński, A.; Zielewicz, W. Leguminous plants in natural environment. Water Environ. Rural Areas 2011, 11, 161–181. [Google Scholar]
- Martyniuk, S. Scientific and practical aspects of legumes symbiosis with root-nodule bacteria. Pol. J. Agron. 2012, 9, 17–22. [Google Scholar]
- Peoples, M.B.; Craswell, E.T. Biological nitrogen fixation: Investments, expectations, and actual contributions to agriculture. Plant Soil 1992, 141, 13–39. [Google Scholar] [CrossRef]
- Cheng, Q. Perspectives in biological nitrogen fixation research. J. Integr. Plant Biol. 2008, 50, 784–796. [Google Scholar] [CrossRef]
- Duzdemir, O.; Kurunc, A.; Unlukara, A. Response of pea (Pisum sativum) to salinity and irrigation water regime. Bulg. J. Agric. Sci. 2009, 15, 400–409. [Google Scholar]
- Hauggaard-Nielsen, H.; Holdensen, L.; Wulfsohn, D.; Jensen, E.S. Spatial variation of N2-fixation in field pea (Pisum sativum L.) at the field scale determined by the 15N natural abundance method. Plant Soil 2010, 327, 167–184. [Google Scholar] [CrossRef]
- Reiter, K.; Schmidtke, K.; Rauber, R. The influence of long-term tillage systems on symbiotic N2 fixation of pea (Pisum sativum L.) and red clover (Trifolium pratense L.). Plant Soil 2002, 238, 41–55. [Google Scholar] [CrossRef]
- Karahne, V.; Singh, V.P. Effect of rhizobial inoculation on growth, yield, nodulation and biochemical characters of vegetable pea (Pisum sativum L.). Acta Agron. Hung. 2009, 57, 47–56. [Google Scholar] [CrossRef]
- Bonilla, I.; Bolańos, L. Mineral nutrition for legume-rhizobia symbiosis: B, Ca, N, P, S, K, Fe, Mo, Co, and Ni: A review. In Organic Farming, Pest Control and Remediation of Soil Pollutants; Lichtfouse, E., Ed.; Springer: Dordrecht, The Netherlands, 2010; pp. 253–274. [Google Scholar] [CrossRef]
- Schwember, A.R.; Schulze, J.; del Pozo, A.; Cabeza, R.A. Regulation of symbiotic nitrogen fixation in legume root nodules. Plants 2019, 8, 333. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, L.; Baddeley, J.; Watson, C. Models of biological nitrogen fixation of legumes. A review. Agron. Sustain. Dev. 2011, 31, 155–172. [Google Scholar]
- Rymuza, K.; Radzka, E.; Wysokinski, A. Nitrogen uptake from different sources by non-GMO soybean varieties. Agronomy 2020, 10, 1219. [Google Scholar] [CrossRef]
- Bourion, V.; Laguerre, G.; Depret, G.; Voisin, A.S.; Salon, C.; Duc, G. Genetic variability in nodulation and root growth affects nitrogen fixation and accumulation in pea. Ann. Bot. 2007, 100, 589–598. [Google Scholar] [CrossRef]
- Abi-Ghanem, R.; Carpenter-Boggs, L.; Smith, J.L. Cultivar effects on nitrogen fixation in peas and lentils. Biol. Fertil. Soils 2011, 47, 115–120. [Google Scholar] [CrossRef]
- Abi-Ghanem, R.; Bodah, E.T.; Wood, M.; Braunwart, K. Potential breeding for high nitrogen fixation in Pisum sativum L.: Germplasm phenotypic characterization and genetic investigation. Am. J. Plant Sci. 2013, 4, 1597–1600. [Google Scholar] [CrossRef] [Green Version]
- Kumar, K.; Goh, K.M. Biological nitrogen fixation, accumulation of soil nitrogen and nitrogen balance for white clover (Trifolium repens L.) and field pea (Pisum sativum L.) grown for seed. Field Crops Res. 2000, 68, 49–59. [Google Scholar] [CrossRef]
- Wysokinski, A.; Kalembasa, S.; Symanowicz, B. Dynamic of nitrogen accumulation by pea (Pisum sativum L.) from different sources. Fragm. Agron. 2013, 30, 162–169. [Google Scholar]
- Bleinholder, H.; Weber, E.; Feller, C.; Hess, M.; Wicke, H.; Meier, U.; van den Boom, T.; Lancashire, P.D.; Buhr, L.; Hack, H.; et al. Growth stages of mono- and dicotyledonous plants. In BBCH Monograph; Meier, U., Ed.; JKI: Quedlinburg, Germany, 2001; pp. 1–160. [Google Scholar]
- Kalembasa, S.; Wysokinski, A.; Kalembasa, D. Quantitative assessment of the process of biological nitrogen reduction by yellow lupine (Lupinus luteus L.). Acta Sci. Pol. Agric. 2014, 13, 5–20. [Google Scholar]
- Dzieżyc, J.; Nowak, L.; Panek, K. Ten-day indices of rainfall requirements of crops cultivated in Poland. Adv. Agric. Sci. Probl. Iss. 1987, 31, 11–33. (In Polish) [Google Scholar]
- Chmura, K.; Chylińska, E.; Dmowski, Z.; Nowak, L. The role of the water factor in shaping the yield of selected field plants. Infrastruct. Ecol. Rural Areas 2009, 9, 33–44. (In Polish) [Google Scholar]
- Azam, F.; Farooq, S. An appraisal of methods for measuring symbiotic nitrogen fixation in legumes. Pak. J. Biol. Sci. 2003, 6, 1631–1640. [Google Scholar] [CrossRef]
- McPhee, K.E.; Muehlbauer, F.J. Variation for biomass and residue production by dry pea. Field Crops Res. 1999, 62, 203–212. [Google Scholar] [CrossRef]
- Andrzejewska, J.; Wiatr, K.; Pilarczyk, W. Economic value of selected pea (Pisum sativum L.) cultivars on very good rye soil complex. Acta Sci. Pol. Agric. 2002, 1, 59–72. [Google Scholar]
- Poggio, S.L.; Satorre, E.H.; Dethiou, S.; Gonzallo, G.M. Pod and seed numbers as a function of photothermal quotient during the seed set period of field pea (Pisum sativum) crops. Eur. J. Agron. 2005, 22, 55–69. [Google Scholar] [CrossRef]
- Annicchiarico, P.; Iannucci, A. Adaptation strategy, germplasm type and adaptive traits for field pea improvement in Italy based on variety response across climatically contrasting environments. Field Crops Res. 2008, 108, 133–142. [Google Scholar] [CrossRef]
- Grabowska, K.; Banaszkiewicz, B. Effect of air temperature and atmospheric precipitation on yielding of sowing pea in central Poland. Acta Agroph. 2009, 13, 113–120. [Google Scholar]
- Woźniak, A. The yielding of pea (Pisum sativum L.) under different tillage conditions. Acta Sci. Pol. Hortorum Cultus 2013, 12, 133–141. [Google Scholar]
- Santín-Montanyá, M.I.; Zambrana, E.; Fernández-Getino, A.P.; Tenorio, J.I. Dry pea (Pisum sativum L.) yielding and weed infestation response, under different tillage conditions. Crop Prot. 2014, 65, 122–128. [Google Scholar] [CrossRef]
- Podleśny, J.; Podleśna, A. Effect of high temperature in the flowering period on pea growth, development and yielding. Adv. Agric. Sci. Problem Iss. 2010, 545, 73–78. [Google Scholar]
- Tanguilig, V.C.; Yambao, E.B.; O’toole, J.C.; De Datta, S.K. Water stress effects on leaf elongation, leaf water potential, transpiration and nutrient uptake of rice, maize and soybean. Plant Soil 1987, 103, 155–168. [Google Scholar] [CrossRef]
- Herdina, L.; Silsbury, J.H. Growth, nitrogen accumulation and partitioning, and N2 fixation in faba bean (Vicia faba cv. Fiord) and pea (Pisum sativum cv. Early Dun). Field Crops Res. 1990, 24, 173–188. [Google Scholar] [CrossRef]
- Jensen, E.S. Seasonal patterns of growth and nitrogen fixation in field-grown pea. Plant Soil 1987, 101, 29–37. [Google Scholar] [CrossRef]
- Marrou, H.; Ricaute, J.J.; Ghanem, M.E.; Clavijo Michelangeli, J.A.; Ghaouti, L.; Rao, I.M.; Sinclair, T.R. Is nitrogen accumulation in grain legumes responsive to growth or ontogeny? Physiol. Plant. 2018, 162, 109–122. [Google Scholar] [CrossRef] [Green Version]
- Schiltz, S.; Munier-Jolain, N.; Jeudy, C.; Burstin, J.; Salon, C. Dynamics of exogenous nitrogen partitioning and nitrogen remobilization from vegetative organs in pea revealed by 15N in vivo labeling throughout seed filling. Plant Physiol. 2005, 137, 1463–1473. [Google Scholar] [CrossRef] [Green Version]
- Peoples, M.B.; Brockwell, J.; Herridge, D.F.; Rochester, I.J.; Alves, B.J.R.; Urquiaga, S.; Boddey, R.M.; Dakora, F.D.; Bhattarai, S.; Maskey, S.L.; et al. The contributions of nitrogen-fixing crop legumes to the productivity of agricultural systems. Symbiosis 2009, 48, 1–17. [Google Scholar] [CrossRef]
- Hungria, M.; Vargas, M.A.T. Environmental factors impacting N2 fixation in legumes grown in the tropics, with an emphasis on Brazil. Field Crops Res. 2000, 65, 151–164. [Google Scholar] [CrossRef]
- Meena, V.S.; Maurya, B.R.; Verma, J.P. Does a rhizospheric microorganism enhance K+ availability in agricultural soil. Microbiol. Res. 2014, 169, 337–347. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Bahadur, I.; Maurya, B.R.; Raghuwanshi, R.; Meena, V.S.; Singh, D.K.; Dixit, J. Does a plant growth-promoting rhizobacteria enhance agricultural sustainability? J. Pure Appl. Microbiol. 2015, 9, 715–724. [Google Scholar]
- Armstrong, E.L.; Pate, J.S.; Tennant, D. The field pea in south western Australia. Patterns of water use and root growth in genotypes of contrasting morphology and growth habit. Aust. J. Plant Physiol. 1994, 21, 517–532. [Google Scholar] [CrossRef]
- Huang, J.; Afshar, R.K.; Tao, A.; Chen, C. Efficacy of starter N fertilizer and rhizobia inoculant in dry pea (Pisum sativum L.) production in a semi-arid temperate environment. Soil Sci. Plant Nutr. 2017, 63, 248–253. [Google Scholar] [CrossRef] [Green Version]
- Clayton, G.W.; Rice, W.A.; Lupwayi, N.Z.; Johnston, A.M.; Lafond, G.P.; Grant, C.A.; Walley, F.L. Inoculant formulation and fertilizer nitrogen effects on field pea: Nodulation, N2 fixation and nitrogen partitioning. Can. J. Plant Sci. 2004, 84, 79–88. [Google Scholar] [CrossRef]
- Buttery, B.R.; Gibson, A.H. The effect of nitrate on the time course of nitrogen fixation and growth in Pisum sativum and Vicia faba. Plant Soil 1990, 127, 143–146. [Google Scholar] [CrossRef]
- Voisin, A.S.; Salon, C.; Munier-Jolain, N.G.; Ney, B. Quantitative effects of soil nitrate, growth potential and phenology on symbiotic nitrogen fixation of pea (Pisum sativum L.). Plant Soil 2002, 243, 31–42. [Google Scholar] [CrossRef]
- Van Kessel, C.; Hartley, C. Agricultural management of grain legumes: Has it led to an increase in nitrogen fixation? Field Crops Res. 2000, 65, 165–181. [Google Scholar] [CrossRef]
- Salvagiotti, F.; Cassman, K.G.; Specht, J.E.; Walters, D.T.; Weiss, A.; Dobermann, A. Nitrogen uptake, fixation and response to fertilizer N in soybeans: A review. Field Crops Res. 2008, 108, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Fustec, J.; Lesuffleur, F.; Mahieu, S.; Cliquet, J.B. Nitrogen rhizodeposition of legumes. A review. Agron. Sustain. Dev. 2010, 30, 57–66. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
| Soil Properties | Unit | Year | |
|---|---|---|---|
| 2015 | 2016 | ||
| pH1 mol·dm-3 KCl | - | 6.6 | 6.5 |
| Ctotal | g·kg−1 | 34.2 | 23.5 |
| Ntotal | 2.10 | 1.45 | |
| PEgnera-Rhiema | mg·kg−1 | 309.0 | 301.0 |
| KEgnera-Rhiema | 86.0 | 111.0 | |
| Fe1 mol·dm-3 HCl | 1327 | 1189 | |
| Mo1 mol·dm-3 HCl | 0.015 | 0.013 | |
| B1 mol·dm-3 HCl | 0.806 | 0.278 | |
| Month | Total Monthly Rainfall, mm | Average Monthly Temperatures, °C | ||||
|---|---|---|---|---|---|---|
| Year | Long-Term Mean 1981–2014 | Year | Long-Term Mean 1981–2014 | |||
| 2015 | 2016 | 2015 | 2016 | |||
| III | 53.1 | 46.4 | 29.6 | 4.8 | 3.3 | 2.0 |
| IV | 30.0 | 50.2 | 33.4 | 8.2 | 8.9 | 8.1 |
| V | 100.2 | 35.5 | 6.3 | 12.3 | 14.6 | 13.6 |
| VI | 43.3 | 55.6 | 72.9 | 16.5 | 18.1 | 16.3 |
| VII | 62.6 | 126.8 | 67.6 | 18.7 | 19.0 | 18.5 |
| Sum IV–VII | 236.1 | 268.1 | 234.2 | 13.9 | 15.1 | 14.1 |
| Month | Year | |||
|---|---|---|---|---|
| 2015 | 2016 | |||
| k | wm | k | wm | |
| IV | 1.2 | md | 1.9 | mw |
| V | 2.6 | vw | 0.8 | d |
| VI | 0.9 | d | 1.0 | d |
| VII | 1.1 | md | 2.2 | w |
| VIII | 0.2 | ed | 1.1 | md |
| Part of Plant | Growth Stages (BBCH) | |||||
|---|---|---|---|---|---|---|
| 14 | 33 | 55 | 65 | 75 | 90 | |
| Seed | 260.4 ± 79.1 | |||||
| Above ground part | 18.6 ± 3.7 a | 74.1 ± 12.1 a | 271.2 ± 62.3 b | 491.5 ± 151.0 d | 571.7 ± 129.3 e | 407.2 ± 81.8 c |
| Root | 14.8 ± 2.7 a | 20.9 ± 3.0 b | 24.9 ± 4.7 cd | 25.9 ± 4.3 d | 21.7 ± 4.6 bc | 13.3 ± 4.2 a |
| Total mass | 33.4 ± 2.4 a | 95.0 ± 12.3 b | 296.1 ± 65.7 c | 517.4 ± 154.3 d | 593.4 ± 133.2 e | 680.8 ± 147.0 f |
| Part of Plant | Source of Variation | |||
|---|---|---|---|---|
| Pea Cultivar | Year of Research | |||
| ‘Milwa’ | ‘Batuta’ | 2015 | 2016 | |
| Seed | 259.5 ± 89.9 a | 261.2 ± 65.5 a | 334.5 ± 35.8 b | 186.1 ± 14.5 a |
| Aboveground part | 409.1 ± 101.2 a | 405.4 ± 55.9 a | 461.3 ± 55.2 b | 353.2 ± 67.0 a |
| Root | 13.9 ± 5.1 a | 12.7 ± 3.1 a | 016.6 ± 3.5 b | 10.0 ± 1.5 a |
| Total mass | 682.5 ± 183.6 a | 679.3 ± 97.5 a | 812.4 ± 63.2 b | 549.3 ± 67.8 a |
| Part of Plant | Source of Variation | |||
|---|---|---|---|---|
| Pea Cultivar | Year of Research | |||
| ‘Milwa’ | ‘Batuta’ | 2015 | 2016 | |
| Seed | 40.1 ± 1.8 a | 38.3 ± 0.8 a | 39.2 ± 1.9 a | 39.2 ± 1.4 a |
| Aboveground part | 11.8 ± 1.4 a | 13.5 ± 1.4 a | 12.2 ± 1.6 a | 13.1 ± 1.5 a |
| Root | 18.6 ± 1.6 a | 19.0 ± 2.2 a | 20.5 ± 1.0 b | 17.1 ± 0.5 a |
| Weighted average | 22.6 ± 2.0 a | 23.0 ± 2.1 a | 23.5 ± 1.8 a | 22.0 ± 2.0 a |
| N Source | Part of Plant | Growth Stages (BBCH) | |||||
|---|---|---|---|---|---|---|---|
| 14 | 33 | 55 | 65 | 75 | 90 | ||
| Ndfa
(air) | Seed | 61.1 ± 24.0 | |||||
| Aboveground part | 0.9 ± 0.4 | 10.4 ± 4.1 | 52.1 ± 21.9 | 77.2 ± 30.5 | 85.3 ± 32.0 | 29.6 ± 8.3 | |
| Root | 0.5 ± 0.2 | 2.6 ± 0.7 | 4.5 ± 2.0 | 3.8 ± 1.8 | 2.8 ± 1.2 | 1.4 ± 0.6 | |
| Sum | 1.4 ± 0.8 a | 13.0 ± 4.5 a | 56.6 ± 23.6 b | 81.0 ± 31.9 c | 88.1 ± 32.9 c | 92.1 ± 30.9 c | |
| Ndff (fertilizer) | Seed | 6.9 ± 2.8 | |||||
| Aboveground part | 1.3 ± 0.5 | 5.3 ± 0.9 | 7.7 ± 1.7 | 9.6 ± 2.5 | 9.9 ± 2.5 | 3.5 ± 0.5 | |
| Root | 0.6 ± 0.1 | 1.3 ± 0.3 | 0.6 ± 0.2 | 0.5 ± 0.1 | 0.4 ± 0.1 | 0.2 ± <0.1 | |
| Sum | 1.9 ± 0.6 a | 6.6 ± 1.0 bc | 8.3 ± 1.8 cd | 10.1 ± 2.6 de | 10.3 ± 2.6 de | 10.6 ± 2.9 e | |
| Ndfs (soil) | Seed | 34.0 ± 9.7 | |||||
| Aboveground part | 6.8 ± 1.8 | 18.0 ± 4.0 | 30.0 ± 8.9 | 41.4 ± 13.3 | 48.8 ± 7.4 | 17.8 ± 3.9 | |
| Root | 3.4 ± 0.6 | 4.6 ± 1.1 | 2.9 ± 1.1 | 2.4 ± 0.8 | 1.8 ± 0.6 | 1.0 ± 0.3 | |
| Sum | 10.2 ± 2.1 a | 22.6 ± 4.4 b | 32.9 ± 9.7 bc | 43.8 ± 13.9 cd | 50.6 ± 7.7 de | 52.8 ± 9.4 e | |
| Total uptake (from all sources) | Seed | 102.0 ± 32.0 | |||||
| Aboveground part | 9.0 ± 1.2 | 33.7 ± 3.9 | 89.8 ± 30.1 | 128.2 ± 42.4 | 144.0 ± 36.4 | 50.9 ± 8.9 | |
| Root | 4.5 ± 0.5 | 8.5 ± 1.7 | 8.0 ± 3.2 | 6.7 ± 2.4 | 5.0 ± 1.8 | 2.6 ± 0.9 | |
| Sum | 13.5 ± 1.1 a | 42.2 ± 4.6 b | 97.8 ± 32.9 c | 134.9 ± 44.4 d | 149.0 ± 37.8 d | 155.5 ± 37.3 d | |
| N source | Part of Plant | Source of Variation | |||
|---|---|---|---|---|---|
| Pea Cultivars | Year of Research | ||||
| ‘Milwa’ | ‘Batuta’ | 2015 | 2016 | ||
| Ndfa (air) | Seed | 62.5 ± 28.5 | 59.6 ± 18.3 | 84.1 ± 7.6 | 38.0 ± 7.1 |
| Aboveground part | 27.1 ± 8.5 | 32.2 ± 7.2 | 36.0 ± 3.6 | 23.3 ± 6.6 | |
| Root | 1.5 ± 0.8 | 1.4 ± 0.5 | 2.0 ± 0.4 | 0.8 ± 0.2 | |
| Sum | 91.1 ± 37.0 a | 93.2 ± 23.1 a | 122.1 ± 8.1 b | 62.1 ± 8.5 a | |
| Ndff (fertilizer) | Seed | 7.0 ± 3.2 | 6.8 ± 2.2 | 8.8 ± 2.7 | 5.1 ± 0.8 |
| Aboveground part | 3.3 ± 0.5 | 3.6 ± 0.5 | 3.7 ± 0.3 | 3.2 ± 0.6 | |
| Root | 0.2 ± <0.1 | 0.2 ± <0.1 | 0.2 ± <0.1 | 0.1 ± <0.1 | |
| Sum | 10.5 ± 3.3 a | 10.6 ± 2.6 a | 12.7 ± 2.6 b | 8.4 ± 0.6 a | |
| Ndfs (soil) | Seed | 34.6 ± 11.2 | 33.4 ± 7.8 | 38.3 ± 10.9 | 29.8 ± 5.3 |
| Aboveground part | 17.0 ± 4.8 | 18.7 ± 2.6 | 16.0 ± 1.5 | 19.6 ± 4.7 | |
| Root | 1.0 ± 0.3 | 0.9 ± 0.2 | 1.2 ± 0.2 | 0.7 ± 0.1 | |
| Sum | 52.6 ± 10.9 a | 53.0 ± 7.6 a | 55.5 ± 10.6 b | 50.1 ± 6.9 a | |
| Total uptake
(from all sources) | Seed | 104.1 ± 37.8 | 99.8 ± 24.7 | 131.2 ± 17.5 | 72.9 ± 5.4 |
| Aboveground part | 47.4 ± 9.2 | 54.5 ± 7.1 | 55.7 ± 3.9 | 46.1 ± 9.8 | |
| Root | 2.7 ± 1.1 | 2.5 ± 0.8 | 3.4 ± 0.7 | 1.6 ± 0.3 | |
| Sum | 154.2 ± 44.0 a | 156.8 ± 29.1 a | 190.3 ± 15.7 b | 120.6 ± 9.7 a | |
| N Source | Intervals between Growth Stages, (BBCH) | Meanly during
the Period from Sowing to Harvesting | |||||
|---|---|---|---|---|---|---|---|
| Sowing-14 | 14–33 | 33–55 | 55–65 | 65–75 | 75–90 | ||
| Ndfa (air) | 0.046 a | 0.833 b | 3.253 d | 1.895 c | 0.794 b | 0.128 a | 1.158 |
| Ndff (fertilizer) | 0.062 b | 0.333 d | 0.130 c | 0.134 c | 0.019 a | 0.011 a | 0.115 |
| Ndfs (soil) | 0.335 b | 0.882 d | 0.760 dc | 0.872 d | 0.697 c | 0.071 a | 0.603 |
| Total uptake | 0.443 a | 2.048 c | 4.143 e | 2.901 d | 1.510 b | 0.210 a | 1.876 |
| Part of Plant | N Sources | Sum | ||
|---|---|---|---|---|
| Ndfa (Air) | Ndff (Fertilizer) | Ndfs (Soil) | ||
| Crop residues | 31.0 | 3.7 | 18.8 | 53.5 |
| Seeds | 61.1 | 6.9 | 34.0 | 102.0 |
| Sum | 92.1 | 10.6 | 52.8 | 155.5 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wysokinski, A.; Lozak, I. The Dynamic of Nitrogen Uptake from Different Sources by Pea (Pisum sativum L.). Agriculture 2021, 11, 81. https://doi.org/10.3390/agriculture11010081
Wysokinski A, Lozak I. The Dynamic of Nitrogen Uptake from Different Sources by Pea (Pisum sativum L.). Agriculture. 2021; 11(1):81. https://doi.org/10.3390/agriculture11010081
Chicago/Turabian StyleWysokinski, Andrzej, and Izabela Lozak. 2021. "The Dynamic of Nitrogen Uptake from Different Sources by Pea (Pisum sativum L.)" Agriculture 11, no. 1: 81. https://doi.org/10.3390/agriculture11010081
APA StyleWysokinski, A., & Lozak, I. (2021). The Dynamic of Nitrogen Uptake from Different Sources by Pea (Pisum sativum L.). Agriculture, 11(1), 81. https://doi.org/10.3390/agriculture11010081