
Nosema Ceranae Interactions with Nosema apis and Black Queen Cell Virus




Abstract
:1. Introduction
2. Material and Methods
2.1. The Origin of Test Subjects
2.2. The Bees
2.3. Material for Infection
2.4. Experiment 1: Competition between Nosema ceranae and Nosema apis in Mixed Infection
2.4.1. The Infection Procedure
2.4.2. Nosema DNA Extraction and Quantification
2.5. Experiment 2: Influence of BQCV on the Course of Nosema Infection
2.5.1. The Infection Procedure
2.5.2. BQCV RNA Extraction and Nosema Spore Count Assessment
2.5.3. RT qPCR for BQCV Detection and Quantification
2.5.4. Statistical Analysis
3. Results
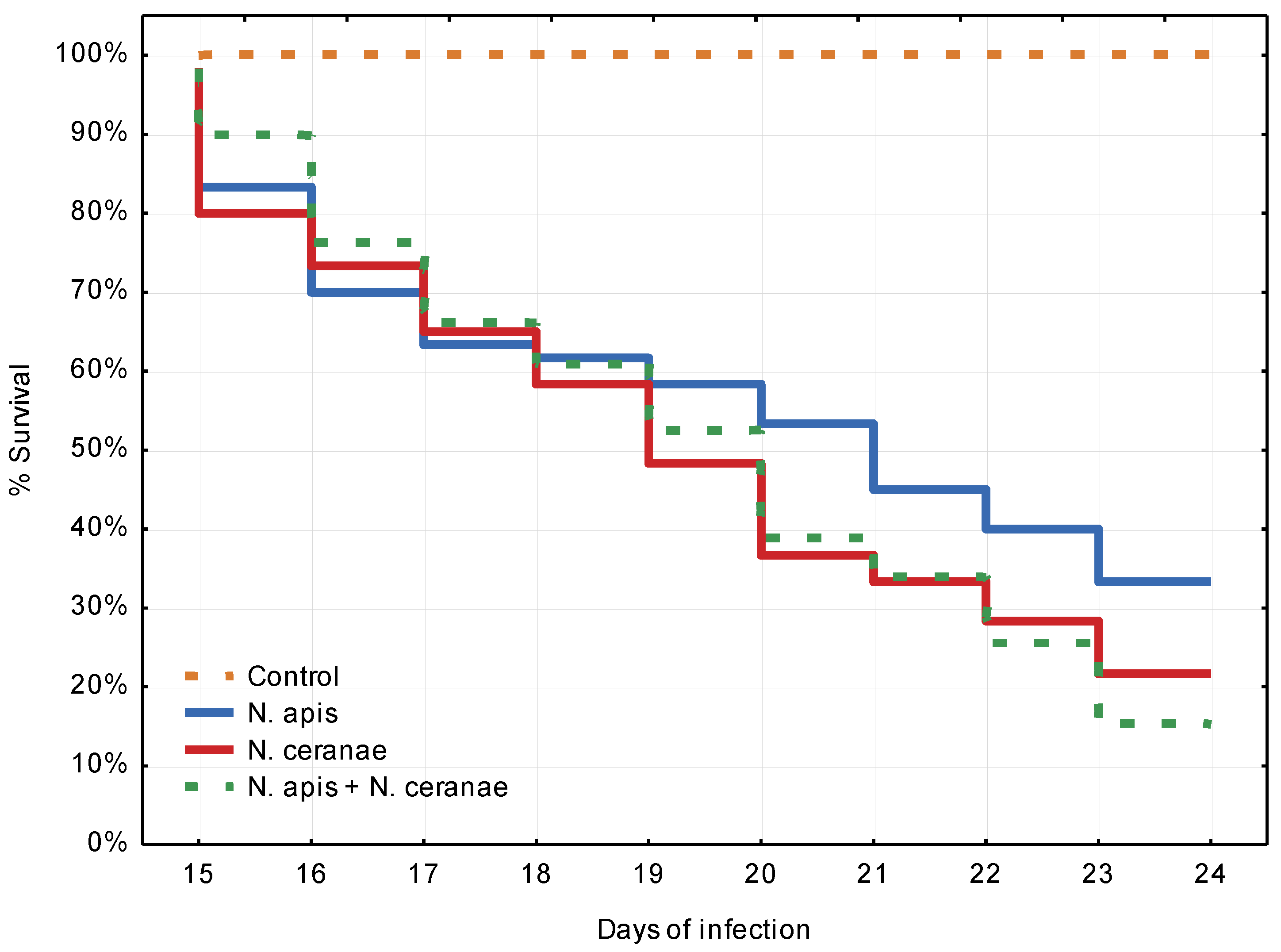
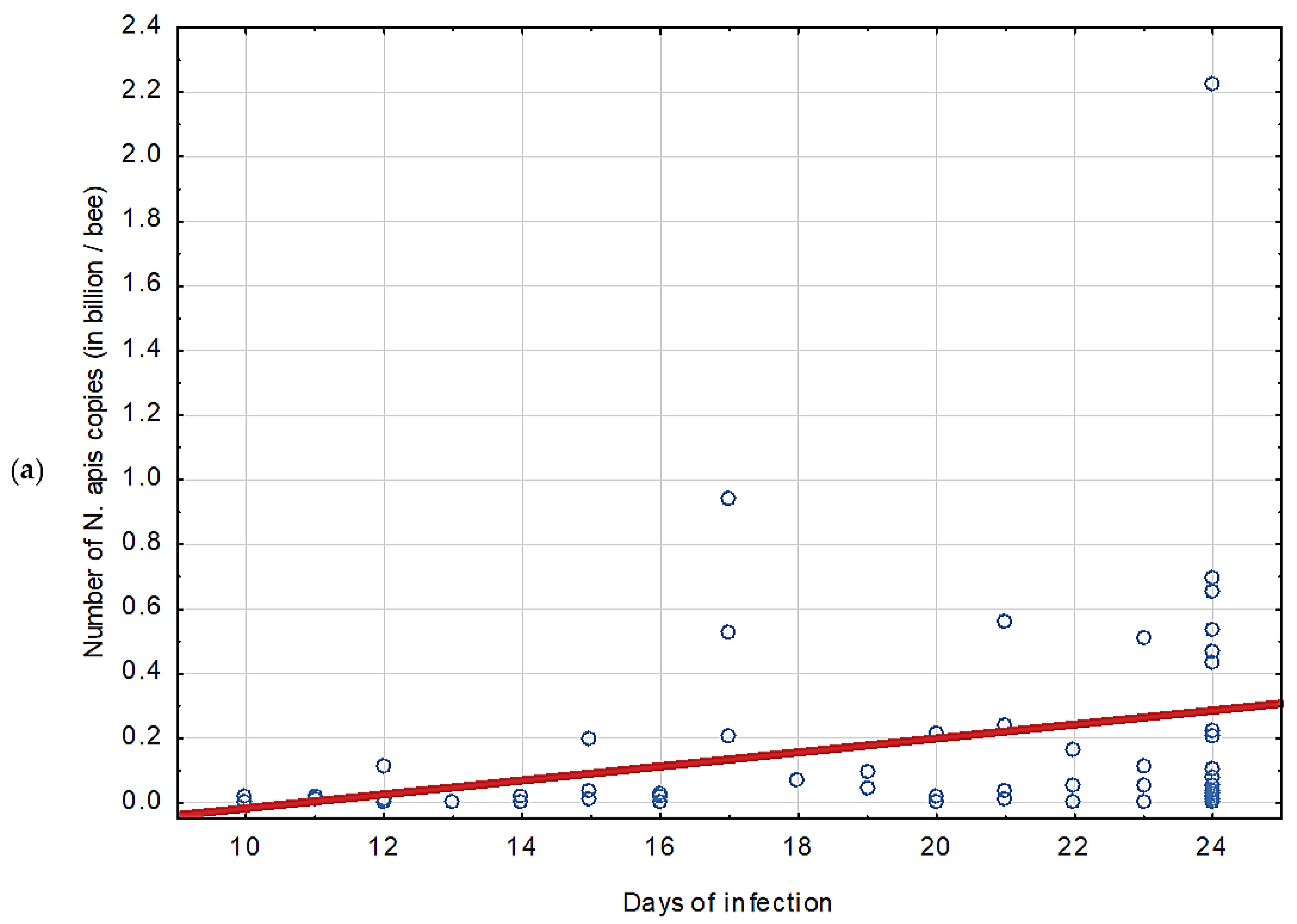
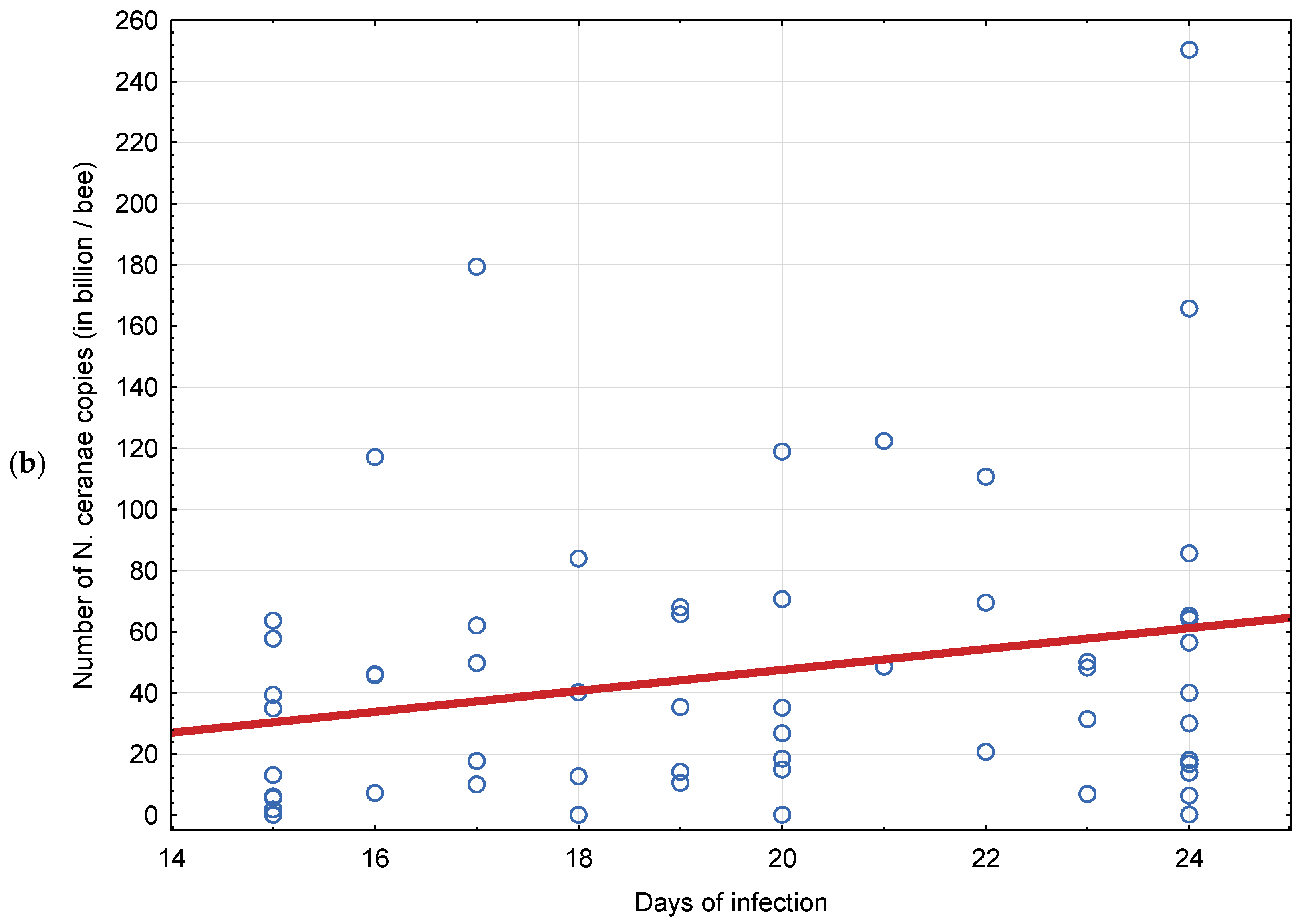
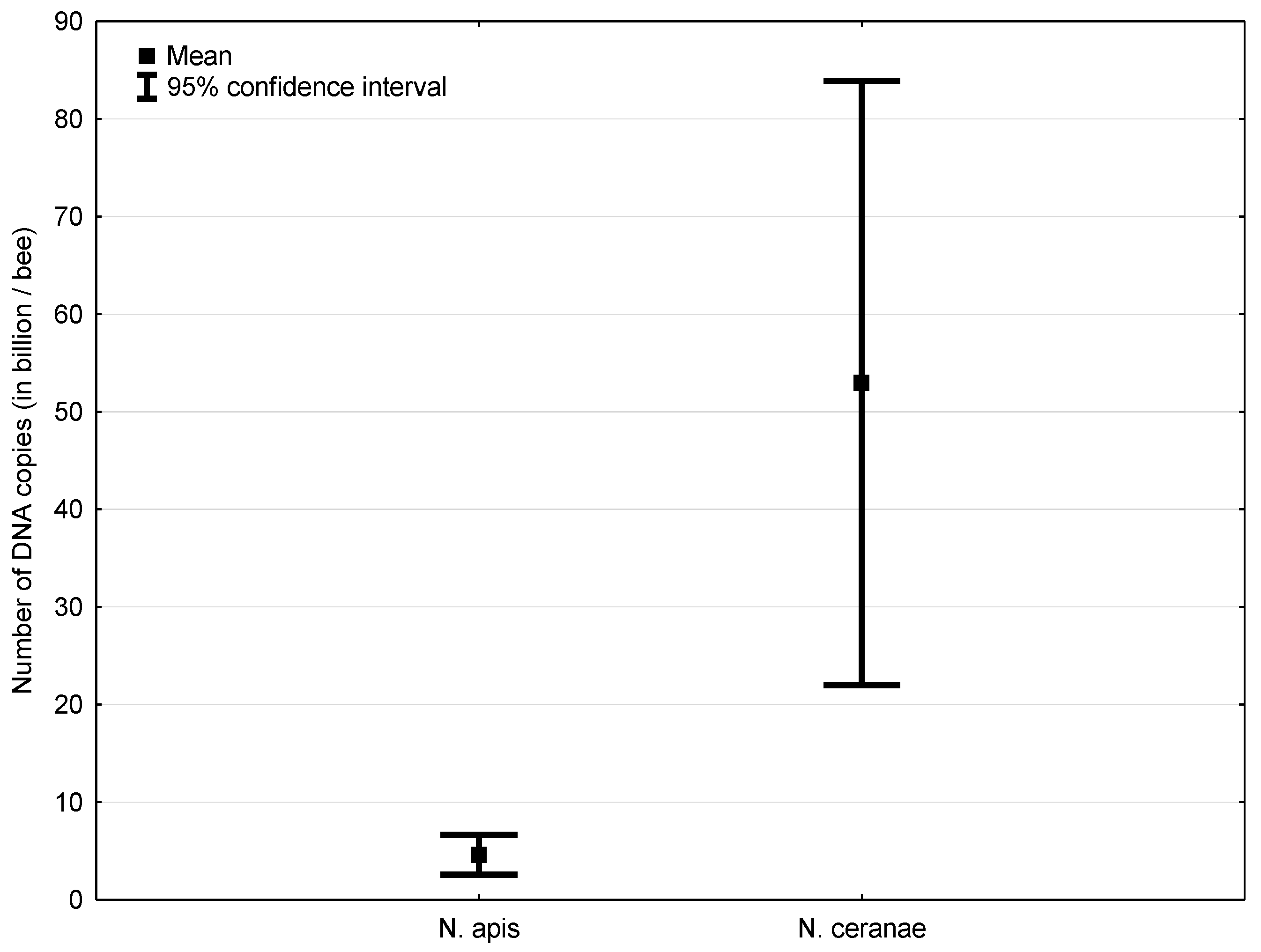
3.1. Experiment 1: Competition between Nosema ceranae and Nosema apis in Mixed Infection
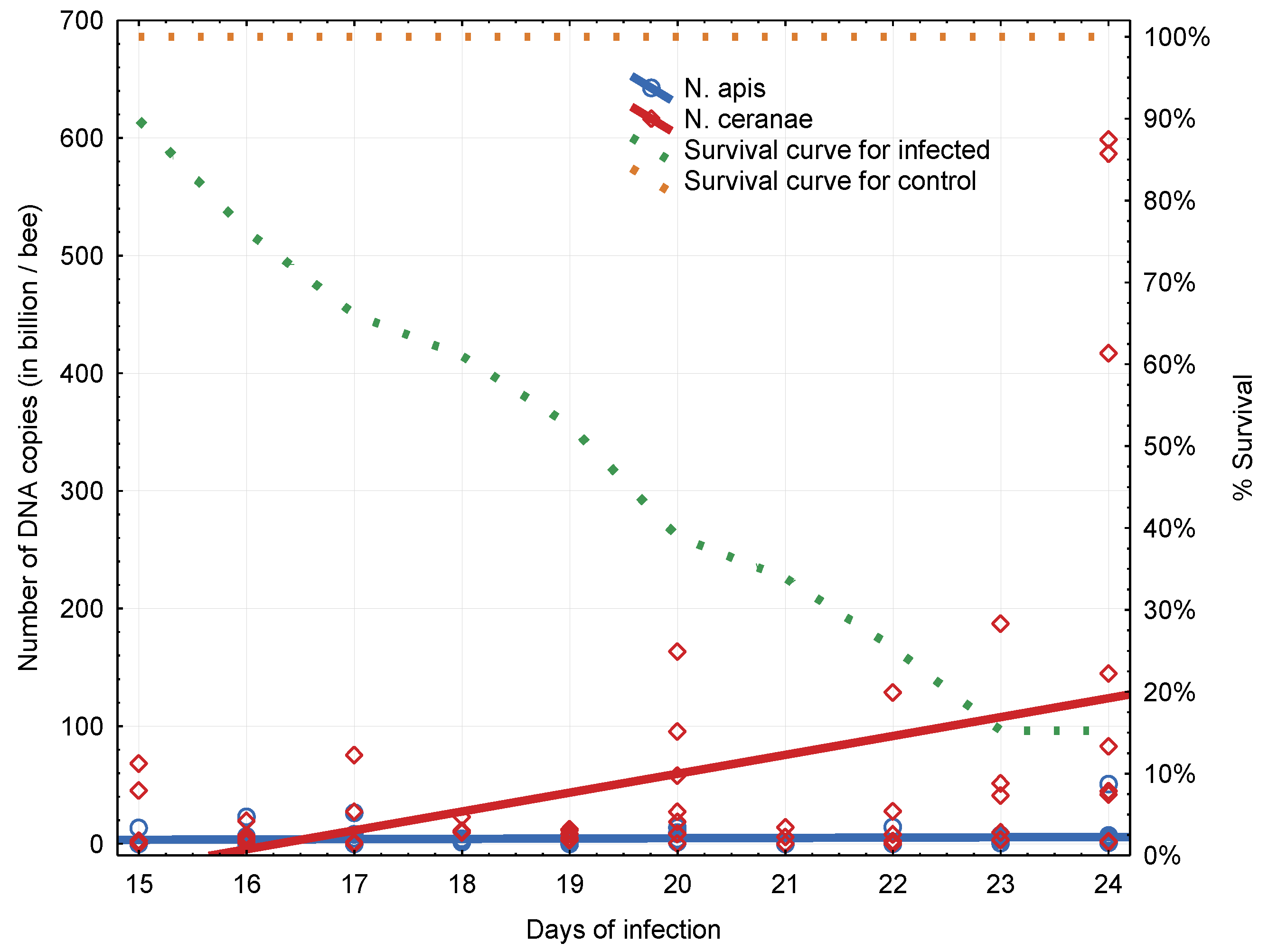
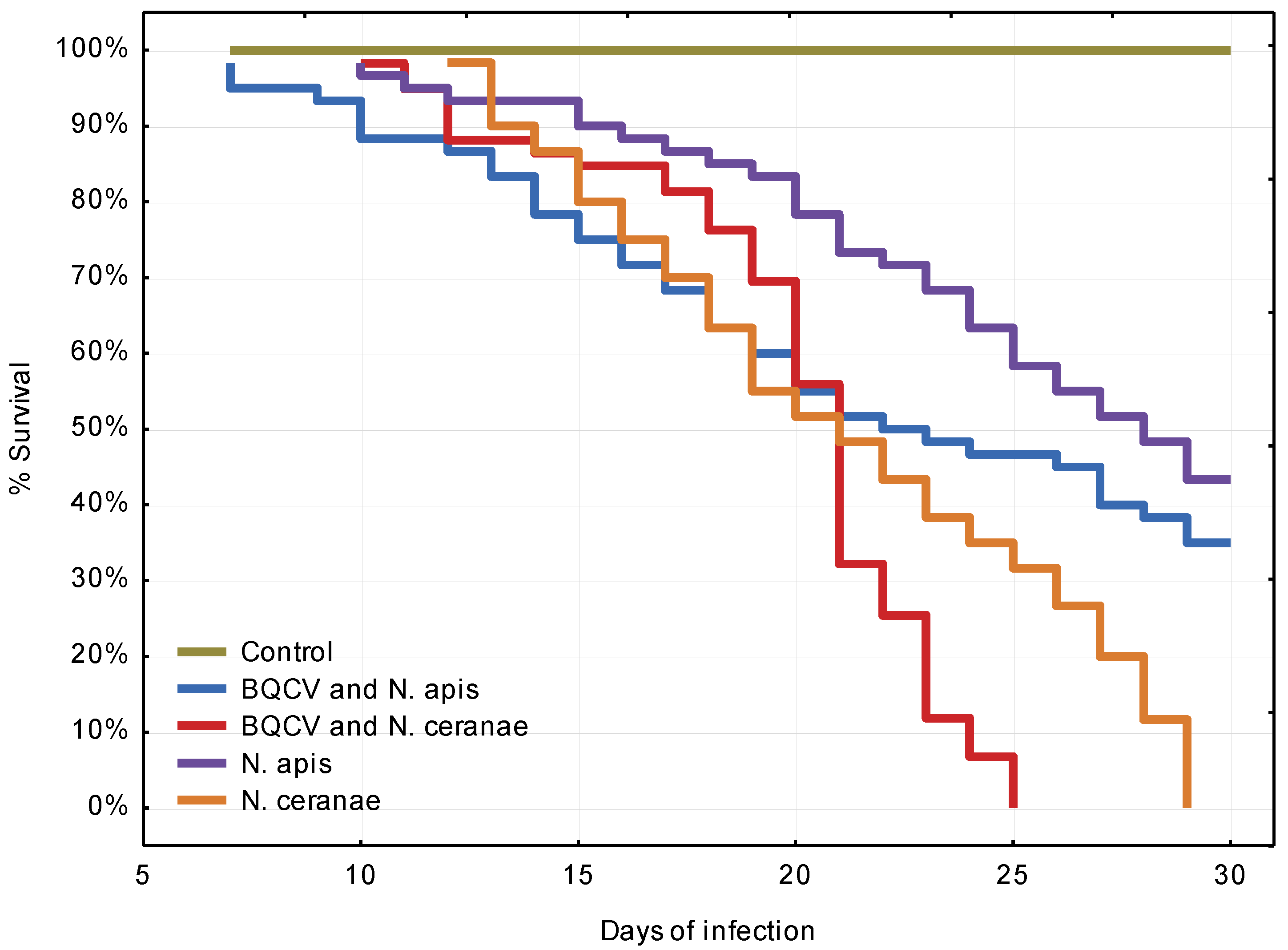
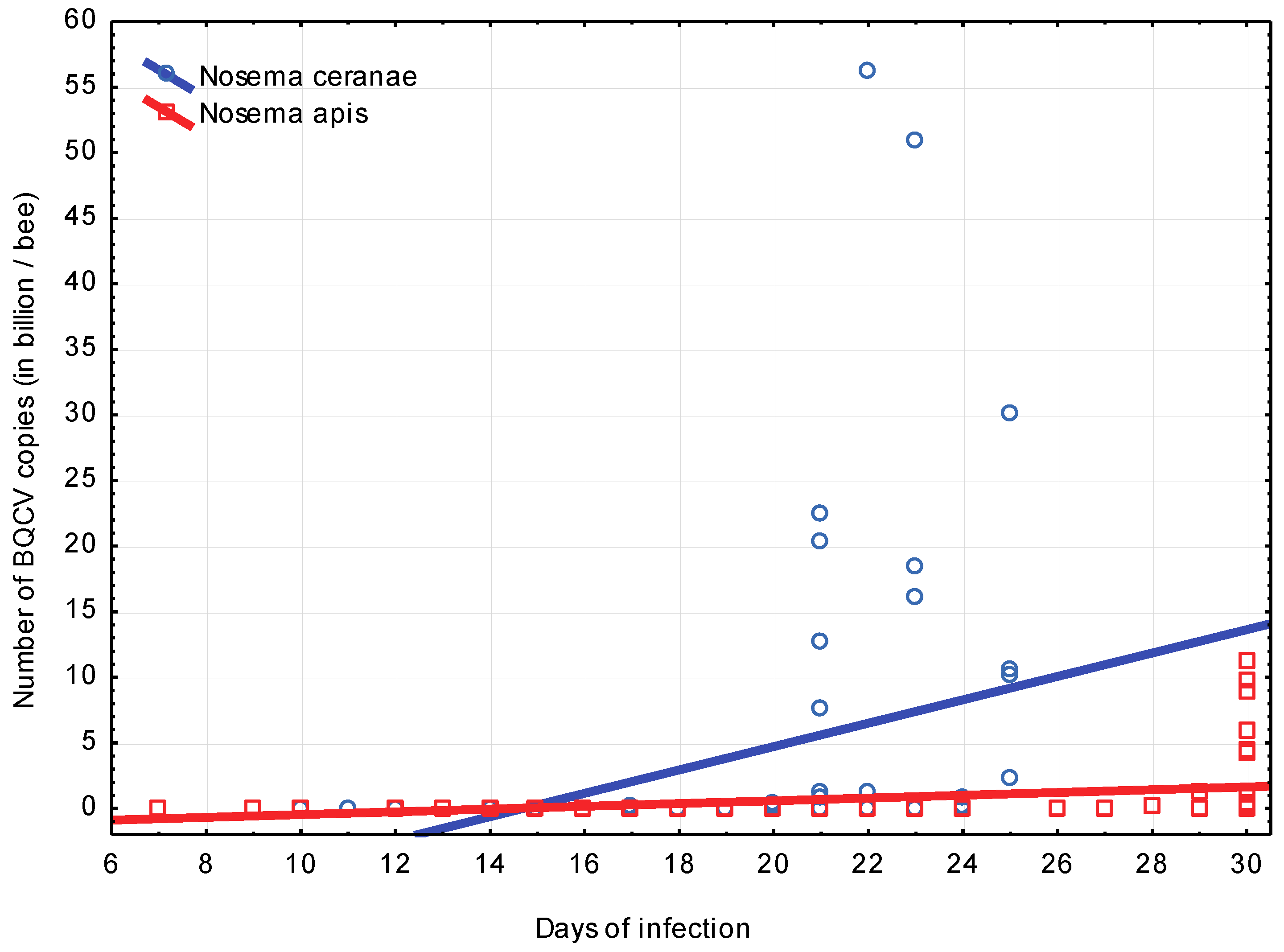
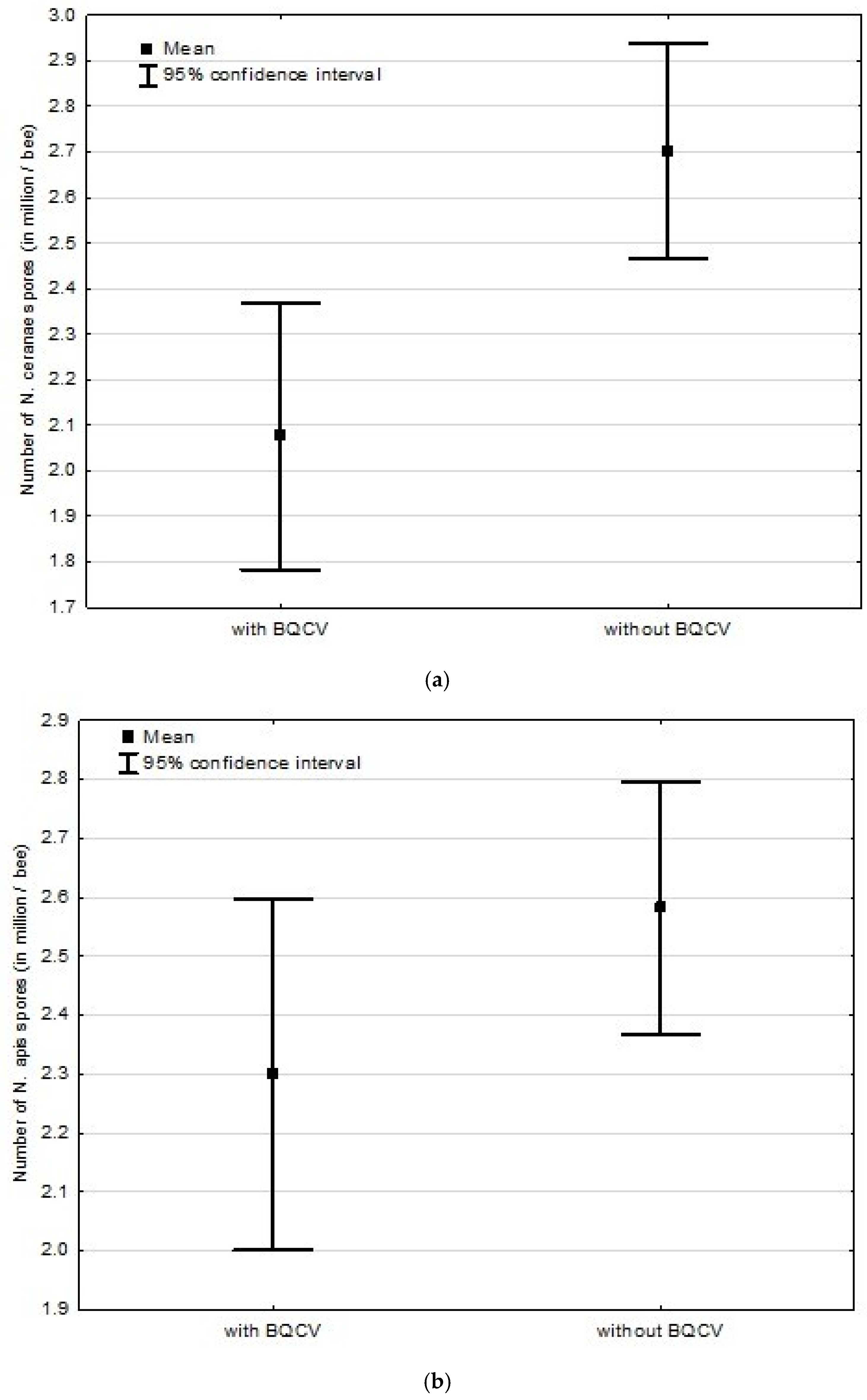
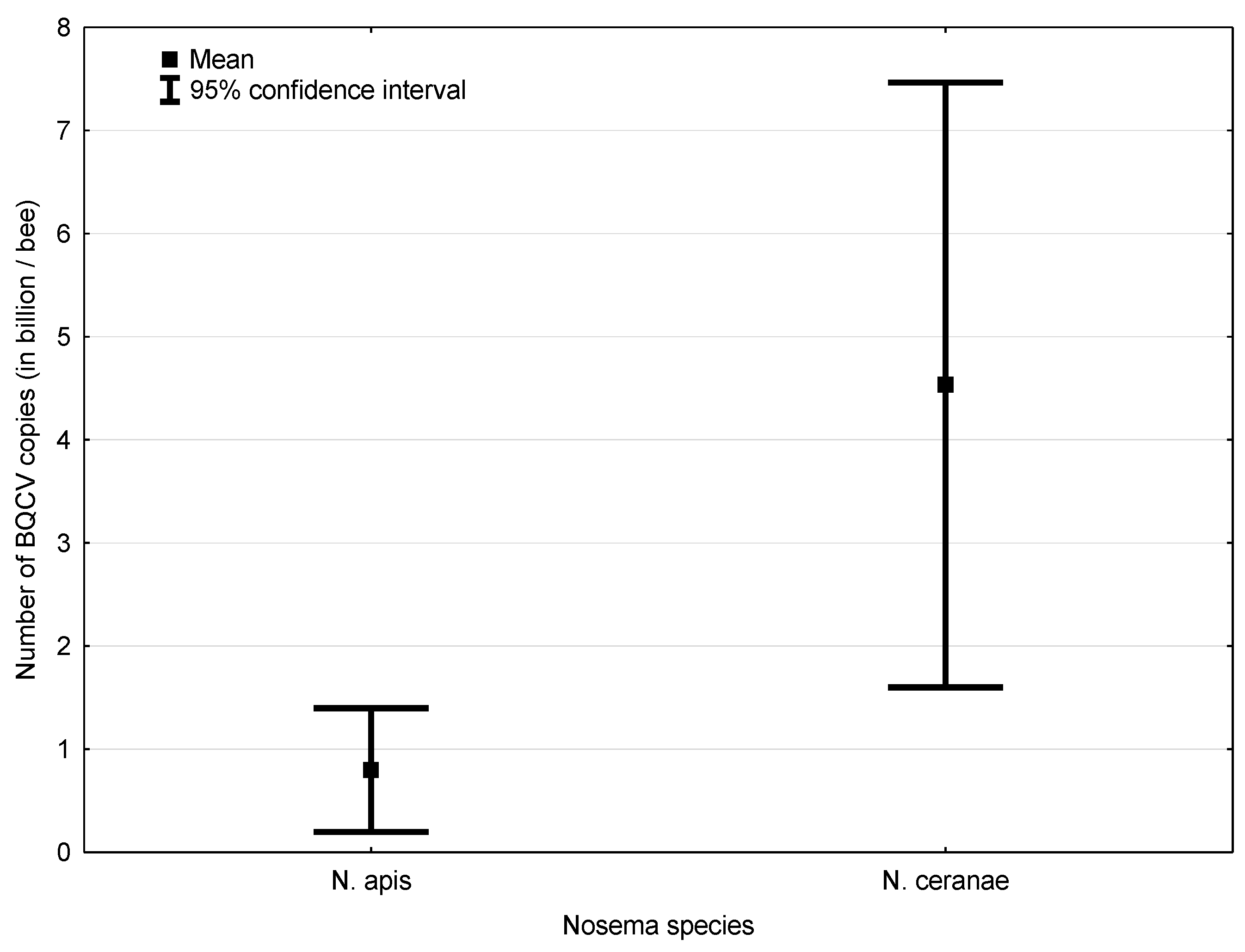
3.2. Experiment 2: Influence of BQCV on the Course of Nosema Infection
4. Discussion and Conclusions
4.1. Experiment 1: Competition between Nosema ceranae and Nosema apis in Mixed Infection
4.2. Experiment 2: Influence of BQCV on the Course of Nosema Infection
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Appendix A. Pre-Testing Colonies for the Presence of Viruses and Nosema spp.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Virus | Primer Pairs | Source |
|---|---|---|
| BQCV | BQCV76f, BQCV299r | (Bakonyi, 2002) [51] |
| ABPV | ABPV1f, ABPV2r | (Bakonyi, 2002) [51] |
| CBPV | CBPV1f, CBPV2r | (Ribiere et al., 2002) [52] |
| SBV | SBV1f, SBV2r | (Ghosh et al., 1997) [53] |
| DWV | DWV2345f, DWV2779r | (Lanzi et al., 2006) [54] |
| Primer Pairs | Annealing Temperatures and Times |
|---|---|
| BQCV76f, BQCV299r | 50 °C for 1 min |
| ABPV1f, ABPV2r | 55 °C for 1 min |
| CBPV1f, CBPV2r | 50 °C for 1 min |
| SBV1f, SBV2r | 50 °C for 1 min |
| Virus | PCR Product Size (bp) |
|---|---|
| BQCV | 224 |
| ABPV | 398 |
| CBPV | 455 |
| SBV | 646 |
| DWV | 435 |
References
- Fries, I.; Feng, F.; da Silva, A.; Slemenda, S.B.; Pieniazek, N.J. Nosema ceranae n. sp. (Microspora, Nosematidae), morphological and molecular characterization of a microsporidian parasite of the Asian honey bee Apis cerana (Hymenoptera, Apidae). Eur. J. Protistol. 1996, 32, 356–365. [Google Scholar] [CrossRef]
- Higes, M.; Martín, R.; Meana, A. Nosema ceranae, a new microsporidian parasite in honeybees in Europe. J. Invertebr. Pathol. 2006, 92, 93–95. [Google Scholar] [CrossRef]
- Klee, J.; Besana, A.M.; Genersch, E.; Gisder, S.; Nanetti, A.; Tam, D.Q.; Chinh, T.X.; Puerta, F.; Ruz, J.M.; Kryger, P.; et al. Widespread dispersal of the microsporidian Nosema ceranae, an emergent pathogen of the western honey bee, Apis mellifera. J. Invertebr. Pathol. 2007, 96, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Topolska, G.; Kasprzak, S. Pierwsze przypadki zarażenia pszczół w Polsce przez Nosema ceranae. Med. Wet. 2007, 63, 1504–1506. [Google Scholar]
- Higes, M.; Esperón, F.; Sánchez-Vizcaino, J.M. Short communication. First report of black queen-cell virus detection in honey bees (Apis mellifera) in Spain. Span. J. Agric. Res. 2007, 5, 322–325. [Google Scholar] [CrossRef]
- Vidau, C.; Panek, J.; Texier, C.; Biron, D.G.; Belzunces, L.P.; Le Gall, M.; Broussard, C.; Delbac, F.; El Alaoui, H. Differential proteomic analysis of midguts from Nosema ceranae-infected honeybees reveals manipulation of key host functions. J. Invertebr. Pathol. 2014, 121, 89–96. [Google Scholar] [CrossRef]
- Mayack, C.; Naug, D. Energetic stress in the honeybee Apis mellifera from Nosema ceranae infection. J. Invertebr. Pathol. 2009, 100, 185–188. [Google Scholar] [CrossRef]
- Gajda, A.M.; Grzęda, U.; Topolska, G.; Wilde, J.; Bieńkowska, M.; Gerula, D.; Panasiuk, B. Nosema ceranae has been present in honey bee colonies in Poland at least since 1994 and appears to have ousted Nosema apis. In Proceedings of the 9th COLOSS Conference, Kyiv, Ukraine, 27–29 September 2013; p. 38. [Google Scholar]
- Meixner, M.D.; Francis, R.M.; Gajda, A.; Kryger, P.; Andonov, S.; Uzunov, A.; Topolska, G.; Costa, C.; Amiri, E.; Berg, S.; et al. Occurrence of parasites and pathogens in honey bee colonies used in a European genotype-environment interactions experiment. J. Apic. Res. 2014, 53, 215–219. [Google Scholar] [CrossRef]
- Higes, M.; Martin-Hernández, R.; Botias, C.; Garrido-Bailón, E.; González-Porto, A.V.; Barrios, L.; Del Nozal, M.J.; Bernal, J.L.; Jiménez, J.J.; Palencia, P.G.; et al. How natural infection by Nosema ceranae causes honeybee colony collapse. Environ. Microbiol. 2008, 10, 2659–2669. [Google Scholar] [CrossRef]
- Higes, M.; Martín-Hernández, R.; Martínez-Salvador, A.; Garrido-Bailón, E.; González-Porto, A.V.; Meana, A.; Bernal, J.L.; Del Nozal, M.J.; Bernal, J. A preliminary study of the epidemiological factors related to honey bee colony loss in Spain. Environ. Microbiol. Rep. 2010, 2, 243–250. [Google Scholar] [CrossRef] [PubMed]
- Hatjina, F.; Tsoktouridis, G.; Bouga, M.; Charistos, L.; Evangelou, V.; Avtzis, D.; Meeus, I.; Brunain, M.; Smagghe, G.; de Graaf, D.C. Polar tube protein gene diversity among Nosema ceranae strains derived from a Greek honey bee health study. J. Invertebr. Pathol. 2011, 108, 131–134. [Google Scholar] [CrossRef]
- Topolska, G.; Gajda, A. Polish Honey Bee Colony-Loss During the Winter of 2007/2008. J. Apic. Sci. 2008, 52, 95–104. [Google Scholar]
- Topolska, G.; Gajda, A.; Pohorecka, K.; Bober, A.; Kasprzak, S.; Semkiw, P.; Skubida, M. Winter colony losses in Poland. J. Apic. Res. 2010, 49, 126–128. [Google Scholar] [CrossRef]
- Vercelli, M.; Novelli, S.; Ferrazzi, P.; Lentini, G.; Ferracini, C. A Qualitative Analysis of Beekeepers’ Perceptions and Farm Management Adaptations to the Impact of Climate Change on Honey Bees. Insects 2021, 12, 228. [Google Scholar] [CrossRef] [PubMed]
- Gisder, S.; Hedtke, K.; Möckel, N.; Frielitz, M.-C.; Linde, A.; Genersch, E. Five-year cohort study of Nosema spp. in Germany: Does climate shape virulence and assertiveness of Nosema ceranae? Appl. Environ. Microbiol. 2010, 76, 3032–3038. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guzman-Novoa, E.; Eccles, L.; Calvete, Y.; Mcgowan, J.; Kelly, P.G.; Correa-Benitez, A. Varroa destructor is the main culprit for the death and reduced populations of overwintered honey bee (Apis mellifera) colonies in Ontario, Canada. Apidologie 2021, 41, 443–450. [Google Scholar] [CrossRef] [Green Version]
- Williams, G.R.; Shutler, D.; Rogers, R.E.L. Effect at Nearctic north-temperate latitudes of indoor versus outdoor overwintering on the microsporidium Nosema ceranae and western honey bees (Apis mellifera). J. Invertebr. Pathol. 2010, 104, 4–7. [Google Scholar] [CrossRef]
- Invernizzi, C.; Abud, C.; Tomasco, I.H.; Harriet, J.; Ramallo, G.; Campa, J.; Katz, H.; Gardiol, G.; Mendoza, Y. Presence of Nosema ceranae in honeybees (Apis mellifera) in Uruguay. J.Invertebr. Pathol. 2009, 101, 150–153. [Google Scholar] [CrossRef]
- Milbrath, M.O.; van Tran, T.; Huang, W.-F.; Solter, L.F.; Tarpy, D.R.; Lawrence, F.; Huang, Z.Y. Comparative virulence and competition between Nosema apis and Nosema ceranae in honey bees (Apis mellifera). J. Invertebr. Pathol. 2015, 125, 9–15. [Google Scholar] [CrossRef]
- Forsgren, E.; Fries, I. Temporal study of Nosema spp. in a cold climate. Environ. Microbiol. Rep. 2013, 5, 78–82. [Google Scholar] [CrossRef]
- Bailey, L.; Ball, V.B.; Perry, J.N. Association of viruses with two protozoal pathogens of the honey bee. Ann. Appl. Biol. 1983, 103, 13–20. [Google Scholar] [CrossRef]
- Al Naggar, Y.; Paxton, R.J. Mode of Transmission Determines the Virulence of Black Queen Cell Virus in Adult Honey Bees, Posing a Future Threat to Bees and Apiculture. Viruses 2020, 12, 535. [Google Scholar] [CrossRef]
- Bailey, L.; Woods, R.D. Two more small RNA viruses from honey bees and further observations on sacbrood and acute bee-paralysis viruses. J. Gen. Virol. 1977, 37, 175–182. [Google Scholar] [CrossRef]
- Chen, Y.; Evans, J.; Feldlaufer, M. Horizontal and vertical transmission of viruses in the honey bee, Apis mellifera. J. Invertebr. Pathol. 2006, 92, 152–159. [Google Scholar] [CrossRef]
- Bailey, E. Biochemistry of Insect Flight. In Insect Biochemistry and Function; Candy, D.J., Kilby, B.A., Eds.; Chapman and Hall: London, UK, 1975. [Google Scholar] [CrossRef]
- Lodesani, M.; Costa, C.; Besana, A.; Dall’Olio, R.; Franceschetti, S.; Tesoriero, D.; Giacomo, D. Impact of control strategies for Varroa destructor on colony survival and health in northern and central regions of Italy. J. Apic. Res. 2014, 53, 155–164. [Google Scholar] [CrossRef]
- Tentcheva, D.; Gauthier, L.; Zappulla, N.; Dainat, B.; Cousserans, F.; Colin, M.E.; Bergoin, M. Prevalence and Seasonal Variations of Six Bee Viruses in Apis mellifera L. and Varroa destructor Mite Populations in France. Appl. Environ. Microbiol. 2004, 70, 7185–7191. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ravoet, J.; Maharramov, J.; Meeus, I.; De Smet, L.; Wenseleers, T.; Smagghe, G.; de Graaf, D.C. Comprehensive Bee Pathogen Screening in Belgium Reveals Crithidia mellificae as a New Contributory Factor to Winter Mortality. PLoS ONE 2013, 8, e72443. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Topolska, G.; Krzyżańska, K.; Hartwig, A.; Gajda, A. The investigation of bee virus infections in Poland. Ann. Wars. Univ. Life Sci. SGGW 2009, 46, 125–133. [Google Scholar]
- Higes, M.; García-Palencia, P.; Martín-Hernández, R.; Meana, A. Experimental infection of Apis mellifera honeybees with Nosema ceranae (Microsporidia). J. Invertebr. Pathol. 2007, 94, 211–217. [Google Scholar] [CrossRef]
- Siede, R.; Büchler, R. Symptomatic Black Queen Cell Virus infection of drone brood in Hessian apiaries. Berl. Munch. Tierarztl. Wochensch. 2003, 116, 130–133. [Google Scholar]
- Nielsen, S.L.; Nicolaisen, M.; Kryger, P. Incidence of acute bee paralysis virus, black queen cell virus, chronic bee paralysis virus, deformed wing virus, Kashmir bee virus and sacbrood virus in honey bees (Apis mellifera) in Denmark. Apidologie. 2008, 39, 310–314. [Google Scholar] [CrossRef] [Green Version]
- Nosemosis of Honey Bees. OIE Terrestrial Manual. 2018. Available online: https://www.oie.int/fileadmin/Home/eng/Health_standards/tahm/3.02.04_NOSEMOSIS_FINAL.pdf (accessed on 13 July 2021).
- Forsgren, E.; Fries, I. Comparative virulence of Nosema ceranae and Nosema apis in individual European honey bees. Vet. Parasitol. 2010, 170, 212–217. [Google Scholar] [CrossRef]
- Williams, G.R.; Shutler, D.; Burgher-MacLellan, K.L.; Rogers, R.E.L. Infra-population and -community dynamics of the parasites Nosema apis and Nosema ceranae, and consequences for honey bee (Apis mellifera) hosts. PLoS ONE 2014, 9, e99465. [Google Scholar] [CrossRef] [Green Version]
- Natsopoulou, M.E.; Doublet, V.; Paxton, R.J. European isolates of the Microsporidia Nosema apis and Nosema ceranae have similar virulence in laboratory tests on European worker honey bees. Apidologie 2016, 47, 57–65. [Google Scholar] [CrossRef] [Green Version]
- Huang, W.-F.; Solter, L.; Aronstein, K.; Huang, Z. Infectivity and virulence of Nosema ceranae and Nosema apis in commercially available North American honey bees. J. Invertebr. Pathol. 2015, 124, 107–113. [Google Scholar] [CrossRef] [PubMed]
- Paxton, R.J.; Klee, J.; Korpela, S.; Fries, I. Nosema ceranae has infected Apis mellifera in Europe since at least 1998 and may be more virulent than Nosema apis. Apidologie 2007, 38, 558–565. [Google Scholar] [CrossRef]
- Martín-Hernández, R.; Meana, A.; García-Palencia, P.; Marín, P.; Botías, C.; Garrido-Bailón, E.; Barrios, L.; Higes, M. Effect of temperature on the biotic potential of honeybee microsporidia. Appl. Environ. Microbiol. 2009, 75, 2554–2557. [Google Scholar] [CrossRef] [Green Version]
- Czekońska, K. The effect of different concentrations of carbon dioxide (CO2) in a mixture with air or nitrogen upon the survival of the honey bee (Apis mellifera). J. Apic. Res. 2009, 48, 67–71. [Google Scholar] [CrossRef]
- Tustain, R.C.R.; Faulke, J. Effect of carbon dioxide anaesthesia on the longevity of honey bees in the laboratory. N. Z. J. Exp. Agric. 1979, 7, 327–329. [Google Scholar] [CrossRef]
- Milbrath, M.O.; Xie, X.; Huang, Z.Y. Nosema ceranae induced mortality in honey bees (Apis mellifera) depends on infection methods. J. Invertebr. Pathol. 2013, 114, 42–44. [Google Scholar] [CrossRef]
- Dussaubat, C.; Sagastume, S.; Gómez-Moracho, T.; Botías, C.; García-Palencia, P.; Martín-Hernández, R.; Le Conte, Y.; Higes, M. Comparative study of Nosema ceranae (Microsporidia) isolates from two different geographic origins. Vet. Microbiol. 2013, 162, 670–678. [Google Scholar] [CrossRef]
- Huang, Q.; Kryger, P.; Le Conte, Y.; Moritz, R.F.F. Survival and immune response of drones of a Nosemosis tolerant honey bee strain towards N. ceranae infections. J. Invertebr. Pathol. 2012, 109, 297–302. [Google Scholar] [CrossRef]
- Martín-Hernández, R.; Botías, C.; Barrios, L.; Martinez-Salvador, A.; Meana, A.; Mayack, C.; Higes, M. Comparison of the energetic stress associated with experimental Nosema ceranae and Nosema apis infection of honeybees (Apis mellifera). Parasitol. Res. 2011, 109, 605–612. [Google Scholar] [CrossRef]
- Branchiccela, B.; Arredondo, D.; Higes, M.; Invernizzi, C.; Martín-Hernández, R.; Tomasco, I.; Zunino, P.; Antúnez, K. Characterization of Nosema ceranae genetic variants from different geographic origins. Microb. Ecol. 2017, 73, 978–987. [Google Scholar] [CrossRef]
- Doublet, V.; Labarussias, M.; de Miranda, J.R.; Moritz, R.F.A.; Paxton, R.J. Bees under stress: Sublethal doses of a neonicotinoid pesticide and pathogens interact to elevate honey bee mortality across the life cycle. Environ. Microbiol. 2015, 17, 969–983. [Google Scholar] [CrossRef]
- Bromenshenk, J.J.; Henderson, C.B.; Wick, C.H.; Stanford, M.F.; Zulich, A.W.; Jabbour, R.E.; Deshpande, S.V.; McCubbin, P.E.; Seccomb, R.A.; Welch, P.M.; et al. Iridovirus and microsporidian linked to honey bee colony decline. PLoS ONE. 2010, 5, e13181. [Google Scholar] [CrossRef] [PubMed]
- Virology and the Honey Bee. European Commission, Directorate-General for Research. 2008. Available online: http://edepot.wur.nl/1862 (accessed on 13 July 2021).
- Bakonyi, T. Occurrence of the Honey Bee Viruses in Hungary, Investigation of Molecular Structure of Certain Viruses. Ph.D. Thesis, Szent Istvan University, Budapest, Hungary, 2002. [Google Scholar]
- Ribiere, M.; Triboulot, C.; Mathieu, L.; Aurieres, C.; Faucon, J.-P.; Pepin, M. Molecular diagnosis of chronic bee paralysis virus infection. Apidologie 2002, 33, 339–351. [Google Scholar] [CrossRef]
- Ghosh, R.C.; Ball, B.V.; Willcocks, M.M.; Carter, M.J. The nucleotide sequence of sacbrood virus of the honey bee: An insect picorna-like virus. J. Gen. Virol. 1999, 80, 1541–1549. [Google Scholar] [CrossRef] [PubMed]
- Lanzi, G.; de Miranda, J.R.; Boniotti, M.B.; Cameron, C.E.; Lavazza, A.; Capucci, L.; Camazine, S.M.; Rossi, C. Molecular and biological characterization of deformed wing virus of honeybees (Apis mellifera L.). J. Virol. 2006, 80, 4998–5009. [Google Scholar] [CrossRef] [PubMed] [Green Version]










| Group | Spores Per Bee |
|---|---|
| N. apis | 106 |
| N. ceranae | 106 |
| N. apis + N. ceranae | 106 + 106 |
| Control | 0 (sucrose solution) |
| Nosema sp. | Spores Per Bee | Virus Copies Per Bee |
|---|---|---|
| N. apis + BQCV | 106 | 1010 |
| N. ceranae + BQCV | 106 | 1010 |
| N. apis | 106 | 0 (sucrose solution) |
| N. ceranae | 106 | 0 (sucrose solution) |
| Control | 0 (sucrose solution) | 0 (sucrose solution) |
| Group | N. apis + BQCV | N. ceranae + BQCV | N. apis | N. ceranae |
|---|---|---|---|---|
| N. apis + BCQV | XXXX | |||
| N. ceranae + BCQV | 0.08 | XXXX | ||
| N. apis | 0.04 | <0.0001 | XXXX | |
| N. ceranae | 0.15 | 0.22 | <0.0001 | XXXX |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gajda, A.M.; Mazur, E.D.; Bober, A.M.; Czopowicz, M. Nosema Ceranae Interactions with Nosema apis and Black Queen Cell Virus. Agriculture 2021, 11, 963. https://doi.org/10.3390/agriculture11100963
Gajda AM, Mazur ED, Bober AM, Czopowicz M. Nosema Ceranae Interactions with Nosema apis and Black Queen Cell Virus. Agriculture. 2021; 11(10):963. https://doi.org/10.3390/agriculture11100963
Chicago/Turabian StyleGajda, Anna Maria, Ewa Danuta Mazur, Andrzej Marcin Bober, and Michał Czopowicz. 2021. "Nosema Ceranae Interactions with Nosema apis and Black Queen Cell Virus" Agriculture 11, no. 10: 963. https://doi.org/10.3390/agriculture11100963
APA StyleGajda, A. M., Mazur, E. D., Bober, A. M., & Czopowicz, M. (2021). Nosema Ceranae Interactions with Nosema apis and Black Queen Cell Virus. Agriculture, 11(10), 963. https://doi.org/10.3390/agriculture11100963