
In Vitro Evaluation and Genome Mining of Bacillus subtilis Strain RS10 Reveals Its Biocontrol and Plant Growth-Promoting Potential



Abstract
:1. Introduction
2. Materials and Methods
2.1. Strain Isolation and Antimicrobial Activity Assay
2.2. In Vitro Plant Growth-Promoting Assay
2.3. DNA Extraction, Whole-Genome Sequencing, Assembly, and Annotation
2.4. Taxonomic Investigation and Genome Characterization of Strain RS10
2.5. Genome Mining for Synthesis of Bioactive Compounds
2.6. Pan- and Core-Genome Estimation and Phylogenetic Analysis
2.7. Availability of Data
3. Results
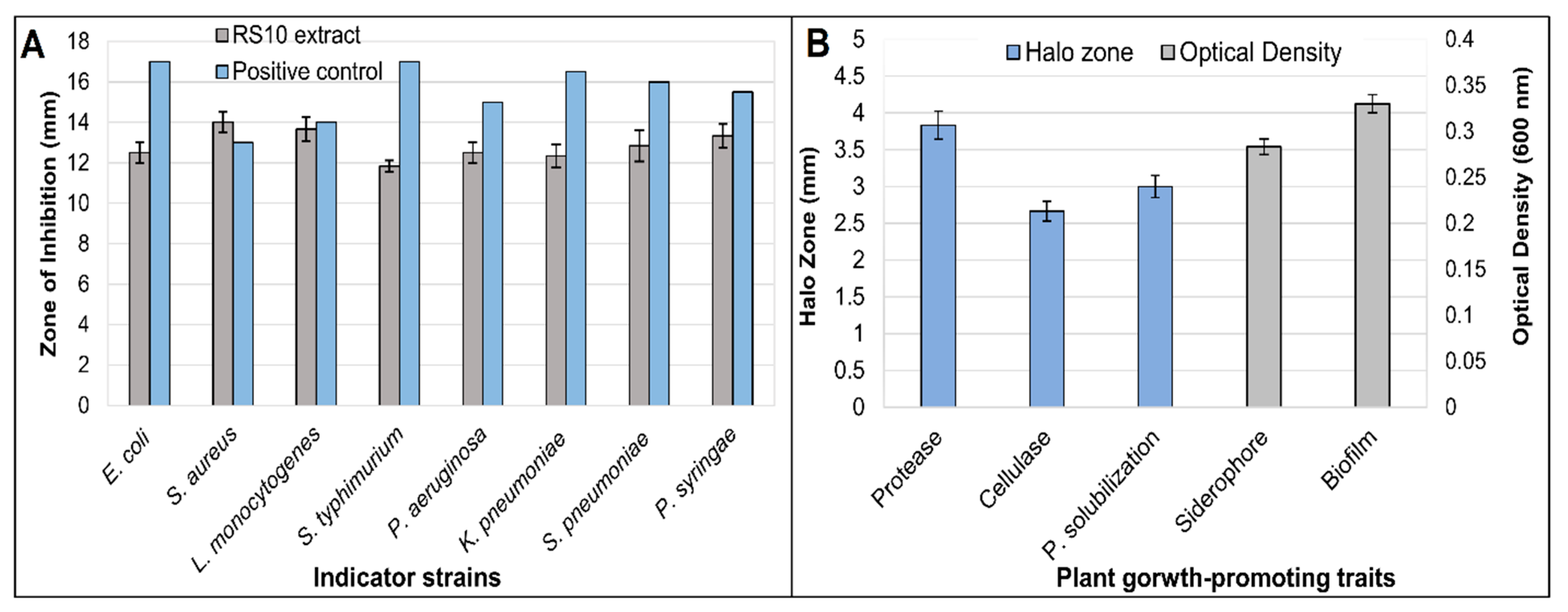
3.1. Phenotypic Characterization and Antimicrobial Activity
3.2. In Vitro Plant Growth-Promoting Traits
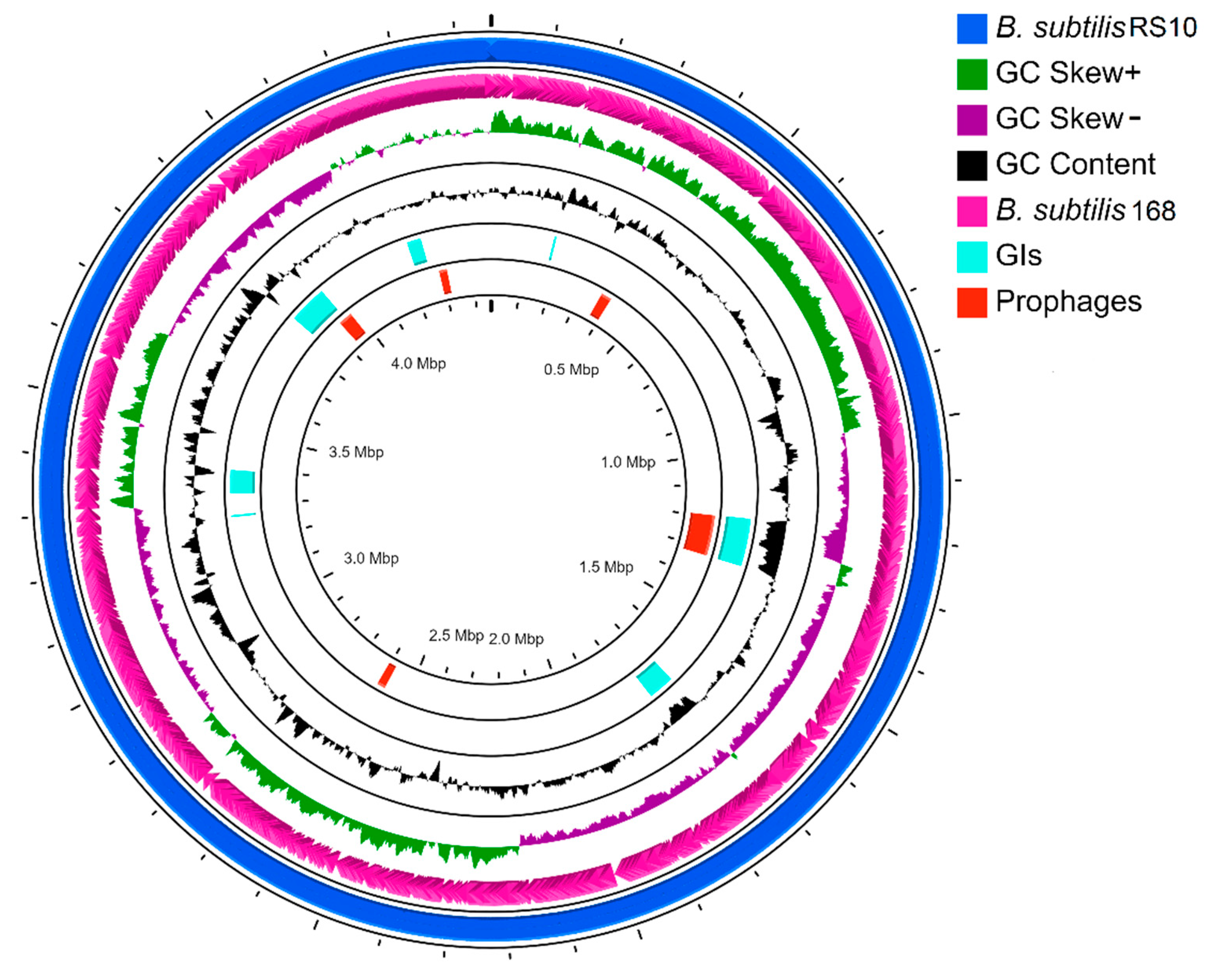
3.3. Whole-Genome Sequence and Annotations
3.4. Characterization of Genome
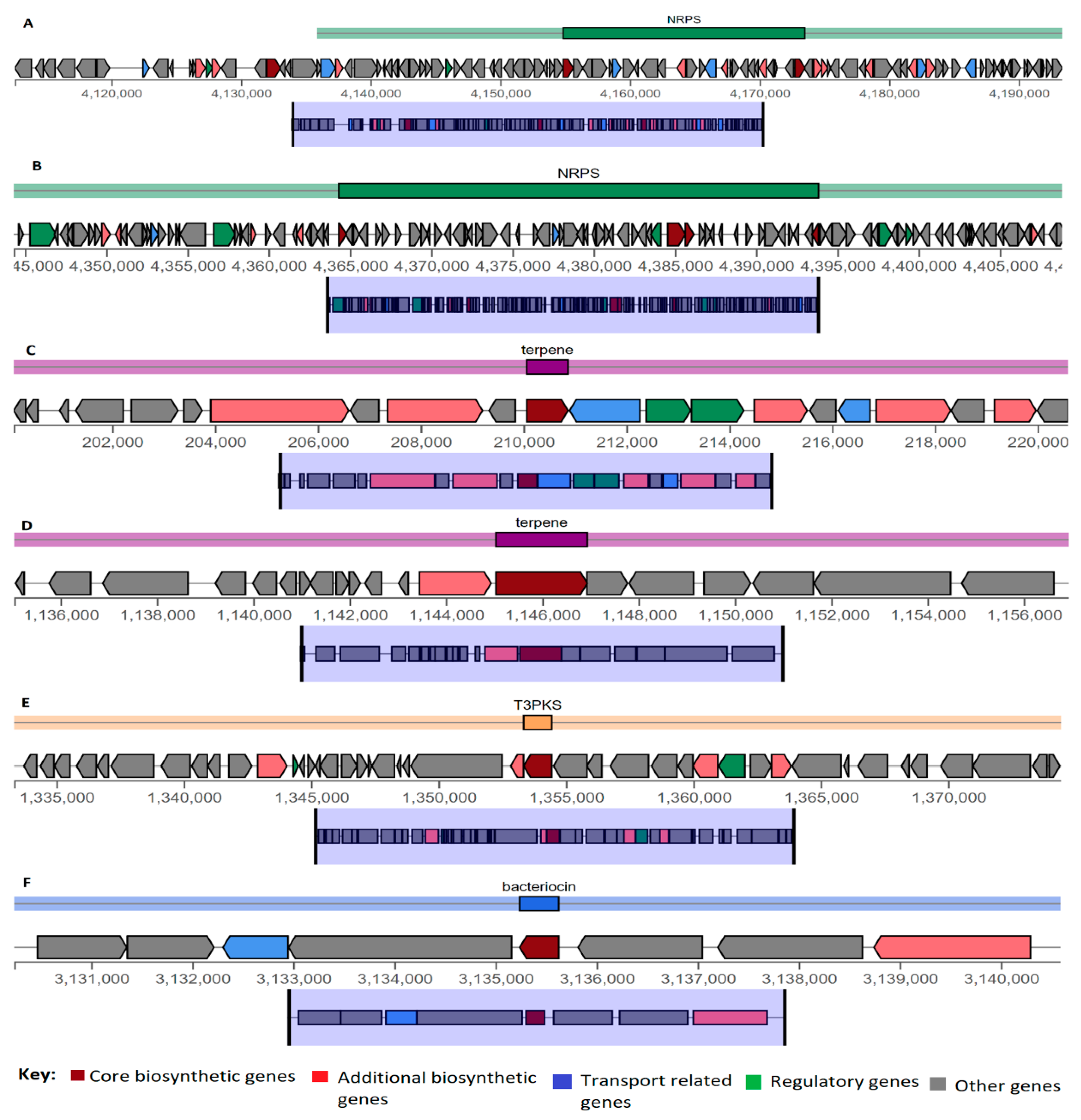
3.5. Genome Mining for Synthesis of Bioactive Compounds
3.6. Plant Growth-Promoting Capabilities
3.6.1. Nitrogen Metabolism and Phosphate Solubilization
3.6.2. Phytohormon and Siderophore Biosynthesis
3.6.3. Lytic Enzymes and Volatile Organic Compounds
3.6.4. Abiotic Stress-Related Genes
3.6.5. Chemotaxis and Colonization
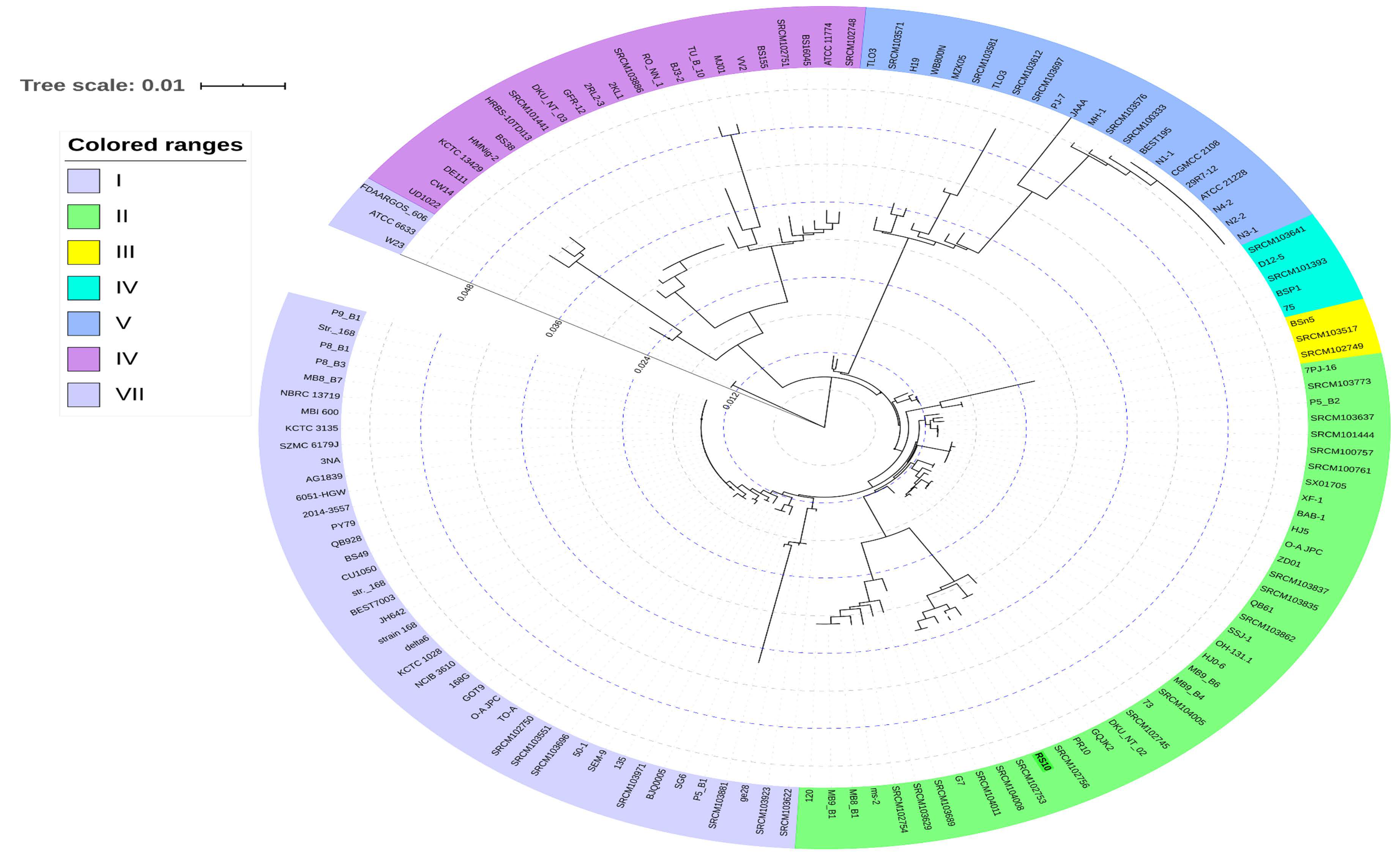
3.7. Pan- and Core-Genome Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ahemad, M.; Kibret, M. Mechanisms and applications of plant growth promoting rhizobacteria: Current perspective. J. King Saud. Univ. Sci. 2014, 26, 39–54. [Google Scholar] [CrossRef] [Green Version]
- Lugtenberg, B.; Kamilova, F. Plant-Growth-Promoting Rhizobacteria. Annu. Rev. Microbiol. 2009, 63, 541–556. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Setlow, P. Spores of Bacillus subtilis: Their resistance to and killing by radiation, heat and chemicals. J. Appl. Microbiol. 2006, 101, 514–525. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Kuipers, O.P. Identification and classification of known and putative antimicrobial compounds produced by a wide variety of Bacillales species. BMC Genom. 2016, 17, 882. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caulier, S.; Nannan, C.; Gillis, A.; Licciardi, F.; Bragard, C.; Mahillon, J. Overview of the antimicrobial compounds produced by members of the Bacillus subtilis group. Front. Microbiol. 2019, 10, 302. [Google Scholar] [CrossRef] [Green Version]
- Bais, H.P.; Fall, R.; Vivanco, J.M. Biocontrol of Bacillus subtilis against infection of Arabidopsis roots by Pseudomonas syringae is facilitated by biofilm formation and surfactin production. Plant Physiol. 2004, 134, 307–319. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Romero, D.; de Vicente, A.; Rakotoaly, R.H.; Dufour, S.E.; Veening, J.-W.; Arrebola, E.; Cazorla, F.M.; Kuipers, O.P.; Paquot, M.; Pérez-García, A. The iturin and fengycin families of lipopeptides are key factors in antagonism of Bacillus subtilis toward Podosphaera fusca. Mol. Plant Microbe Interact. 2007, 20, 430–440. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, X.H.; Scholz, R.; Borriss, M.; Junge, H.; Mögel, G.; Kunz, S.; Borriss, R. Difficidin and bacilysin produced by plant-associated Bacillus amyloliquefaciens are efficient in controlling fire blight disease. J. Biotechnol. 2009, 140, 38–44. [Google Scholar] [CrossRef] [Green Version]
- Xing, Z.; Wu, X.; Zhao, J.; Zhao, X.; Zhu, X.; Wang, Y.; Fan, H.; Chen, L.; Liu, X.; Duan, Y. Isolation and identification of induced systemic resistance determinants from Bacillus simplex Sneb545 against Heterodera glycines. Sci. Rep. 2020, 10, 11586. [Google Scholar] [CrossRef]
- Zhou, C.; Zhu, J.; Qian, N.; Guo, J.; Yan, C. Bacillus subtilis SL18r Induces Tomato Resistance Against Botrytis cinerea, Involving Activation of Long Non-coding RNA, MSTRG18363, to Decoy miR1918. Front. Plant Sci. 2021, 11, 2317. [Google Scholar] [CrossRef]
- Tahir, H.A.S.; Gu, Q.; Wu, H.; Raza, W.; Hanif, A.; Wu, L.; Colman, M.V.; Gao, X. Plant Growth Promotion by Volatile Organic Compounds Produced by Bacillus subtilis SYST2. Front. Microbiol. 2017, 8, 171. [Google Scholar] [CrossRef] [Green Version]
- Wäli, P.P.; Wäli, P.R.; Saikkonen, K.; Tuomi, J. Is the pathogenic ergot fungus a conditional defensive mutualist for its host grass? PLoS ONE 2013, 8, e69249. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schöniger, S.; Roschanski, N.; Rösler, U.; Vidovic, A.; Nowak, M.; Dietz, O.; Wittenbrink, M.M.; Schoon, H.-A. Prototheca species and Pithomyces chartarum as Causative Agents of Rhinitis and/or Sinusitis in Horses. J. Comp. Pathol. 2016, 155, 121–125. [Google Scholar] [CrossRef] [PubMed]
- Iqbal, S.; Vollmers, J.; Janjua, H.A. Genome Mining and Comparative Genome Analysis Revealed Niche-Specific Genome Expansion in Antibacterial Bacillus pumilus Strain SF-4. Genes 2021, 12, 1060–1079. [Google Scholar] [CrossRef] [PubMed]
- Iqbal, S.; Vohra, M.S.; Janjua, H.A. Whole–genome sequence and broad–spectrum antibacterial activity of Chryseobacterium cucumeris strain MW–6 isolated from the Arabian Sea. 3 Biotech 2021, 11, 489. [Google Scholar] [CrossRef]
- O’Toole, G.A. Microtiter dish biofilm formation assay. J. Vis. Exp. 2011, 2437. [Google Scholar] [CrossRef]
- Kasana, R.C.; Salwan, R.; Dhar, H.; Dutt, S.; Gulati, A. A rapid and easy method for the detection of microbial cellulases on agar plates using gram’s iodine. Curr. Microbiol. 2008, 57, 503–507. [Google Scholar] [CrossRef]
- Arora, N.K.; Verma, M. Modified microplate method for rapid and efficient estimation of siderophore produced by bacteria. 3 Biotech 2017, 7, 381. [Google Scholar] [CrossRef] [Green Version]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [Green Version]
- Bankevich, A.; Nurk, S.; Antipov, D.; Gurevich, A.A.; Dvorkin, M.; Kulikov, A.S.; Lesin, V.M.; Nikolenko, S.I.; Pham, S.; Prjibelski, A.D.; et al. SPAdes: A new genome assembly algorithm and its applications to single-cell sequencing. J. Comput. Biol. 2012, 19, 455–477. [Google Scholar] [CrossRef] [Green Version]
- Tsai, I.J.; Otto, T.D.; Berriman, M. Improving draft assemblies by iterative mapping and assembly of short reads to eliminate gaps. Genome Biol. 2010, 11, R41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tatusova, T.; Dicuccio, M.; Badretdin, A.; Chetvernin, V.; Nawrocki, E.P.; Zaslavsky, L.; Lomsadze, A.; Pruitt, K.D.; Borodovsky, M.; Ostell, J. NCBI prokaryotic genome annotation pipeline. Nucleic Acids Res. 2016, 44, 6614–6624. [Google Scholar] [CrossRef] [PubMed]
- Chaudhari, N.M.; Gupta, V.K.; Dutta, C. BPGA-an ultra-fast pan-genome analysis pipeline. Sci. Rep. 2016, 6, 24373. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]
- Bremer, E.; Krämer, R. Responses of Microorganisms to Osmotic Stress. Annu. Rev. Microbiol. 2019, 73, 313–334. [Google Scholar] [CrossRef] [PubMed]
- Glick, B.R.; Todorovic, B.; Czarny, J.; Cheng, Z.; Duan, J.; McConkey, B. Promotion of Plant Growth by Bacterial ACC Deaminase. CRC Crit. Rev. Plant Sci. 2007, 26, 227–242. [Google Scholar] [CrossRef]
- Kiesewalter, H.T.; Lozano-Andrade, C.N.; Wibowo, M.; Strube, M.L.; Maroti, G.; Snyder, D.; Jørgensen, T.S.; Larsen, T.O.; Cooper, V.S.; Weber, T.; et al. Genomic and Chemical Diversity of Bacillus subtilis Secondary Metabolites against Plant Pathogenic Fungi. mSystems 2021, 6, e00770-20. [Google Scholar] [CrossRef]
- Iqbal, S.; Rahman, H.; Begum, F.; Sajid, I.; Qasim, M. Characterization and Antibacterial Activity of Bacillus subtilis MK-4 Isolated from Southern Area of Pakistan. Int. J. Mol. Microbiol. 2019, 2, 41–50. [Google Scholar]
- Gislin, D.; Sudarsanam, D.; Raj, G.A.; Baskar, K. Antibacterial activity of soil bacteria isolated from Kochi, India and their molecular identification. J. Genet. Eng. Biotechnol. 2018, 16, 287–294. [Google Scholar] [CrossRef]
- Rahimi, T.; Niazi, A.; Deihimi, T.; Taghavi, S.M.; Ayatollahi, S.; Ebrahimie, E. Genome annotation and comparative genomic analysis of Bacillus subtilis MJ01, a new bio-degradation strain isolated from oil-contaminated soil. Funct. Integr. Genom. 2018, 18, 533–543. [Google Scholar] [CrossRef]
- Franco-Sierra, N.D.; Posada, L.F.; Santa-María, G.; Romero-Tabarez, M.; Villegas-Escobar, V.; Álvarez, J.C. Bacillus subtilis EA-CB0575 genome reveals clues for plant growth promotion and potential for sustainable agriculture. Funct. Integr. Genom. 2020, 20, 575–589. [Google Scholar] [CrossRef] [PubMed]
- Nederbragt, A.J.; Rounge, T.B.; Kausrud, K.L.; Jakobsen, K.S. Identification and Quantification of Genomic Repeats and Sample Contamination in Assemblies of 454 Pyrosequencing Reads. Sequencing 2010, 2010, 782465. [Google Scholar] [CrossRef] [Green Version]
- Lodewyckx, C.; Vangronsveld, J.; Porteous, F.; Moore, E.R.B.; Taghavi, S.; Mezgeay, M.; van der Lelie, D. Endophytic Bacteria and Their Potential Applications. CRC Crit. Rev. Plant Sci. 2002, 21, 583–606. [Google Scholar] [CrossRef]
- Stéphane, C.; Birgit, R.; Angela, S.; Jerzy, N.; Christophe, C.; Essaïd, A.B. Endophytic Colonization of Vitis vinifera L. by Plant Growth-Promoting Bacterium Burkholderia sp. Strain PsJN. Appl. Environ. Microbiol. 2005, 71, 1685–1693. [Google Scholar] [CrossRef] [Green Version]
- Rosenblueth, M.; Martínez-Romero, E. Bacterial endophytes and their interactions with hosts. Mol. Plant Microbe Interact. 2006, 19, 827–837. [Google Scholar] [CrossRef] [Green Version]
- Tariq, M.; Hameed, S.; Yasmeen, T.; Zahid, M.; Zafar, M. Molecular characterization and identification of plant growth promoting endophytic bacteria isolated from the root nodules of pea (Pisum sativum L.). World J. Microbiol. Biotechnol. 2014, 30, 719–725. [Google Scholar] [CrossRef]
- Lee, S.; Flores-Encarnación, M.; Contreras-Zentella, M.; Garcia-Flores, L.; Escamilla, J.E.; Kennedy, C. Indole-3-acetic acid biosynthesis is deficient in Gluconacetobacter diazotrophicus strains with mutations in cytochrome c biogenesis genes. J. Bacteriol. 2004, 186, 5384–5391. [Google Scholar] [CrossRef] [Green Version]
- Kavamura, V.N.; Santos, S.N.; da Silva, J.L.; Parma, M.M.; Ávila, L.A.; Visconti, A.; Zucchi, T.D.; Taketani, R.G.; Andreote, F.D.; de Melo, I.S. Screening of Brazilian cacti rhizobacteria for plant growth promotion under drought. Microbiol. Res. 2013, 168, 183–191. [Google Scholar] [CrossRef] [PubMed]
- Passari, A.K.; Mishra, V.K.; Leo, V.V.; Gupta, V.K.; Singh, B.P. Phytohormone production endowed with antagonistic potential and plant growth promoting abilities of culturable endophytic bacteria isolated from Clerodendrum colebrookianum Walp. Microbiol. Res. 2016, 193, 57–73. [Google Scholar] [CrossRef] [PubMed]
- Szilagyi-Zecchin, V.J.; Ikeda, A.C.; Hungria, M.; Adamoski, D.; Kava-Cordeiro, V.; Glienke, C.; Galli-Terasawa, L.V. Identification and characterization of endophytic bacteria from corn (Zea mays L.) roots with biotechnological potential in agriculture. AMB Express 2014, 4, 26. [Google Scholar] [CrossRef]
- Seydlová, G.; Svobodová, J. Review of surfactin chemical properties and the potential biomedical applications. Cent. Eur. J. Med. 2008, 3, 123–133. [Google Scholar] [CrossRef]
- Ahmed, E.; Holmström, S.J.M. Siderophores in environmental research: Roles and applications. Microb. Biotechnol. 2014, 7, 196–208. [Google Scholar] [CrossRef] [PubMed]
- Barsby, T.; Kelly, M.T.; Gagné, S.M.; Andersen, R.J. Bogorol A Produced in Culture by a Marine Bacillus sp. Reveals a Novel Template for Cationic Peptide Antibiotics. Org. Lett. 2001, 3, 437–440. [Google Scholar] [CrossRef] [PubMed]
- Yamada, Y.; Kuzuyama, T.; Komatsu, M.; Shin-ya, K.; Omura, S.; Cane, D.E.; Ikeda, H. Terpene synthases are widely distributed in bacteria. Proc. Natl. Acad. Sci. USA 2015, 112, 857–862. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fan, B.; Wang, C.; Song, X.; Ding, X.; Wu, L.; Wu, H.; Gao, X.; Borriss, R. Bacillus velezensis FZB42 in 2018: The Gram-Positive Model Strain for Plant Growth Promotion and Biocontrol. Front. Microbiol. 2018, 9, 2491. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Villegas-Escobar, V.; Ceballos, I.; Mira, J.J.; Argel, L.E.; Orduz Peralta, S.; Romero-Tabarez, M. Fengycin C Produced by Bacillus subtilis EA-CB0015. J. Nat. Prod. 2013, 76, 503–509. [Google Scholar] [CrossRef]
- Demtröder, L.; Narberhaus, F.; Masepohl, B. Coordinated regulation of nitrogen fixation and molybdate transport by molybdenum. Mol. Microbiol. 2019, 111, 17–30. [Google Scholar] [CrossRef] [Green Version]
- Aznar, A.; Dellagi, A. New insights into the role of siderophores as triggers of plant immunity: What can we learn from animals? J. Exp. Bot. 2015, 66, 3001–3010. [Google Scholar] [CrossRef] [Green Version]
- Willmann, R.; Nürnberger, T. How plant lysin motif receptors get activated: Lessons learned from structural biology. Sci. Signal. 2012, 5, pe28. [Google Scholar] [CrossRef] [PubMed]
- Erland, L.A.E.; Saxena, P. Auxin driven indoleamine biosynthesis and the role of tryptophan as an inductive signal in Hypericum perforatum (L.). PLoS ONE 2019, 14, e0223878. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hassan, T.; Bano, A. The stimulatory effects of L-tryptophan and plant growth promoting rhizobacteria (PGPR) on soil health and physiology of wheat. J. Soil Sci. Plant Nutr. 2015, 15, 190–201. [Google Scholar] [CrossRef] [Green Version]
- Hussain, M.; Akhtar, M.; Asghar, H.; Ahmad, M. Growth, nodulation and yield of mash bean (Vigna mungo L.) as affected by Rhizobium inoculation and soil applied L-tryptophan. Soil Environ. 2011, 30, 13–17. [Google Scholar]
- Zulqadar, S.; Arshad, M.; Naveed, M.; Hussain, A.; Nazir, Q.; Rizwan, M. Response of okra (Abelmoschus esculentus L.) to soil and foliar application of L-methionine. Soil Environ. 2015, 34, 180–186. [Google Scholar]
- Sharifi, R.; Ryu, C.-M. Revisiting bacterial volatile-mediated plant growth promotion: Lessons from the past and objectives for the future. Ann. Bot. 2018, 122, 349–358. [Google Scholar] [CrossRef] [Green Version]
- Loper, J.E.; Hassan, K.A.; Mavrodi, D.V.; Davis, E.W., II; Lim, C.K.; Shaffer, B.T.; Elbourne, L.D.H.; Stockwell, V.O.; Hartney, S.L.; Breakwell, K.; et al. Comparative Genomics of Plant-Associated Pseudomonas spp.: Insights into Diversity and Inheritance of Traits Involved in Multitrophic Interactions. PLOS Genet. 2012, 8, e1002784. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nouioui, I.; Cortés-albayay, C.; Carro, L.; Castro, J.F.; Gtari, M.; Ghodhbane-Gtari, F.; Klenk, H.-P.; Tisa, L.S.; Sangal, V.; Goodfellow, M. Genomic Insights Into Plant-Growth-Promoting Potentialities of the Genus Frankia. Front. Microbiol. 2019, 10, 1457. [Google Scholar] [CrossRef] [PubMed]
- Marín-Rodríguez, M.C.; Orchard, J.; Seymour, G.B. Pectate lyases, cell wall degradation and fruit softening. J. Exp. Bot. 2002, 53, 2115–2119. [Google Scholar] [CrossRef]
- Gao, S.; Wu, H.; Yu, X.; Qian, L.; Gao, X. Swarming motility plays the major role in migration during tomato root colonization by Bacillus subtilis SWR01. Biol. Control 2016, 98, 11–17. [Google Scholar] [CrossRef]
- Allard-Massicotte, R.; Tessier, L.; LÚcuyer, F.; Lakshmanan, V.; Lucier, J.F.; Garneau, D.; Caudwell, L.; Vlamakis, H.; Bais, H.P.; Beauregard, P.B.; et al. Bacillus subtilis Early Colonization of Arabidopsis thaliana Roots Involves Multiple Chemotaxis Receptors. MBio 2021, 7, e01664-16. [Google Scholar] [CrossRef] [Green Version]
- Gu, X.; Zeng, Q.; Wang, Y.; Li, J.; Zhao, Y.; Li, Y.; Wang, Q. Comprehensive genomic analysis of Bacillus subtilis 9407 reveals its biocontrol potential against bacterial fruit blotch. Phytopathol. Res. 2021, 3, 4. [Google Scholar] [CrossRef]
- Verstraeten, N.; Braeken, K.; Debkumari, B.; Fauvart, M.; Fransaer, J.; Vermant, J.; Michiels, J. Living on a surface: Swarming and biofilm formation. Trends Microbiol. 2008, 16, 496–506. [Google Scholar] [CrossRef] [PubMed]
- Kearns, D.B. A field guide to bacterial swarming motility. Nat. Rev. Microbiol. 2010, 8, 634–644. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lephatsi, M.M.; Meyer, V.; Piater, L.A.; Dubery, I.A.; Tugizimana, F. Plant Responses to Abiotic Stresses and Rhizobacterial Biostimulants: Metabolomics and Epigenetics Perspectives. Metabolites 2021, 11, 457. [Google Scholar] [CrossRef] [PubMed]
- Vernikos, G.; Medini, D.; Riley, D.R.; Tettelin, H. Ten years of pan-genome analyses. Curr. Opin. Microbiol. 2015, 23, 148–154. [Google Scholar] [CrossRef]
- Zhang, N.; Yang, D.; Kendall, J.R.A.; Borriss, R.; Druzhinina, I.S.; Kubicek, C.P.; Shen, Q.; Zhang, R. Comparative Genomic Analysis of Bacillus amyloliquefaciens and Bacillus subtilis Reveals Evolutional Traits for Adaptation to Plant-Associated Habitats. Front. Microbiol. 2016, 7, 2039. [Google Scholar] [CrossRef] [PubMed]
- Lopes, L.D.; Pereira, E.; de Silva, M.C.; Andreote, F.D. Bacterial Abilities and Adaptation Toward the Rhizosphere Colonization. Front. Microbiol. 2016, 7, 1341. [Google Scholar] [CrossRef] [Green Version]
- Itakura, M.; Saeki, K.; Omori, H.; Yokoyama, T.; Kaneko, T.; Tabata, S.; Ohwada, T.; Tajima, S.; Uchiumi, T.; Honnma, K.; et al. Genomic comparison of Bradyrhizobium japonicum strains with different symbiotic nitrogen-fixing capabilities and other Bradyrhizobiaceae members. ISME J. 2009, 3, 326–339. [Google Scholar] [CrossRef]
- Kim, Y.; Koh, I.; Young Lim, M.; Chung, W.-H.; Rho, M. Pan-genome analysis of Bacillus for microbiome profiling. Sci. Rep. 2017, 7, 10984. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hartmann, A.; Schmid, M.; van Tuinen, D.; Berg, G. Plant-driven selection of microbes. Plant Soil 2009, 321, 235–257. [Google Scholar] [CrossRef]
- Hao, W.; Golding, G.B. The fate of laterally transferred genes: Life in the fast lane to adaptation or death. Genome Res. 2006, 16, 636–643. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Richards, V.P.; Palmer, S.R.; Bitar, P.D.P.; Qin, X.; Weinstock, G.M.; Highlander, S.K.; Town, C.D.; Burne, R.A.; Stanhope, M.J. Phylogenomics and the dynamic genome evolution of the genus Streptococcus. Genome Biol. Evol. 2014, 6, 741–753. [Google Scholar] [CrossRef] [PubMed] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| B. subtilis Strain | Type | from–to (Location) | Similar Known Cluster | Similarity (%) | MiBIG Accession | Core Genes/Product | Locus Tag |
|---|---|---|---|---|---|---|---|
| B. subtilis strain RS10 | Terpene | 200,088–220,597 | - | - | - | farP | GPJ55_01150 |
| NRPS/PKS (Hybrid) | 811,852–926,624 | Bacillaene | 100 | BGC0001089 | pksM, pksL, pksJ, pksF, pabD | GPJ55_04475, GPJ55_04470, GPJ55_04465, GPJ55_04445, GPJ55_04435 | |
| NRPS | 991,179–1,073,561 | Fengycin | 100 | BGC0001095 | ppsA, ppsB, ppsC, ppsD, ppsE | GPJ55_05125, GPJ55_05120, GPJ55_05115, GPJ55_05110, GPJ55_05105 | |
| Terpene | 1,135,043–1,156,941 | - | - | - | sqhC | GPJ55_05630 | |
| T3PKS | 1,333,347–1,374,462 | - | - | - | Type III PK synthase | GPJ55_06995 | |
| RiPP | 2,182,762–2,204,373 | Subtilosin | 100 | BGC0000602 | albA | GPJ55_11595 | |
| NRPS | 2,732,417–2,781,432 | Bacillibactin | 100 | BGC0000309 | dhbF | GPJ55_14440 | |
| RiPP-like | 3,130,241–3,140,627 | - | - | - | Lactococcin_972 | GPJ55_16320 | |
| NRPS | 3,060,360–3,123,912 | Surfactin | 82 | BGC0000433 | srfAA, srfAB, srfAC | GPJ55_16175, GPJ55_16170, GPJ55_16165 | |
| Others | 3,621,808–3,661,798 | Bacilyicin | 100 | BGC0001184 | bacD | GPJ55_18880 | |
| NRPS | 4,134,934–4,193,216 | Bogorol A | 27 | BGC0001532 | AMP binding D | GPJ55_21740 | |
| Other | 4,292,650–4,333,087 | Zwittermicin A | 14 | BGC0001059 | ATP-grasp D | GPJ55_23355 | |
| NRPS | 4,365,207–4,413,851 | Surfactin | 21 | BGC0000433 | AMP-binding D | GPJ55_24185, GPJ55_24285 | |
| B. subtilis strain 168 | RiPP (Sectipeptide) | 204,184–226,257 | Sporulation killing factor | 100 | BGC0000601 | skfC, | HIR77_01215, HIR77_01210 |
| NRPS | 358,312–421,753 | Surfactin (Lipopeptide) | 82 | BGC0000433 | srfAA, srfAB, srfAC | HIR77_02010, HIR77_02015, HIR77_02020 | |
| NRPS/PKS (Hybride) | 1,763,795–1,878,554 | Bacillaene | 100 | BGC0001089 | fabD, pksF, pksJ, pksL, pksM | HIR77_09265, HIR77_09285, HIR77_09305, HIR77_09310, HIR77_09315 | |
| NRPS (Beta-lactone) | 1,935,481–2,017,771 | Fengycin | 100 | BGC0001095 | ppsE, ppsD, ppsC, ppsB, ppaA | HIR77_09915, HIR77_09920, HIR77_09925, HIR77_09930, HIR77_09935 | |
| RiPP (Glycocin) | 2,371,880–2,392,050 | Sublancin | 100 | BGC0000558 | sunA | HIR77_12430 | |
| RiPP (Sectipeptide) | 3,926,529–3,948,140 | Subtilosin | 100 | BGC0000602 | albA | HIR77_20830 | |
| Others | 3,951,139–3,992,557 | Bacilyicin | 100 | BGC0001184 | bacD | HIR77_21005 | |
| Terpene | 2,092,203–2,114,101 | - | - | - | sqhC | HIR77_10505 | |
| Terpene | 1,149,982–1,170,501 | - | - | - | farP | HIR77_05960 | |
| T3PKS | 2,409,316–2,450,431 | - | - | - | Type III PK synthase | HIR77_12750 | |
| NRPS | 3,373,534–3,423,275 | Bacillibactin | 100 | BGC0000309 | dhbF | HIR77_18050 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Iqbal, S.; Ullah, N.; Janjua, H.A. In Vitro Evaluation and Genome Mining of Bacillus subtilis Strain RS10 Reveals Its Biocontrol and Plant Growth-Promoting Potential. Agriculture 2021, 11, 1273. https://doi.org/10.3390/agriculture11121273
Iqbal S, Ullah N, Janjua HA. In Vitro Evaluation and Genome Mining of Bacillus subtilis Strain RS10 Reveals Its Biocontrol and Plant Growth-Promoting Potential. Agriculture. 2021; 11(12):1273. https://doi.org/10.3390/agriculture11121273
Chicago/Turabian StyleIqbal, Sajid, Nimat Ullah, and Hussnain Ahmed Janjua. 2021. "In Vitro Evaluation and Genome Mining of Bacillus subtilis Strain RS10 Reveals Its Biocontrol and Plant Growth-Promoting Potential" Agriculture 11, no. 12: 1273. https://doi.org/10.3390/agriculture11121273
APA StyleIqbal, S., Ullah, N., & Janjua, H. A. (2021). In Vitro Evaluation and Genome Mining of Bacillus subtilis Strain RS10 Reveals Its Biocontrol and Plant Growth-Promoting Potential. Agriculture, 11(12), 1273. https://doi.org/10.3390/agriculture11121273