
Field Inoculation of Arbuscular Mycorrhizal Fungi Improves Fruit Quality and Root Physiological Activity of Citrus

 , ,
, ,  and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Setup
2.2. Determinations of Variables
2.3. Statistical Analysis
3. Results
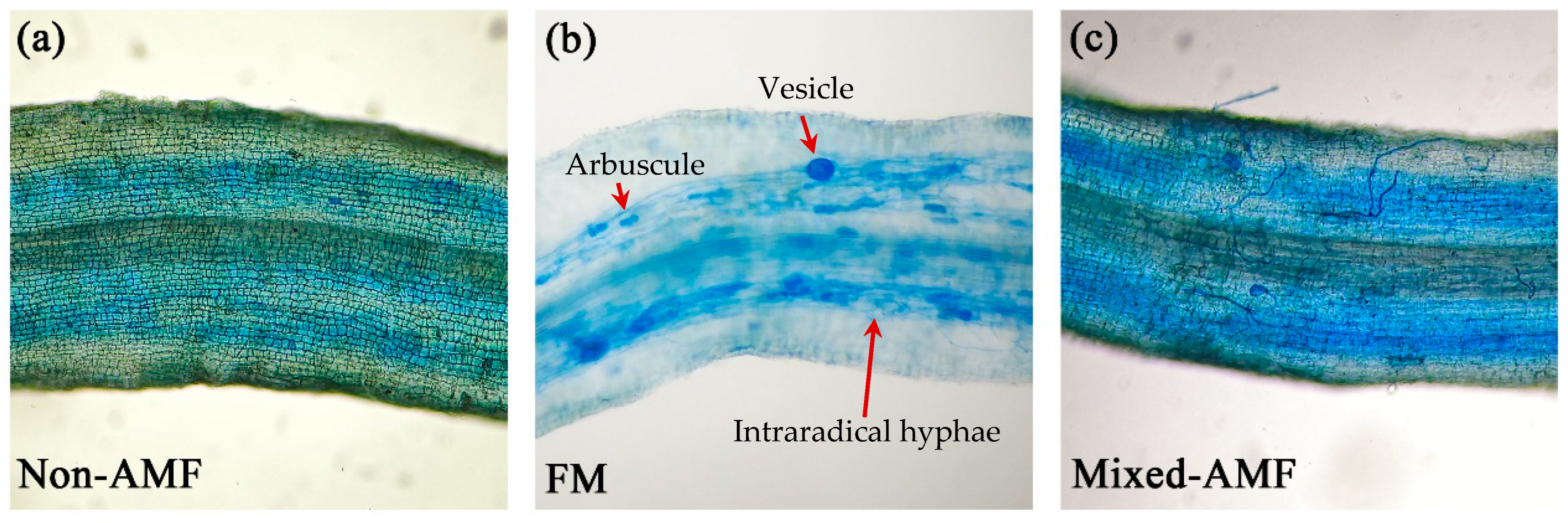
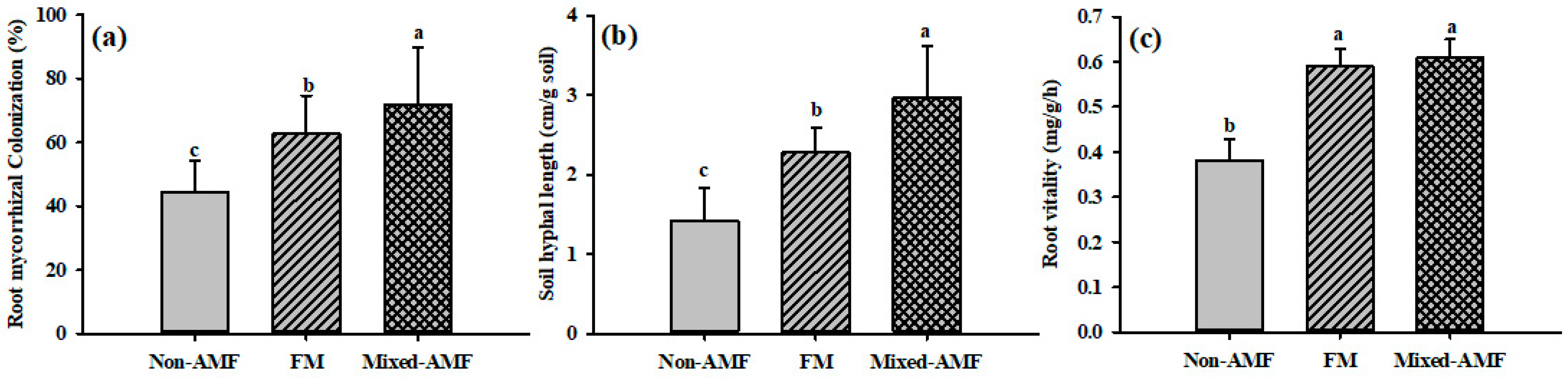
3.1. Changes in Degree of Root Mycorrhizal Colonization, Soil Hyphal Length, and Root Vitality
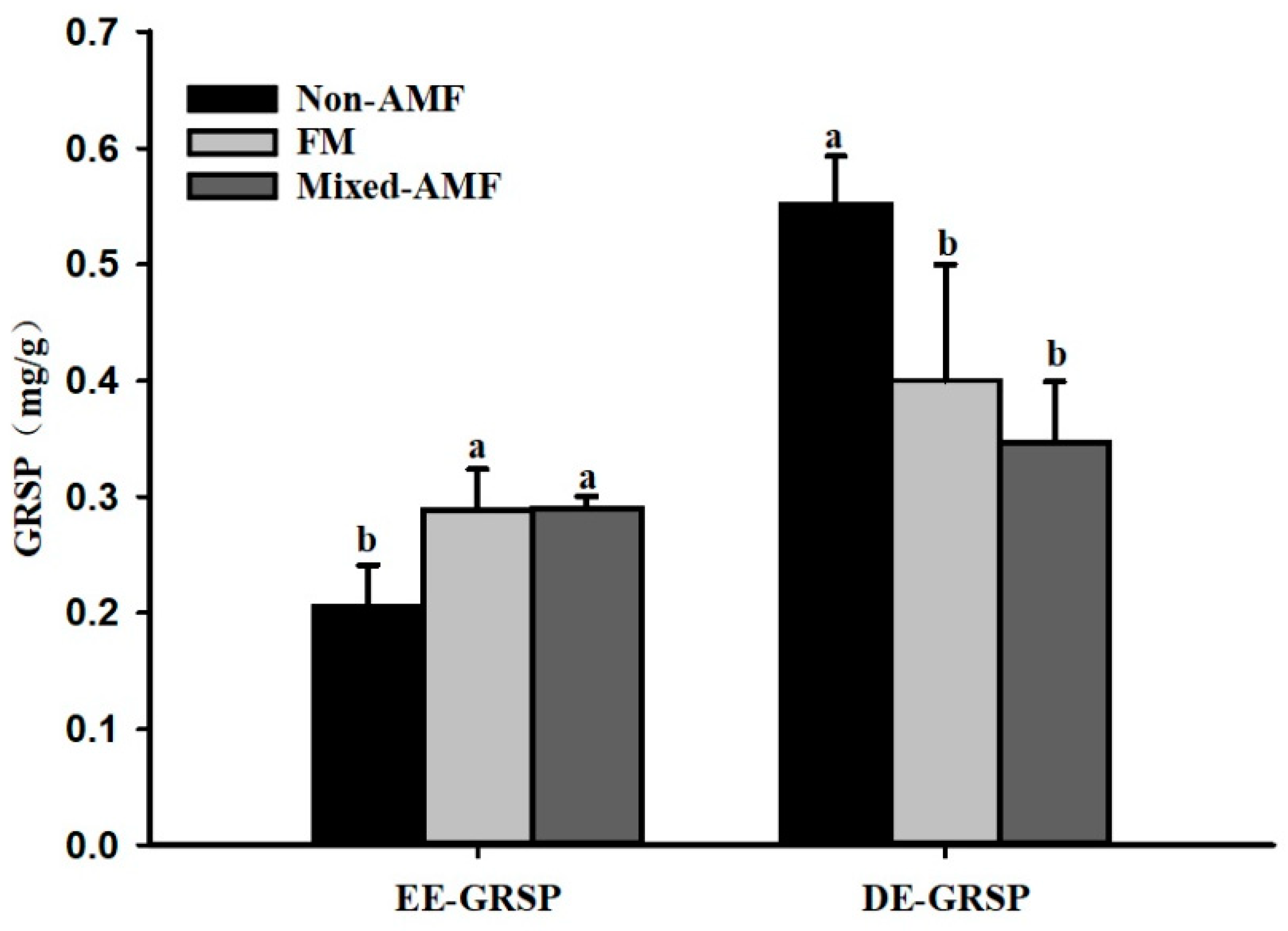
3.2. Changes in Soil GRSP Levels
3.3. Changes in Internal and External Quality Parameters of Fruits
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zou, Y.N.; Wu, Q.S.; Kuča, K. Unravelling the role of arbuscular mycorrhizal fungi in mitigating the oxidative burst of plants under drought stress. Plant Biol 2021, 23 (Suppl. 1), 50–57. [Google Scholar] [CrossRef]
- Agnolucci, M.; Avio, L.; Pepe, A.; Turrini, A.; Cristani, C.; Bonini, P.; Cirino, V.; Colosimo, F.; Ruzzi, M.; Giovannetti, M. Bacteria associated with a commercial mycorrhizal inoculum: Community composition and multifunctional activity as assessed by Illumina sequencing and culture-dependent tools. Front. Plant Sci. 2019, 9, 1956. [Google Scholar] [CrossRef]
- Wright, S.F.; Franke-Snyder, M.; Morton, J.B.; Morton, A.; Upadhyaya, A. Time-course study and partial characterization of a protein on hyphae of arbuscular mycorrhizal fungi during active colonization of roots. Plant Soil 1996, 181, 193–203. [Google Scholar] [CrossRef]
- He, S.B.; Long, M.X.; He, X.Q.; Guo, L.X.; Yang, J.Y.; Yang, P.Z.; Hu, T.M. Arbuscular mycorrhizal fungi and water availability affect biomass and C: N: P ecological stoichiometry in alfalfa (Medicago sativa L.) during regrowth. Acta Physiol. Plant. 2017, 39, 199–208. [Google Scholar] [CrossRef]
- Wright, S.F.; Upadhyaya, A. A survey of soils for aggregate stability and glomalin a glycoprotein of arbuscular mycorrhizal fungi. Plant Soil 1998, 198, 97–107. [Google Scholar] [CrossRef]
- He, J.D.; Chi, G.G.; Zou, Y.N.; Shu, B.; Wu, Q.S.; Srivastava, A.K.; Kuča, K. Contribution of glomalin-related soil proteins to soil organic carbon in trifoliate orange. Appl. Soil Ecol. 2020, 154, 103592. [Google Scholar] [CrossRef]
- Meng, L.L.; He, J.D.; Zou, Y.N.; Wu, Q.S.; Kuča, K. Mycorrhiza-released glomalin-related soil protein fractions contribute to soil total nitrogen in trifoliate orange. Plant Soil Environ. 2020, 66, 183–189. [Google Scholar] [CrossRef]
- Ali, A.; Ghani, M.I.; Ding, H.Y.; Fan, Y.; Cheng, Z.H.; Iqbal, M. Co-amended synergistic interactions between arbuscular mycorrhizal fungi and the organic substrate-induced cucumber yield and fruit quality associated with the regulation of the AM-fungal community structure under anthropogenic cultivated soil. Int. J. Mol. Sci. 2019, 20, 1539. [Google Scholar] [CrossRef] [Green Version]
- Wu, Q.S.; Zou, Y.N.; Liu, C.Y.; Lu, T. Interacted effect of arbuscular mycorrhizal fungi and polyamines on root system architecture of citrus seedlings. J. Integr. Agric. 2012, 11, 1675–1681. [Google Scholar] [CrossRef]
- Song, F.; Bai, F.X.; Wang, J.J.; Wu, L.M.; Jiang, Y.C.; Pan, Z.Y. Influence of citrus scion/rootstock genotypes on arbuscular mycorrhizal community composition under controlled environment condition. Plants 2020, 9, 901. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.; Wang, T.Y.; Wu, S.H.; Weng, M.X.; Lu, L.M.; Ke, F.Z.; Wu, Q.S. Effect of arbuscular mycorrhizal fungi on rhizosphere organic acid content and microbial activity of trifoliate orange under different low P conditions. Arch. Agron. Soil Sci. 2019, 65, 2029–2042. [Google Scholar] [CrossRef]
- Liang, S.M.; Zhang, F.; Zou, Y.N.; Kucča, K.; Wu, Q.S. Metabolomics analysis reveals drought responses of trifoliate orange by arbuscular mycorrhizal fungi with a focus on terpenoid profile. Front. Plant Sci. 2021, 12, 740524. [Google Scholar] [CrossRef]
- Cheng, H.Q.; Zou, Y.N.; Wu, Q.S.; Kuča, K. Arbuscular mycorrhizal fungi alleviate drought stress in trifoliate orange by regulating H+-ATPase activity and gene expression. Front. Plant Sci. 2021, 12, 659694. [Google Scholar] [CrossRef] [PubMed]
- Sabra, M.; Aboulnasr, A.; Franken, P.; Perreca, E.; Wright, L.P.; Camehl, I. Beneficial root endophytic fungi increase growth and quality parameters of sweet basil in heavy metal contaminated soil. Front. Plant Sci. 2018, 27, 1726. [Google Scholar] [CrossRef] [Green Version]
- Lin, A.J.; Zhang, X.H.; Wong, M.H.; Ye, Z.H.; Lou, L.Q.; Wang, Y.S.; Zhu, Y.G. Increase of multi-metal tolerance of three leguminous plants by arbuscular mycorrhizal fungi colonization. Environ. Geochem. Health 2007, 29, 473–481. [Google Scholar] [CrossRef]
- Regvar, M.; Vogel-Miku, K.; Everkar, T. Effect of AMF inoculum from field isolates on the yield of green pepper, parsley, carrot, and tomato. Folia Geobot. 2003, 38, 223–234. [Google Scholar] [CrossRef]
- Ercol, L.; Arthur, S.; Iduna, A.; Elisa, P. Strong increase of durum wheat iron and zinc content by field-inoculation with arbuscular mycorrhizal fungi at different soil nitrogen availabilities. Plant Soil 2017, 419, 153–167. [Google Scholar] [CrossRef]
- Marie-Stéphanie Kouadio, A.N.; Nandjui, J.; Krou, S.M.; Jean-Marc Séry, D.; Nelson, P.N.; Adolphe, Z. A native arbuscular mycorrhizal fungus inoculant outcompetes an exotic commercial species under two contrasting yam field conditions. Rhizosphere 2017, 4, 112–118. [Google Scholar] [CrossRef]
- Paskovic, I.; Soldo, B.; Ban, S.G.; Radic, T.; Lukic, M.; Urlic, B.; Mimica, M.; Bubola, K.B.; Colla, G.; Rouphael, Y.; et al. Fruit quality and volatile compound composition of processing tomato as affected by fertilisation practices and arbuscular mycorrhizal fungi application. Food Chem. 2021, 359, 129961. [Google Scholar] [CrossRef]
- Ortas, I. Mycorrhiza in citrus: Growth and nutrition. In Advances in Citrus Nutrition; Srivastava, A.K., Ed.; Springer: New York, NY, USA, 2012; pp. 333–352. [Google Scholar]
- Phillips, J.M. Improved procedures for clearing roots and staining parasitic and vesicular-arbuscular mycorrhizal fungi for rapid assessment of infection. Trans. Br. Mycol. Soc. 1970, 55, 158–161. [Google Scholar] [CrossRef]
- Bethlenfalvay, G.J.; Ames, R.N. Comparison of two methods for quantifying extraradical mycelium of vesicular-arbuscular mycorrhizal fungi. Soil Sci. Soc. Am. J. 1987, 51, 834–837. [Google Scholar] [CrossRef]
- Lindström, A.; Nyström, C. Seasonal variation in root hardiness of container-grown Scots pine, Norway spruce, and lodgepole pine seedlings. Can. J. For. Res. 1987, 17, 787–793. [Google Scholar] [CrossRef]
- Wu, Q.S.; Cao, M.Q.; Zou, Y.N.; He, X.H. Direct and indirect effects of glomalin, mycorrhizal hyphae, and roots on aggregate stability in rhizosphere of trifoliate orange. Sci. Rep. 2014, 4, 5823. [Google Scholar] [CrossRef] [PubMed]
- Valverde-Barrantes, O.J.; Smemo, K.A.; Feinstein, L.M.; Kershner, M.W.; Blackwood, C.B. Patterns in spatial distribution and root trait syndromes for ecto and arbuscular mycorrhizal temperate trees in a mixed broadleaf forest. Oecologia 2018, 186, 731–741. [Google Scholar] [CrossRef]
- Lovelock, C.E.; Wright, S.F.; Ruess, C. Soil stocks of glomalin produced by arbuscular mycorrhizal fungi across a tropical rain forest landscape. J. Ecol. 2004, 92, 278–287. [Google Scholar] [CrossRef]
- Liu, R.C.; Gao, W.Q.; Srivastava, A.K.; Zou, Y.N.; Kuča, K.; Hashem, A.; Abd_Allah, E.F.; Wu, Q.S. Differential effects of exogenous glomalin-related soil proteins on plant growth of trifoliate orange through regulating auxin changes. Front. Plant Sci. 2021, 12, 745402. [Google Scholar] [CrossRef]
- Wu, Q.S.; Li, Y.; Zou, Y.N.; He, X.H. Arbuscular mycorrhiza mediates glomalin-related soil protein production and soil enzyme activities in the rhizosphere of trifoliate orange grown under different P levels. Mycorrhiza 2015, 25, 121–130. [Google Scholar] [CrossRef]
- Tchiechoua, Y.H.; Kinyua, J.; Ngumi, V.W.; Odee, D.W. Effect of indigenous and introduced arbuscular mycorrhizal fungi on growth and phytochemical content of vegetatively propagated Prunus africana (Hook. f.) Kalkman provenances. Plants 2020, 9, 37. [Google Scholar] [CrossRef] [Green Version]
- Shabani, E.; Bolandnazar, S.; Tabatabaei, S.J. Magnetized phosphorus solution and mycorrhization with Diversispora versiformis affect P use efficiency, growth and photosynthetic parameters in sweet basil (Ocimum basilicum). Hortic. Res. 2019, 27, 103–112. [Google Scholar] [CrossRef] [Green Version]
- Crossay, T.; Majorel, C.; Redecker, D.; Gensous, S.; Amir, H. Is a mixture of arbuscular mycorrhizal fungi better for plant growth than single-species inoculants? Mycorrhiza 2019, 29, 325–339. [Google Scholar] [CrossRef]
- Tuheteru, F.D.; Arif, A. The potential of arbuscular mycorrhizal fungi to conserve Kalappia celebica, an endangered endemic legume on gold mine tailings in Sulawesi, Indonesia. J. For. Res. 2021, 32, 675–682. [Google Scholar]
- Singh, N.V.; Singh, S.K.; Singh, A.K.; Meshram, D.T.; Suroshe, S.S.; Mishra, D.C. Arbuscular mycorrhizal fungi (AMF) induced hardening of micropropagated pomegranate (punica granatum L.) plantlets. Sci. Hortic. 2012, 136, 122–127. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatments | Polar Diameter (mm) | Equatorial Diameter (mm) | Single Fruit Weight (g) | Coloration Value | Fruit Peel Weight (g) | Sarcocarp Weight (g) | Soluble Solids Content (%) |
|---|---|---|---|---|---|---|---|
| Non-AMF | 32.0 ± 0.8 b | 41.6 ± 1.2 c | 33.2 ± 2.5 b | 72.4 ± 1.3 b | 7.3 ± 0.4 c | 24.7 ± 2.1 b | 11.5 ± 0.8 b |
| FM | 36.5 ± 0.6 a | 47.1 ± 1.1 b | 43. 9 ± 2.7 a | 74.4 ± 1.7 a | 11.4 ± 0.9 b | 33.8 ± 1.6 a | 12.6 ± 0.8 a |
| Mixed-AMF | 36.5 ± 1.2 a | 49.2 ± 1.2 a | 46.1 ± 2.0 a | 75.8 ± 0.9 a | 12.4 ± 0.5 a | 35.3 ± 2.5 a | 13.2 ± 0.7 a |
| Correlation | |||||||
| Root AM colonization | 0.65 ** | 0.66 ** | 0.62 ** | 0.67 ** | 0.59 * | 0.71 ** | 0.54 * |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cao, M.-A.; Wang, P.; Hashem, A.; Wirth, S.; Abd_Allah, E.F.; Wu, Q.-S. Field Inoculation of Arbuscular Mycorrhizal Fungi Improves Fruit Quality and Root Physiological Activity of Citrus. Agriculture 2021, 11, 1297. https://doi.org/10.3390/agriculture11121297
Cao M-A, Wang P, Hashem A, Wirth S, Abd_Allah EF, Wu Q-S. Field Inoculation of Arbuscular Mycorrhizal Fungi Improves Fruit Quality and Root Physiological Activity of Citrus. Agriculture. 2021; 11(12):1297. https://doi.org/10.3390/agriculture11121297
Chicago/Turabian StyleCao, Ming-Ao, Peng Wang, Abeer Hashem, Stephan Wirth, Elsayed Fathi Abd_Allah, and Qiang-Sheng Wu. 2021. "Field Inoculation of Arbuscular Mycorrhizal Fungi Improves Fruit Quality and Root Physiological Activity of Citrus" Agriculture 11, no. 12: 1297. https://doi.org/10.3390/agriculture11121297
APA StyleCao, M.-A., Wang, P., Hashem, A., Wirth, S., Abd_Allah, E. F., & Wu, Q.-S. (2021). Field Inoculation of Arbuscular Mycorrhizal Fungi Improves Fruit Quality and Root Physiological Activity of Citrus. Agriculture, 11(12), 1297. https://doi.org/10.3390/agriculture11121297