
Genetic Mapping of the Gamete Eliminator Locus, S2, Causing Hybrid Sterility and Transmission Ratio Distortion Found between Oryza sativa and Oryza glaberrima Cross Combination

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Genetic Stocks
2.2. DNA Marker Survey
2.3. Genetic Mapping Using a Segregating Population
2.4. Seed and Pollen Fertility
3. Results
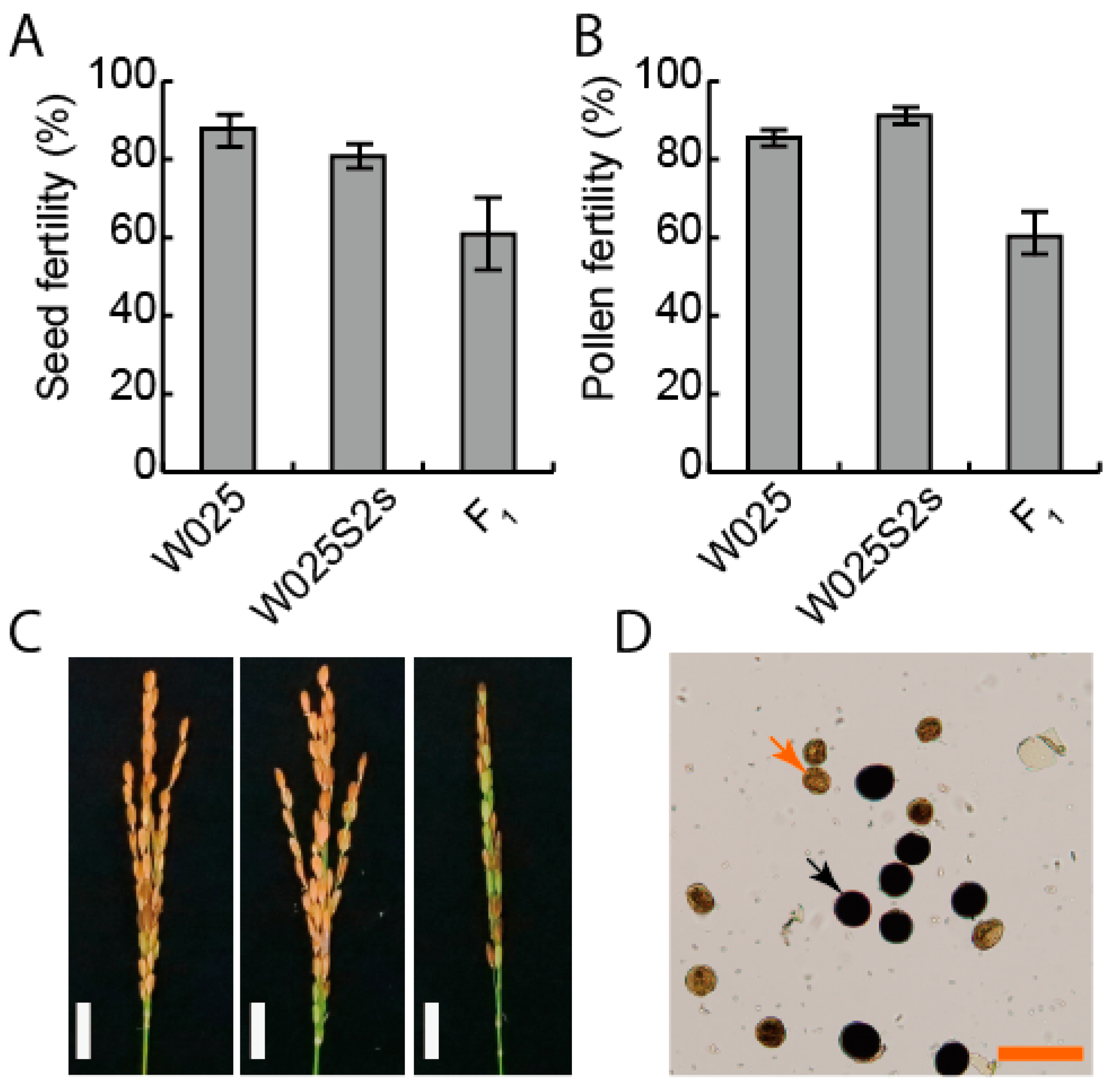
3.1. The S2 Locus Causes Pollen and Seed Sterility in Heterozygotes
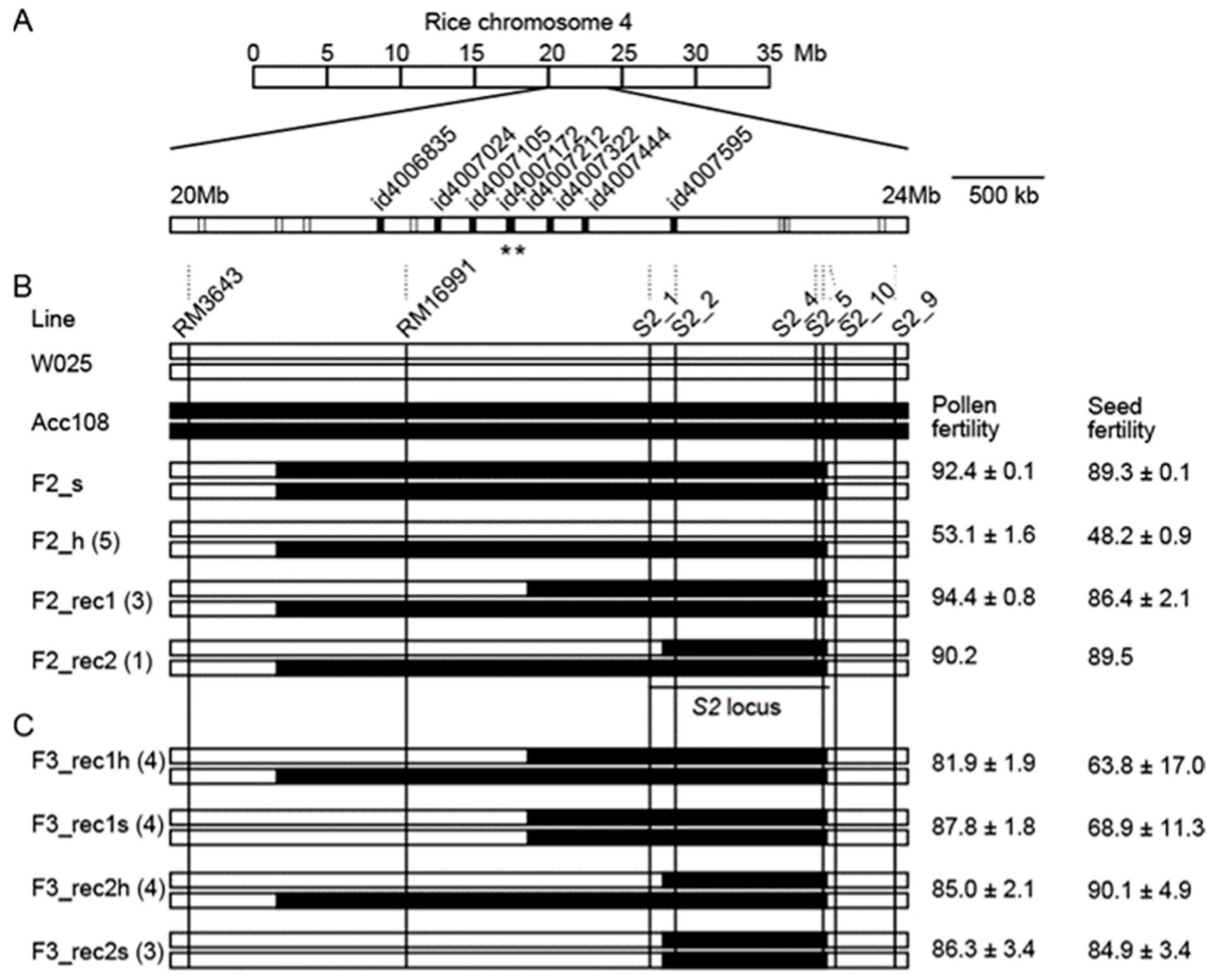
3.2. Chromosomal Regions Introgressed into W025S2s
3.3. Transmission Ratio Distortion Observed in the F2 Population
3.4. Substitution Mapping of the S2 Locus
3.5. Transmission Ratio Distortion Observed in Anther-Culture Derived Calli
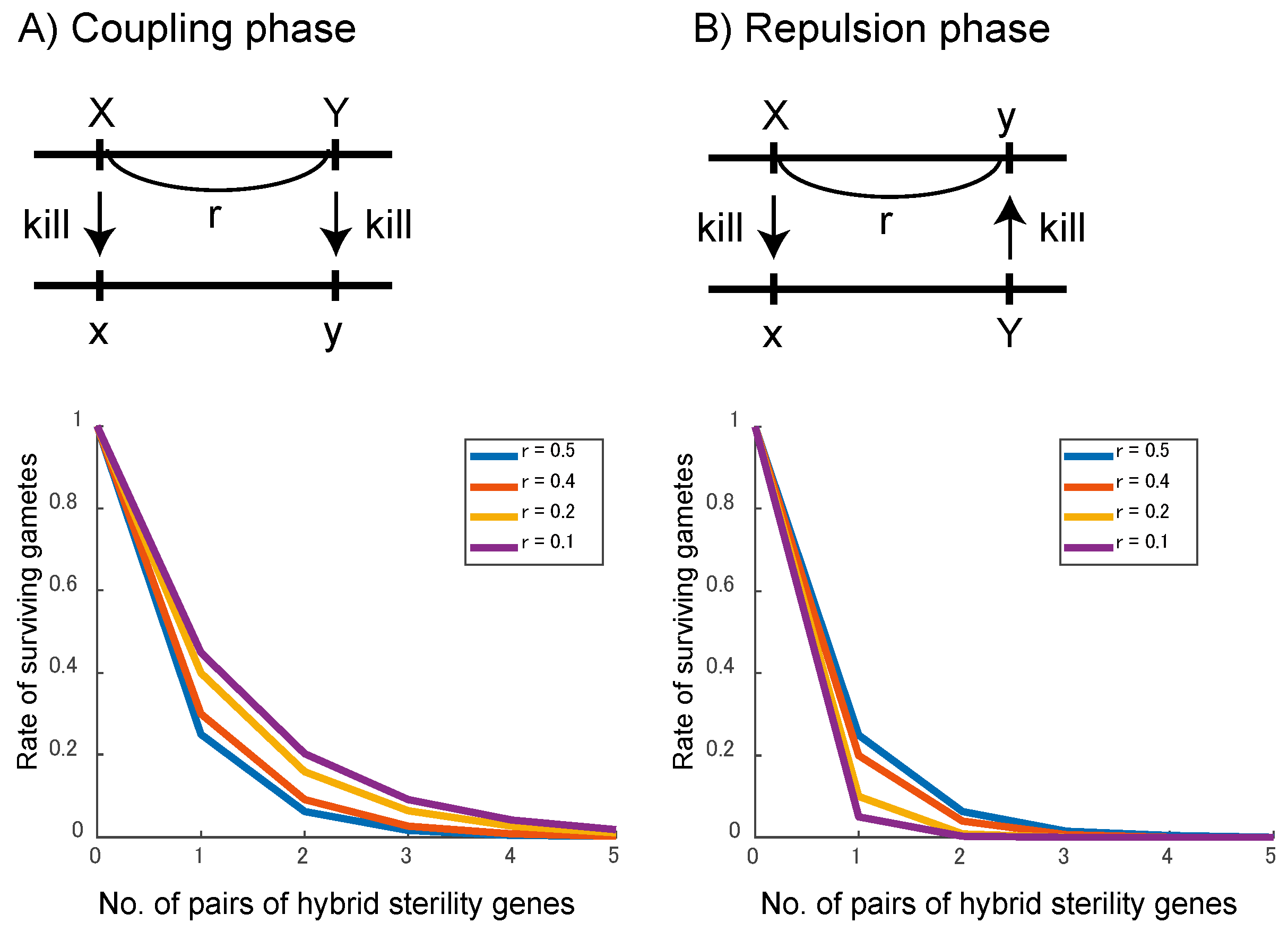
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Chu, Y.E.; Morishima, H.; Oka, H.I. Reproductive barriers distributed in cultivated rice species and their wild relatives. Jpn. J. Genet. 1969, 4, 207–223. [Google Scholar] [CrossRef] [Green Version]
- Bouharmont, J.; Oliver, M.; Dumont de Chassart, M. Cytological observation in some hybrids between the rice species Oryza sativa L. and O. glaberrima Stetud. Euphytica 1985, 34, 75–81. [Google Scholar] [CrossRef]
- Jones, M.P.; Mande, S.; Aluko, K. Diversity and potential of Oryza glaberrima Steud in upland rice breeding. Breed. Sci. 1997, 47, 395–398. [Google Scholar] [CrossRef] [Green Version]
- Oka, H.I. Considerations on the genetic basis of intervarietal sterility in Oryza sativa. In Rice Genetics and Cytogenetics; Chandler, R.F., Ed.; Elsevier: Amsterdam, The Netherlands, 1964; pp. 158–174. [Google Scholar]
- Sano, Y.; Chu, Y.E.; Oka, H.I. Genetic studies of speciation in cultivated rice, 1. Genic analysis for the F1 sterility between O. sativa L. and O. glaberrima Steud. J. Genet. 1979, 54, 121–132. [Google Scholar]
- Doi, K.; Yasui, H.; Yoshimura, A. Genetic variation in rice. Curr. Opin. Plant Biol. 2008, 11, 144–148. [Google Scholar] [CrossRef] [PubMed]
- Koide, Y.; Onishi, K.; Kanazawa, A.; Sano, Y. Genetics of speciation in rice. In Rice Biology in the Genomics Era; Hirano, H.Y., Hirai, A., Sano, Y., Sasaki, T., Eds.; Springer: Berlin, Germany, 2008; pp. 247–259. [Google Scholar]
- Kanaoka, Y.; Kuniyoshi, D.; Inada, E.; Koide, Y.; Okamoto, Y.; Yasui, H.; Kishima, Y. Anther culture in rice proportionally rescues microspores according to gametophytic gene effect and enhances genetic study of hybrid sterility. Plant Methods 2018, 14, 102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sano, Y. The genic nature of gamete eliminator in rice. Genetics 1990, 125, 183–191. [Google Scholar] [CrossRef] [PubMed]
- Xu, P.; Zhou, J.W.; Li, J.; Hu, F.Y.; Deng, X.N.; Feng, S.F.; Ren, G.Y.; Zhang, Z.; Deng, W.; Tao, D.Y. Mapping three new interspecific hybrid sterile loci between Oryza sativa and O. glaberrima. Breed. Sci. 2014, 63, 476–482. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shen, Y.M.; Zhao, Z.G.; Ma, H.Y.; Bian, X.F.; Yu, Y.; Yu, X.W.; Chen, H.Y.; Liu, L.L.; Zhang, W.W.; Jiang, L.; et al. Fine mapping of S37, a locus responsible for pollen and embryo sac sterility in hybrids between Oryza sativa L. and O. glaberrima Steud. Plant Cell Rep. 2015, 34, 1885–1897. [Google Scholar] [CrossRef] [PubMed]
- Xie, Y.; Xu, P.; Huang, J.; Ma, S.; Xie, X.; Tao, D.; Chen, L.; Liu, Y.G. Interspecific hybrid sterility in rice is mediated by OgTPR1 at the S1 locus encoding a peptidase-like protein. Mol. Plants 2017, 10, 1137–1140. [Google Scholar] [CrossRef] [PubMed]
- Koide, Y.; Ogino, A.; Yoshikawa, T.; Kitashima, Y.; Saito, N.; Kanaoka, Y.; Onishi, K.; Yoshitake, Y.; Tsukiyama, T.; Saito, H.; et al. Lineage-specific gene acquisition or loss is involved in interspecific hybrid sterility in rice. Proc. Natl. Acad. Sci. USA 2018, 115, E1955–E1962. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xie, Y.; Tang, J.; Xie, X.; Li, X.; Huang, J.; Fei, Y.; Han, J.; Chen, S.; Tang, H.; Zhao, X.D.; et al. Chen. An asymmetric allelic interaction drives allele transmission bias in interspecific rice hybrids. Nat. Commun. 2019, 10, 2501. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pariaska-Tanaka, J.; Lorieux, M.; He, C.; McCouch, S.; Thomson, M.J.; Wissuwa, M. Development of a SNP genotyping panel for detecting polymorphisms in Oryza glaberrima/O. sativa interspecific crosses. Euphytica 2015, 201, 67–78. [Google Scholar] [CrossRef]
- Yasuda, K.; Ito, M.; Sugita, T.; Tsukiyama, T.; Saito, H.; Naito, K.; Teraishi, M.; Tanisaka, T.; Okumoto, Y. Utilization of transposable element mPing as a novel genetic tool for modification of the stress response in rice. Mol. Breed. 2013, 32, 505–516. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koide, Y.; Ikenaga, M.; Sawamura, N.; Nishimoto, D.; Matsubara, K.; Onishi, K.; Kanazawa, A.; Sano, Y. The evolution of sex-independent transmission ratio distortion involving multiple allelic interactions at a single locus in rice. Genetics 2008, 180, 409–420. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koide, Y.; Onishi, K.; Nishimoto, D.; Baruah, A.R.; Kanazawa, A.; Sano, Y. Sex-independent transmission ratio distortion system responsible for reproductive barriers between Asian and African rice species. New Phytol. 2008, 179, 888–900. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Plant No. | P2 | ||||
|---|---|---|---|---|---|
| Marker Name | Location (Mb) 1 | s/s | s/g | g/g | (1:2:1) |
| RM16991 | 21.27 | 134 | 16 | 0 | 7.16145 × 10−73 |
| S2_1 | 22.08 | 142 | 9 | 0 | 3.69831 × 10−84 |
| S2_2 | 22.15 | 144 | 7 | 0 | 2.34291 × 10−87 |
| S2_4 | 22.74 | 144 | 7 | 0 | 2.34291 × 10−87 |
| S2_5 | 23.50 | 143 | 8 | 0 | 9.49528 × 10−86 |
| No. of Calli | P2 | |||
|---|---|---|---|---|
| Marker Name | Location (Mb) 1 | s/s | g/g | (1:1) |
| S2_4 | 22.74 | 24 | 3 | 5.31 × 10−5 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zin Mar, M.; Koide, Y.; Ogata, M.; Kuniyoshi, D.; Tokuyama, Y.; Hikichi, K.; Obara, M.; Kishima, Y. Genetic Mapping of the Gamete Eliminator Locus, S2, Causing Hybrid Sterility and Transmission Ratio Distortion Found between Oryza sativa and Oryza glaberrima Cross Combination. Agriculture 2021, 11, 268. https://doi.org/10.3390/agriculture11030268
Zin Mar M, Koide Y, Ogata M, Kuniyoshi D, Tokuyama Y, Hikichi K, Obara M, Kishima Y. Genetic Mapping of the Gamete Eliminator Locus, S2, Causing Hybrid Sterility and Transmission Ratio Distortion Found between Oryza sativa and Oryza glaberrima Cross Combination. Agriculture. 2021; 11(3):268. https://doi.org/10.3390/agriculture11030268
Chicago/Turabian StyleZin Mar, Myint, Yohei Koide, Mei Ogata, Daichi Kuniyoshi, Yoshiki Tokuyama, Kiwamu Hikichi, Mitsuhiro Obara, and Yuji Kishima. 2021. "Genetic Mapping of the Gamete Eliminator Locus, S2, Causing Hybrid Sterility and Transmission Ratio Distortion Found between Oryza sativa and Oryza glaberrima Cross Combination" Agriculture 11, no. 3: 268. https://doi.org/10.3390/agriculture11030268
APA StyleZin Mar, M., Koide, Y., Ogata, M., Kuniyoshi, D., Tokuyama, Y., Hikichi, K., Obara, M., & Kishima, Y. (2021). Genetic Mapping of the Gamete Eliminator Locus, S2, Causing Hybrid Sterility and Transmission Ratio Distortion Found between Oryza sativa and Oryza glaberrima Cross Combination. Agriculture, 11(3), 268. https://doi.org/10.3390/agriculture11030268