
Plant Growth-Promoting Rhizobacteria Isolated from the Jujube (Ziziphus lotus) Plant Enhance Wheat Growth, Zn Uptake, and Heavy Metal Tolerance

 ,
,  ,
,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sampling and Screening of Phosphate Solubilizing Rhizobacteria
2.2. Quantitative Phosphate Solubilization Assay
2.3. 16S rRNA Gene Sequencing and Phylogenetic Analysis
2.4. Indole Acetic Acid Production Assay
2.5. Zn Solubilization Assay
2.6. Siderophores Production Assay
2.7. Extracellular Enzyme Assays
2.8. Ammonia Production Assay
2.9. Biofilm Formation Assay
2.10. Antibiotics Resistance and Heavy Metal Tolerance
2.11. NaCl Tolerance and Temperature Growth Monitoring
2.12. Seed Germination Assay
2.13. Inoculation of Wheat and Plant Growth Monitoring
2.14. Statistical Analysis
3. Results
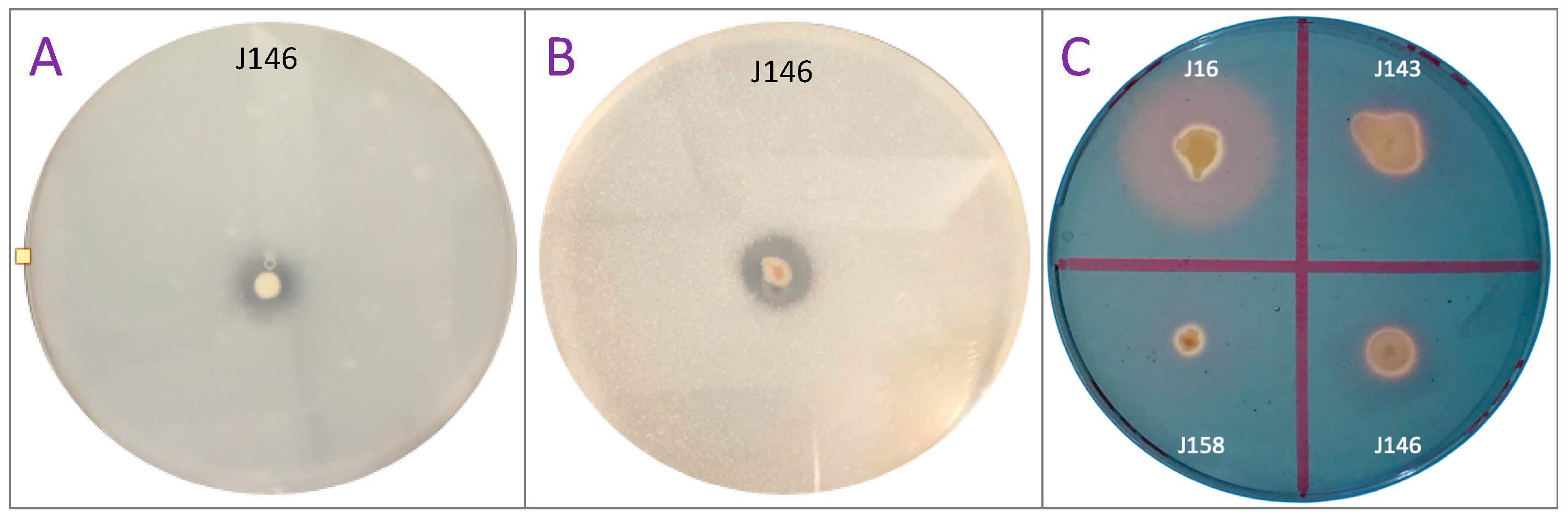
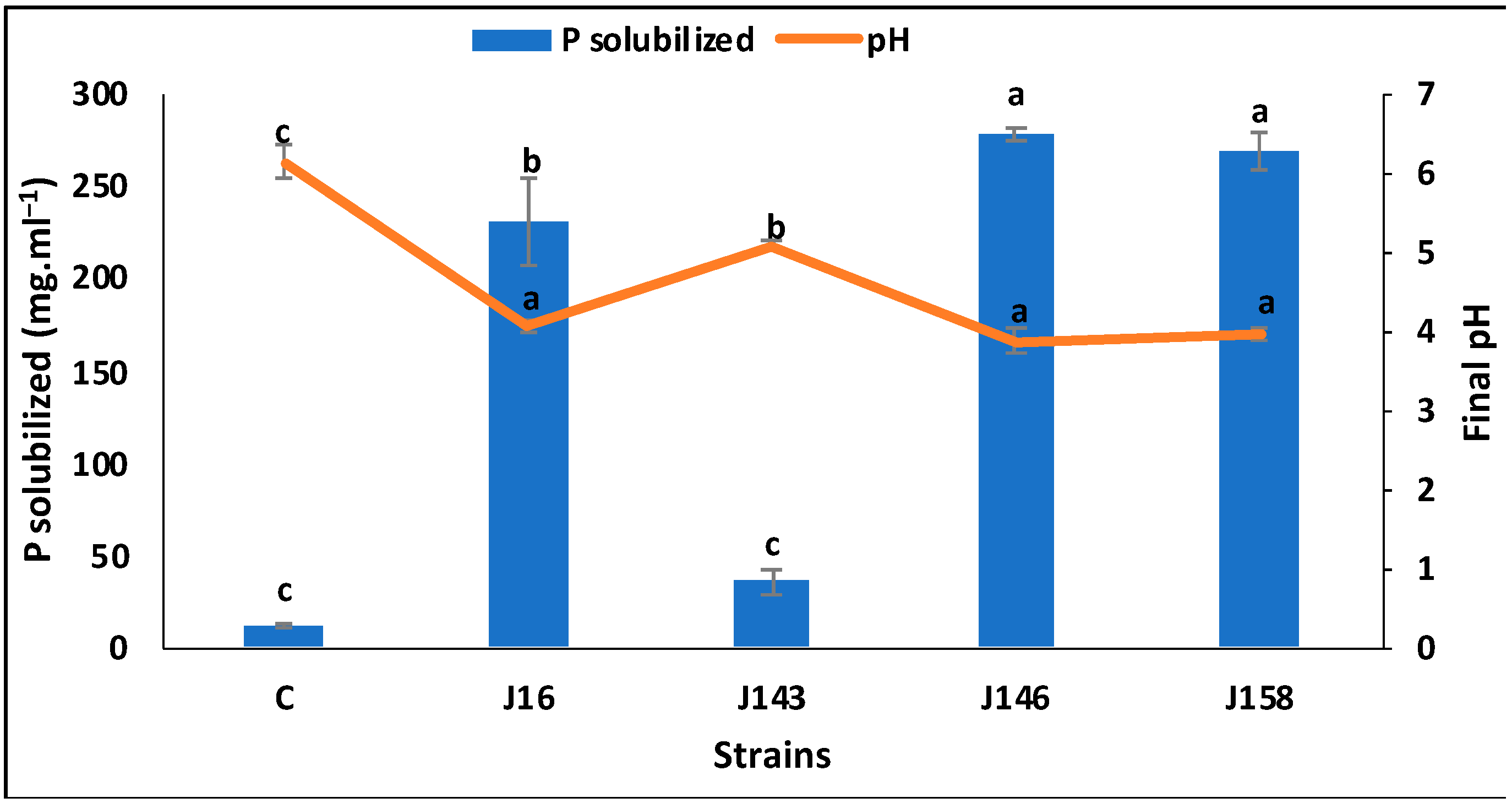
3.1. Screening of the Four P-Solubilizing, Jujube-Associated Rhizobacteria (J16, J143, J146, and J158)
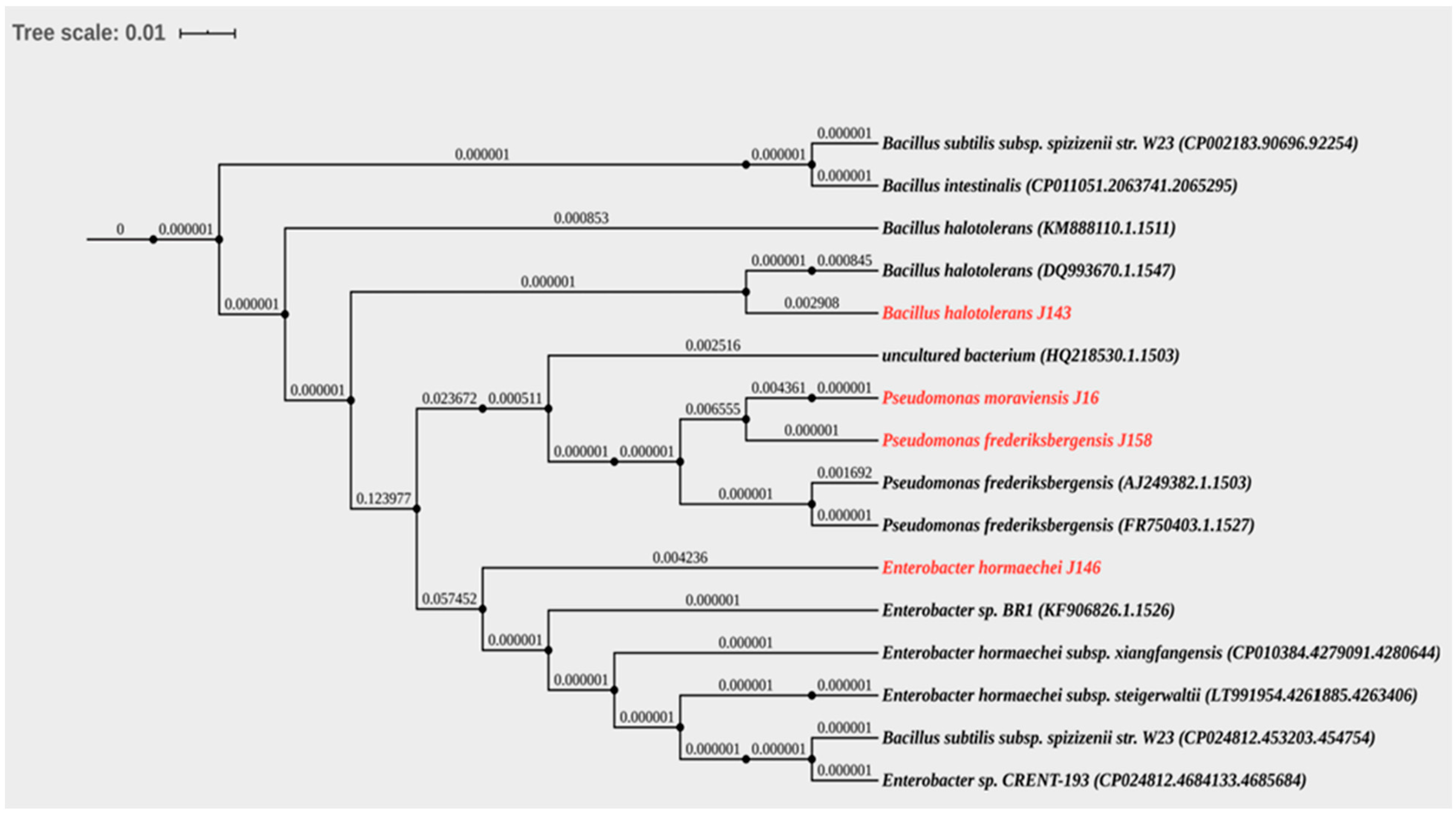
3.2. The Isolates J16, J143, J146, and J158 Belong to the Pseudomonas, Enterobacter, and Bacillus genera
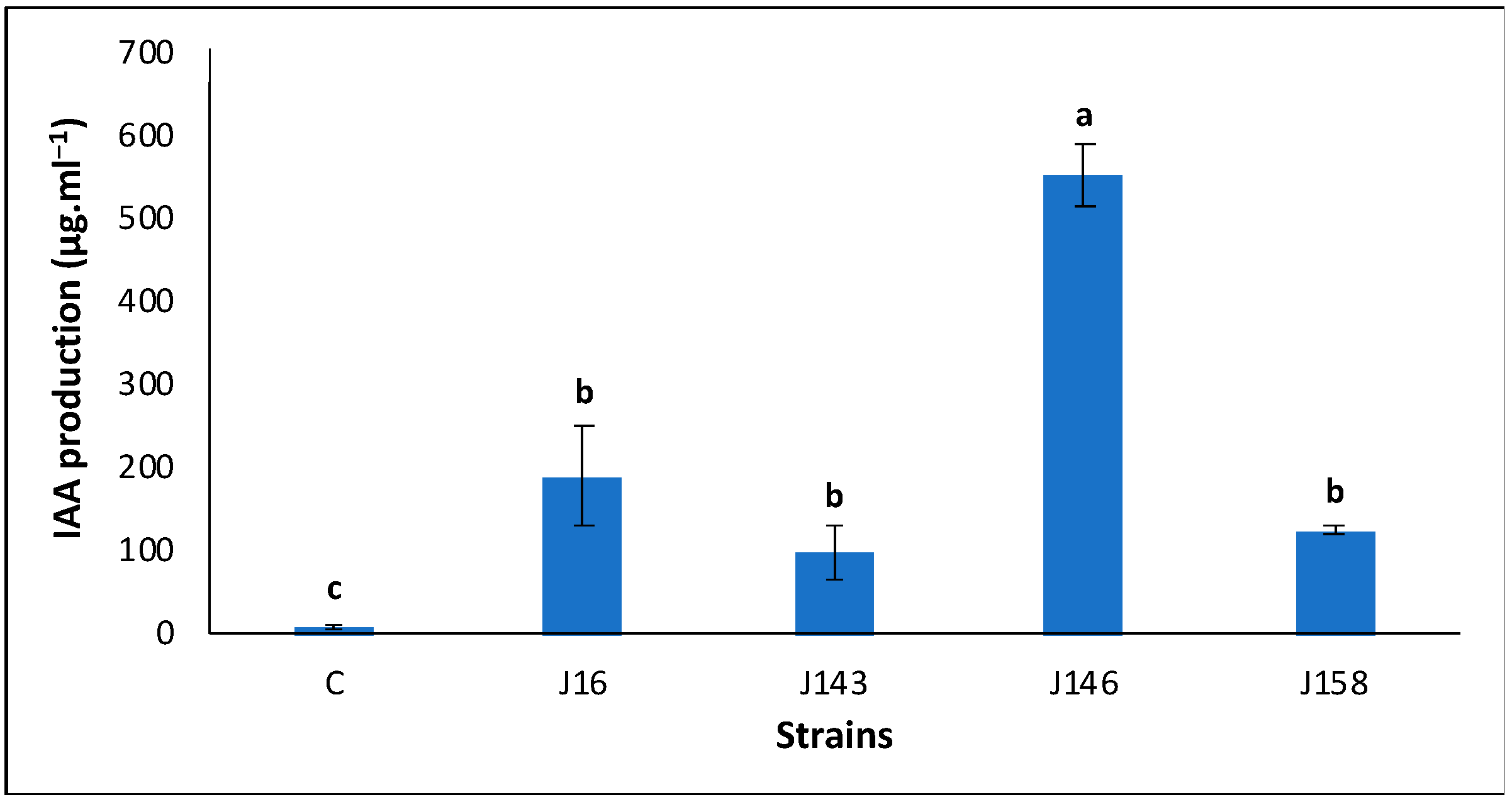
3.3. E. hormaechei J146 Overproduced Indole-3-Acetic Acid (IAA)
3.4. Zn Solubilization from Zinc-Oxide by P. moraviensis J16, E. hormaechei J146, and P. frederiksbergensis J158
3.5. Strain P. moraviensis J16 Is the Best Siderophores Producer
3.6. Lack of Proteases and Cellulase in E. hormaechei J146 and P. frederiksbergensis J158
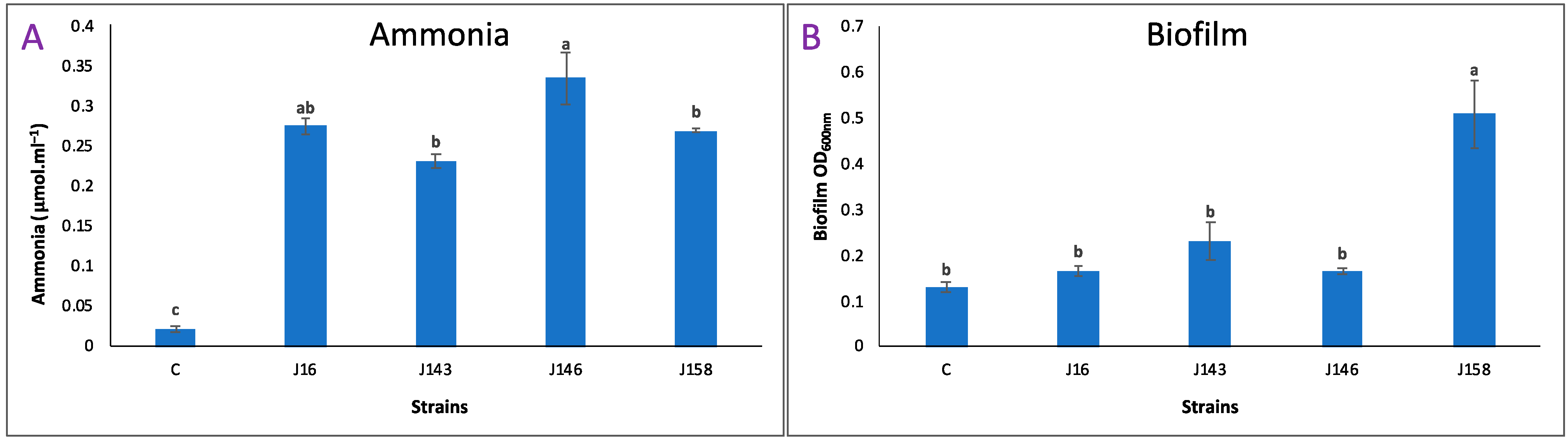
3.7. Strain E. hormaechei J146 Overproduced Ammonia
3.8. Strain P. frederiksbergensis J158 Induced a Strong Biofilm
3.9. PSB Strains Displayed Partial Antibiotics Resistance
3.10. E. hormaechei J146 and B. halotolerans J143 Withstood Copper/Cadmium and Nickel, Respectively
3.11. E. hormaechei J146 and B. halotolerans J143 Tolerated Salt Stress
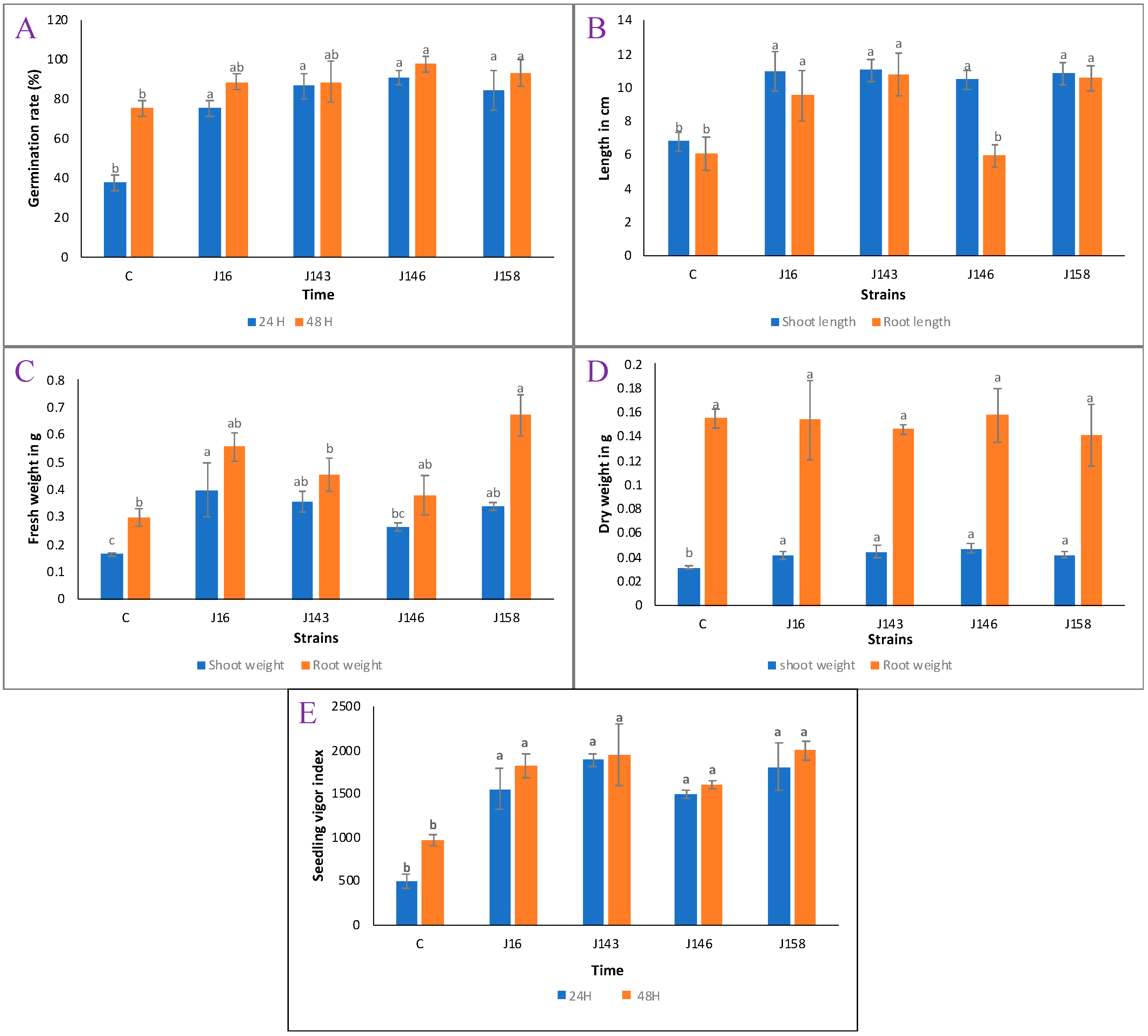
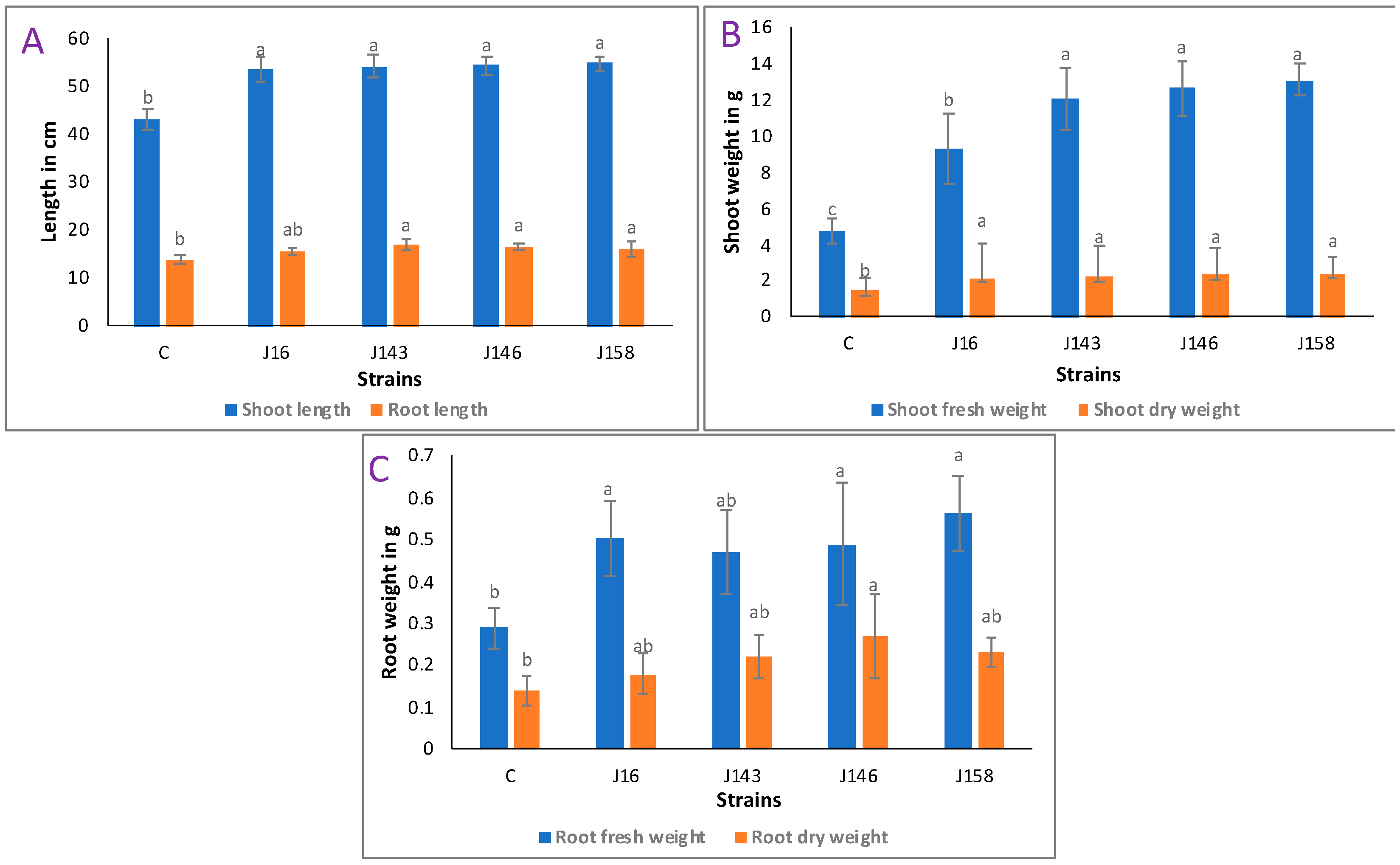
3.12. E. hormaechei J146 Strain Is the Best Inducer of Seeds Germination
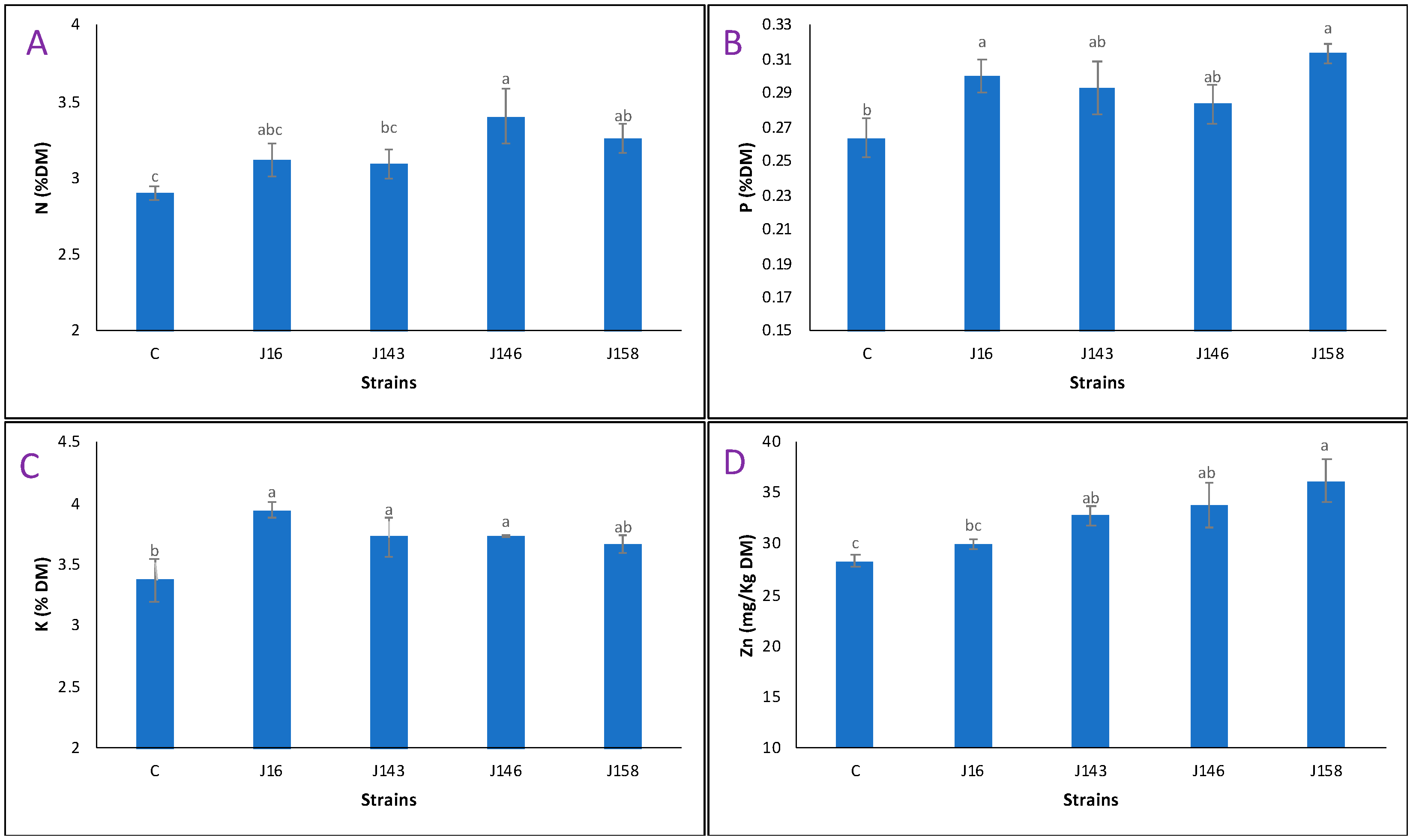
3.13. P. frederiksbergensis J158 Induced the Best Wheat Plant Nutrients Uptake
4. Discussion
4.1. Effect of Studied PSB Strains on Wheat Growth
4.2. Effect of the Four PSB Strains on Nutrient Uptake and Stress Tolerance
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Reetz, H.F. Fertilizers and Their Efficient Use; International Fertilizer industry Association: Paris, France, 2016; ISBN 979-10-92366-04-4. [Google Scholar]
- FAO. The Future of Food and Agriculture: Trends and Challenges; FAO: Rome, Italy, 2017; pp. 1–180. [Google Scholar]
- FAO. The State of Agricultural Commodity Markets 2018. Agricultural Trade, Climate Change and Food Security; FAO: Rome, Italy, 2018. [Google Scholar]
- Soumare, A.; Boubekri, K.; Lyamlouli, K.; Hafidi, M.; Ouhdouch, Y.; Kouisni, L. From Isolation of Phosphate Solubilizing Microbes to Their Formulation and Use as Biofertilizers: Status and Needs. Front. Bioeng. Biotechnol. 2020, 7, 425. [Google Scholar] [CrossRef] [PubMed]
- Asher, C. Crop Nutrition during the Establishment Phase: Role of Seed Reserves; Occasional Publication; Australian Institute of Agricultural Science: St Leonards, Australia, 1987. [Google Scholar]
- Hafeez, B.; Khanif, Y.M.; Saleem, M. Role of Zinc in Plant Nutrition—A Review. Am. J. Exp. Agric. 2013, 3, 374–391. [Google Scholar] [CrossRef]
- Alloway, B. Zinc in Soils and Crop Nutrition; IZA Publications; International Zinc Association Communications: Brussels, Belgium, 2004. [Google Scholar]
- Alloway, B.J. Micronutrient Deficiencies in Global Crop Production; Springer Science & Business Media: Berlin/Heidelberg, Germany, 2008; ISBN 1-4020-6860-3. [Google Scholar]
- Graham, R.D.; Welch, R.M.; Bouis, H.E. Addressing micronutrient malnutrition through enhancing the nutritional quality of staple foods: Principles, perspectives and knowledge gaps. In Advances in Agronomy Volume 40; Elsevier BV: Amsterdam, The Netherlands, 2001; Volume 70, pp. 77–142. [Google Scholar] [CrossRef]
- Yang, X.-W.; Tian, X.-J.; Lu, X.-C.; William, G.; Cao, Y.-X. Foliar Zinc Fertilization Improves the Zinc Nutritional Value of Wheat (Triticum Aestivum L.) Grain. Afr. J. Biotechnol. 2011, 10, 14778–14785. [Google Scholar]
- Mousavi, S.R. Zinc in Crop Production and Interaction with Phosphorus. Aust. J. Basic Appl. Sci. 2011, 5, 1503–1509. [Google Scholar]
- Ali, S.; Khan, A.R.; Mairaj, G.; Arif, M.; Fida, M.; Bibi, S. Assessment of Different Crop Nutrient Management Practices for Yield Improvement. Aust. J. Crop Sci. 2008, 2, 150–157. [Google Scholar]
- Welch, R.M.; Graham, R.D. Breeding for micronutrients in staple food crops from a human nutrition perspective. J. Exp. Bot. 2004, 55, 353–364. [Google Scholar] [CrossRef] [Green Version]
- Sandstead, H.H. Is zinc deficiency a public health problem? Nutrition 1995, 11, 853–859. [Google Scholar]
- Bargaz, A.; Lyamlouli, K.; Chtouki, M.; Zeroual, Y.; Dhiba, D. Soil Microbial Resources for Improving Fertilizers Efficiency in an Integrated Plant Nutrient Management System. Front. Microbiol. 2018, 9, 1606. [Google Scholar] [CrossRef] [Green Version]
- Sharma, S.B.; Sayyed, R.Z.; Trivedi, M.H.; Gobi, T.A. Phosphate solubilizing microbes: Sustainable approach for managing phosphorus deficiency in agricultural soils. SpringerPlus 2013, 2, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.; Rekha, P.; Arun, A.; Shen, F.; Lai, W.-A.; Young, C. Phosphate solubilizing bacteria from subtropical soil and their tricalcium phosphate solubilizing abilities. Appl. Soil Ecol. 2006, 34, 33–41. [Google Scholar] [CrossRef]
- Boubekri, K.; Soumare, A.; Mardad, I.; Lyamlouli, K.; Hafidi, M.; Ouhdouch, Y.; Kouisni, L. The Screening of Potassium- and Phosphate-Solubilizing Actinobacteria and the Assessment of Their Ability to Promote Wheat Growth Parameters. Microorganisms 2021, 9, 470. [Google Scholar] [CrossRef]
- Pérez-Montaño, F.; Alías-Villegas, C.; Bellogín, R.; del Cerro, P.; Espuny, M.; Jiménez-Guerrero, I.; López-Baena, F.; Ollero, F.; Cubo, T. Plant growth promotion in cereal and leguminous agricultural important plants: From microorganism capacities to crop production. Microbiol. Res. 2014, 169, 325–336. [Google Scholar] [CrossRef] [Green Version]
- Khan, M.S.; Zaidi, A.; Ahemad, M.; Oves, M.; Wani, P.A. Plant growth promotion by phosphate solubilizing fungi—Current perspective. Arch. Agron. Soil Sci. 2010, 56, 73–98. [Google Scholar] [CrossRef]
- Selvakumar, G.; Mohan, M.; Kundu, S.; Gupta, A.; Joshi, P.; Nazim, S.; Gupta, H. Cold tolerance and plant growth promotion potential of Serratia marcescens strain SRM (MTCC 8708) isolated from flowers of summer squash (Cucurbita pepo). Lett. Appl. Microbiol. 2007, 46, 171–175. [Google Scholar] [CrossRef] [PubMed]
- Whitelaw, M. Growth Promotion of Plants Inoculated with Phosphate-Solubilizing Fungi. In Advances in Agronomy Volume 40; Elsevier BV: Amsterdam, The Netherlands, 1999; Volume 69, pp. 99–151. [Google Scholar]
- Kang, Y.; Shen, M.; Xia, D.; Ye, K.; Zhao, Q.; Hu, J. Caution of intensified spread of antibiotic resistance genes by inadvertent introduction of beneficial bacteria into soil. Acta Agric. Scand. Sect. B Plant Soil Sci. 2017, 67, 576–582. [Google Scholar] [CrossRef]
- Kluepfel, D.A. The Behavior and Tracking of Bacteria in the Rhizosphere. Annu. Rev. Phytopathol. 1993, 31, 441–472. [Google Scholar] [CrossRef]
- Trivedi, P.; Pandey, A.; Palni, L.M.S.; Bag, N.; Tamang, M. Colonization of Rhizosphere of Tea by Growth Promoting Bacteria. Int. Soc. Tea Sci. 2004, 4. Available online: http://hdl.handle.net/2263/8370 (accessed on 12 December 2020).
- Cray, J.A.; Bell, A.N.W.; Bhaganna, P.; Mswaka, A.Y.; Timson, D.J.; Hallsworth, J.E. The biology of habitat dominance; can microbes behave as weeds? Microb. Biotechnol. 2013, 6, 453–492. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramakrishna, W.; Yadav, R.; Li, K. Plant growth promoting bacteria in agriculture: Two sides of a coin. Appl. Soil Ecol. 2019, 138, 10–18. [Google Scholar] [CrossRef]
- Wang, B.; Huang, Q.; Venkitasamy, C.; Chai, H.; Gao, H.; Cheng, N.; Cao, W.; Lv, X.; Pan, Z. Changes in phenolic compounds and their antioxidant capacities in jujube (Ziziphus jujuba Miller) during three edible maturity stages. LWT 2016, 66, 56–62. [Google Scholar] [CrossRef]
- Rsaissi, N.; Bencharki, B. Importance and Agro-Economical Impact of Wild Jujube (Ziziphus Lotus) in Chaouia Region. Rev. Maroc. Prot. Plantes 2012, 3, 13–27. [Google Scholar]
- Nautiyal, C.S. An Efficient Microbiological Growth Medium for Screening Phosphate Solubilizing Microorganisms. FEMS Microbiol. Lett. 1999, 170, 265–270. [Google Scholar] [CrossRef] [PubMed]
- Mukhtar, S.; Shahid, I.; Mehnaz, S.; Malik, K.A. Assessment of two carrier materials for phosphate solubilizing biofertilizers and their effect on growth of wheat (Triticum aestivum L.). Microbiol. Res. 2017, 205, 107–117. [Google Scholar] [CrossRef]
- Rahman, C.H.; Ahcene, B.; Miloud, B.; Rachid, D. Screening and Characterization of Plant Growth Promoting Traits of Phosphate Solubilizing Bacteria Isolated from Wheat Rhizosphere of Algerian Saline Soil. Malays. J. Microbiol. 2017, 13, 124–131. [Google Scholar]
- Kumar, V.; Behl, R.K.; Narula, N. Establishment of Phosphate-Solubilizing Strains of Azotobacter Chroococcum in the Rhi-zosphere and Their Effect on Wheat Cultivars under Green House Conditions. Microbiol. Res. 2001, 156, 87–93. [Google Scholar] [CrossRef]
- Okonechnikov, K.; Golosova, O.; Fursov, M.; UGENE Team. Unipro UGENE: A unified bioinformatics toolkit. Bioinformatics 2012, 28, 1166–1167. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Glickmann, E.; Dessaux, Y. A Critical Examination of the Specificity of the Salkowski Reagent for Indolic Compounds Pro-duced by Phytopathogenic Bacteria. Appl. Environ. Microbiol. 1995, 61, 793–796. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gandhi, A.; Muralidharan, G. Assessment of zinc solubilizing potentiality of Acinetobacter sp. isolated from rice rhizosphere. Eur. J. Soil Biol. 2016, 76, 1–8. [Google Scholar] [CrossRef]
- Schwyn, B.; Neilands, J. Universal chemical assay for the detection and determination of siderophores. Anal. Biochem. 1987, 160, 47–56. [Google Scholar] [CrossRef]
- Kavitha, T.; Nelson, R.; Jesi, S. Screening of Rhizobacteria for Plant Growth Promoting Traits and Antifungal Activity against Charcoal Rot Pathogen Macrophomina Phaseolina. Int. J. Pharma Bio Sci. 2013, 4, B-177–B-186. [Google Scholar]
- Kasana, R.C.; Salwan, R.; Dhar, H.; Dutt, S.; Gulati, A. A Rapid and Easy Method for the Detection of Microbial Cellulases on Agar Plates Using Gram’s Iodine. Curr. Microbiol. 2008, 57, 503–507. [Google Scholar] [CrossRef]
- Cappuccino, J.; Sherman, N. Microbiology: A Laboratory Manual; Benjamin/Cumming Pub. Co.: New York, NY, USA, 1992; pp. 125–179. [Google Scholar]
- Chrouqi, L.; Lahcen, O.; Jadrane, I.; Koussa, T.; Alfeddy, M.N. Screening of Soil Rhizobacteria Isolated from Wheat Plants Grown in the Marrakech Region (Morocco, North Africa) for Plant Growth Promoting Activities. JMES 2017, 8, 3382–3390. [Google Scholar]
- Christensen, G.D.; Simpson, W.A.; Younger, J.; Baddour, L.; Barrett, F.; Melton, D.; Beachey, E. Adherence of Coagu-lase-Negative Staphylococci to Plastic Tissue Culture Plates: A Quantitative Model for the Adherence of Staphylococci to Medical Devices. J. Clin. Microbiol. 1985, 22, 996–1006. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Armalytė, J.; Skerniškytė, J.; Bakienė, E.; Krasauskas, R.; Šiugždinienė, R.; Kareivienė, V.; Kerzienė, S.; Klimienė, I.; Sužiedėlienė, E.; Ružauskas, M. Microbial Diversity and Antimicrobial Resistance Profile in Microbiota from Soils of Conventional and Organic Farming Systems. Front. Microbiol. 2019, 10, 892. [Google Scholar] [CrossRef]
- Khan, N.; Zandi, P.; Ali, S.; Mehmood, A.; Shahid, M.A.; Yang, J. Impact of Salicylic Acid and PGPR on the Drought Tolerance and Phytoremediation Potential of Helianthus annus. Front. Microbiol. 2018, 9, 2507. [Google Scholar] [CrossRef] [Green Version]
- Biswas, J.K.; Banerjee, A.; Rai, M.; Naidu, R.; Biswas, B.; Vithanage, M.; Dash, M.C.; Sarkar, S.K.; Meers, E. Potential Appli-cation of Selected Metal Resistant Phosphate Solubilizing Bacteria Isolated from the Gut of Earthworm (Metaphire Posthuma) in Plant Growth Promotion. Geoderma 2018, 330, 117–124. [Google Scholar] [CrossRef]
- Patel, K.S.; Naik, J.H.; Chaudhari, S.; Amaresan, N. Characterization of culturable bacteria isolated from hot springs for plant growth promoting traits and effect on tomato (Lycopersicon esculentum) seedling. Comptes Rendus Biol. 2017, 340, 244–249. [Google Scholar] [CrossRef]
- Islam, S.; Akanda, A.M.; Prova, A.; Islam, M.T.; Hossain, M.M. Isolation and Identification of Plant Growth Promoting Rhi-zobacteria from Cucumber Rhizosphere and Their Effect on Plant Growth Promotion and Disease Suppression. Front. Microbiol. 2016, 6, 1360. [Google Scholar] [CrossRef] [Green Version]
- Goldstein, A.H. Bioprocessing of Rock Phosphate Ore: Essential Technical Considerations for the Development of a Successful Commercial Technology. In Proceedings of the 4th International Fertilizer Association Technical Conference; IFA: Paris, France, 2000; Volume 220. [Google Scholar]
- Ahmad, F.; Ahmad, I.; Khan, M. Screening of free-living rhizospheric bacteria for their multiple plant growth promoting activities. Microbiol. Res. 2008, 163, 173–181. [Google Scholar] [CrossRef]
- Salantur, A.; Ozturk, A.; Akten, S. Growth and yield response of spring wheat (Triticum aestivum L.) to inoculation with rhizobacteria. Plant Soil Environ. 2011, 52, 111–118. [Google Scholar] [CrossRef]
- Gupta, A.; Gupta, R.; Singh, R.L. Microbes and Environment. In Principles and Applications of Environmental Biotechnology for a Sustainable Future; Singh, R.L., Ed.; Springer: Singapore, 2017; pp. 43–84. ISBN 978-981-10-1866-4. [Google Scholar]
- Mann, E.E.; Wozniak, D.J. Pseudomonas Biofilm Matrix Composition and Niche Biology. FEMS Microbiol. Rev. 2012, 36, 893–916. [Google Scholar] [CrossRef] [Green Version]
- Jetiyanon, K. Multiple Mechanisms of Enterobacter Asburiae Strain RS83 for Plant Growth Enhancement. Songklanakarin J. Sci. Technol. 2015, 37, 29–36. [Google Scholar]
- Bendaha, M.E.A.; Belaouni, H.A. Tomato Growth and Resistance Promotion by Enterobacter Hormaechei Subsp. Steigerwaltii EB8D. Arch. Phytopathol. Plant Prot. 2019, 52, 318–332. [Google Scholar] [CrossRef]
- Mittal, V.; Singh, O.; Nayyar, H.; Kaur, J.; Tewari, R. Stimulatory effect of phosphate-solubilizing fungal strains (Aspergillus awamori and Penicillium citrinum) on the yield of chickpea (Cicer arietinum L. cv. GPF2). Soil Biol. Biochem. 2008, 40, 718–727. [Google Scholar] [CrossRef]
- Pérez-Flores, P.; Valencia-Cantero, E.; Altamirano-Hernández, J.; Pelagio-Flores, R.; López-Bucio, J.; García-Juárez, P.; Macías-Rodríguez, L. Bacillus methylotrophicus M4-96 isolated from maize (Zea mays) rhizoplane increases growth and auxin content in Arabidopsis thaliana via emission of volatiles. Protoplasma 2017, 254, 2201–2213. [Google Scholar] [CrossRef]
- Prusty, R.; Grisafi, P.; Fink, G.R. The plant hormone indoleacetic acid induces invasive growth in Saccharomyces cerevisiae. Proc. Natl. Acad. Sci. USA 2004, 101, 4153–4157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mahdi, I.; Fahsi, N.; Hafidi, M.; Allaoui, A.; Biskri, L. Plant Growth Enhancement using Rhizospheric Halotolerant Phosphate Solubilizing Bacterium Bacillus licheniformis QA1 and Enterobacter asburiae QF11 Isolated from Chenopodium quinoa Willd. Microorganisms 2020, 8, 948. [Google Scholar] [CrossRef] [PubMed]
- Gontia-Mishra, I.; Sapre, S.; Sharma, A.; Tiwari, S. Alleviation of Mercury Toxicity in Wheat by the Interaction of Mercury-Tolerant Plant Growth-Promoting Rhizobacteria. J. Plant Growth Regul. 2016, 35, 1000–1012. [Google Scholar] [CrossRef]
- Matthijs, S.; Tehrani, K.A.; Laus, G.; Jackson, R.W.; Cooper, R.M.; Cornelis, P. Thioquinolobactin, a Pseudomonas siderophore with antifungal and anti-Pythium activity. Environ. Microbiol. 2007, 9, 425–434. [Google Scholar] [CrossRef] [PubMed]
- Indiragandhi, P.; Anandham, R.; Madhaiyan, M.; Sa, T.M. Characterization of Plant Growth–Promoting Traits of Bacteria Isolated from Larval Guts of Diamondback Moth Plutella xylostella (Lepidoptera: Plutellidae). Curr. Microbiol. 2008, 56, 327–333. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, R.; Alexander, M. Lysis of Soil Fungi by Bacteria. Can. J. Microbiol. 1963, 9, 169–177. [Google Scholar] [CrossRef]
- Bouain, N.; Shahzad, Z.; Rouached, A.; Khan, G.A.; Berthomieu, P.; Abdelly, C.; Poirier, Y.; Rouached, H. Phosphate and zinc transport and signalling in plants: Toward a better understanding of their homeostasis interaction. J. Exp. Bot. 2014, 65, 5725–5741. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saravanan, V.S.; Subramoniam, S.R.; Raj, S.A. Assessing in Vitro Solubilization Potential of Different Zinc Solubilizing Bacterial (ZSB) Isolates. Braz. J. Microbiol. 2004, 35, 121–125. [Google Scholar] [CrossRef]
- Estrada-Bonilla, G.; Baldani, V.L.D.; De Oliveira, D.M.; Urquiaga, S.; Baldani, J.I. Selection of phosphate-solubilizing diazotrophic Herbaspirillum and Burkholderia strains and their effect on rice crop yield and nutrient uptake. Plant Soil 2012, 369, 115–129. [Google Scholar] [CrossRef]
- Abaid-Ullah, M.; Nadeem, M.; Hassan, M.; Ganter, J.; Muhammad, B.; Nawaz, K.; Shah, A.S.; Hafeez, F.Y. Plant Growth Promoting Rhizobacteria: An Alternate Way to Improve Yield and Quality of Wheat (Triticum Aestivum). Int. J. Agric. Biol. 2015, 17, 51–60. [Google Scholar]
- Khanghahi, M.Y.; Ricciuti, P.; Allegretta, I.; Terzano, R.; Crecchio, C. Solubilization of insoluble zinc compounds by zinc solubilizing bacteria (ZSB) and optimization of their growth conditions. Environ. Sci. Pollut. Res. 2018, 25, 25862–25868. [Google Scholar] [CrossRef]
- Joonu, J.; Averal, H.I. Heavy Metal Resistant CZC Genes Identification in Bacillus Cereus, Enterobacter Asburiae and Pseu-domonas Aeruginosa Isolated from BHEL Industry, Tamilnadu. J. Microbiol. Biotechnol. 2016, 5, 27–31. [Google Scholar]
- Wakatsuki, T. Metal oxidoreduction by microbial cells. J. Ind. Microbiol. Biotechnol. 1995, 14, 169–177. [Google Scholar] [CrossRef]
- Chang, H.-B.; Lin, C.-W.; Huang, H.-J. Zinc-induced Cell Death in Rice (Oryza Sativa L.) Roots. Plant Growth Regul. 2005, 46, 261–266. [Google Scholar] [CrossRef]
- Patzer, S.I.; Hantke, K. The ZnuABC high-affinity zinc uptake system and its regulator Zur in Escherichia coli. Mol. Microbiol. 1998, 28, 1199–1210. [Google Scholar] [CrossRef] [PubMed]
- Rfaki, A.; Zennouhi, O.; Nassiri, L.; Ibijbijen, J. Soil Properties Related to the Occurrence of Rock Phosphate-Solubilizing Bacteria in the Rhizosphere Soil of Faba Bean (Vicia faba L.) in Morocco. Soil Syst. 2018, 2, 31. [Google Scholar] [CrossRef] [Green Version]
- Zaidi, A.; Khan, S.; Ahmad, E. Microphos: Principles, Production and Application Strategies. In Phosphate Solubilizing Microorganisms; Springer: Cham, Switzerland, 2014; pp. 1–30. [Google Scholar]
- Arora, N.K.; Tewari, S.; Singh, R. Multifaceted Plant-Associated Microbes and Their Mechanisms Diminish the Concept of Direct and Indirect PGPRs. In Plant Microbe Symbiosis: Fundamentals and Advances; Springer: New Delhi, India, 2013; pp. 411–449. [Google Scholar]
- Kumar, M.; Mishra, S.; Dixit, V.; Agarwal, L.; Chauhan, P.S.; Nautiyal, C.S. Synergistic effect of Pseudomonas putida and Bacillus amyloliquefaciens ameliorates drought stress in chickpea (Cicer arietinum L.). Plant Signal. Behav. 2016, 11, e1071004. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shrivastava, P.; Kumar, R. Soil salinity: A serious environmental issue and plant growth promoting bacteria as one of the tools for its alleviation. Saudi J. Biol. Sci. 2015, 22, 123–131. [Google Scholar] [CrossRef] [PubMed] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Isolates | Strains | Identity | Accession Number |
|---|---|---|---|
| J16 | Pseudomonas moraviensis | 99.77% | MW420981 |
| J143 | Bacillus halotolerans | 99.65% | MW420982 |
| J146 | Enterobacter hormaechei | 99.42% | MW420983 |
| J158 | Pseudomonas frederiksbergensis | 100% | MW420984 |
| Strains | Antibiotic Resistance | NaCl Tolerance | Temperature Tolerance | Siderophores Production | Extracellular Enzymes | Zn Solubilization | |||
|---|---|---|---|---|---|---|---|---|---|
| Proteases | Cellulases | ZnO | Zn3(PO4)2 | ZnCO3 | |||||
| E. coli DH5α | − | 3% | 37 °C | − | − | − | − | − | − |
| P. moraviensis J16 | Amp R, Cm R, Spect R | <5% | 42 °C | +++ | ++ | − | + | − | − |
| B. halotolerans J143 | − | 5% | 42 °C | ++ | ++ | + | − | − | − |
| E. hormaechei J146 | Tet R, Amp R, Cm R | 5% | 42 °C | ++ | − | − | + | − | − |
| P. frederiksbergensis J158 | Amp R, Cm R | <5% | 42 °C | + | − | − | + | − | − |
| Strain | CuO4S·5H2O (µg·mL−1) | CdSO4 (µg·mL−1) | N2NiO8 (µg·mL−1) | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 300 | 500 | 1000 | 1500 | 300 | 500 | 1000 | 1500 | 300 | 500 | 1000 | 1500 | |
| Pseudomonas moraviensis J16 | + | + | + | − | + | − | − | − | + | + | + | − |
| Bacillus halotolerans J143 | + | + | + | − | − | − | − | - | + | + | + | + |
| Enterobacter hormaechei J146 | + | + | + | + | + | + | + | + | + | + | − | − |
| Pseudomonas frederiksbergensis J158 | + | − | − | − | + | − | − | − | + | + | − | − |
| E. coli DH5α | + | + | + | − | + | − | − | − | + | + | − | − |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fahsi, N.; Mahdi, I.; Mesfioui, A.; Biskri, L.; Allaoui, A. Plant Growth-Promoting Rhizobacteria Isolated from the Jujube (Ziziphus lotus) Plant Enhance Wheat Growth, Zn Uptake, and Heavy Metal Tolerance. Agriculture 2021, 11, 316. https://doi.org/10.3390/agriculture11040316
Fahsi N, Mahdi I, Mesfioui A, Biskri L, Allaoui A. Plant Growth-Promoting Rhizobacteria Isolated from the Jujube (Ziziphus lotus) Plant Enhance Wheat Growth, Zn Uptake, and Heavy Metal Tolerance. Agriculture. 2021; 11(4):316. https://doi.org/10.3390/agriculture11040316
Chicago/Turabian StyleFahsi, Nidal, Ismail Mahdi, Abdelhalem Mesfioui, Latefa Biskri, and Abdelmounaaim Allaoui. 2021. "Plant Growth-Promoting Rhizobacteria Isolated from the Jujube (Ziziphus lotus) Plant Enhance Wheat Growth, Zn Uptake, and Heavy Metal Tolerance" Agriculture 11, no. 4: 316. https://doi.org/10.3390/agriculture11040316
APA StyleFahsi, N., Mahdi, I., Mesfioui, A., Biskri, L., & Allaoui, A. (2021). Plant Growth-Promoting Rhizobacteria Isolated from the Jujube (Ziziphus lotus) Plant Enhance Wheat Growth, Zn Uptake, and Heavy Metal Tolerance. Agriculture, 11(4), 316. https://doi.org/10.3390/agriculture11040316