
Genetic Structure and Molecular Variability of Grapevine Fanleaf Virus in Sicily

 ,
,  ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Field Surveys and Samples Collection
2.2. Preliminary Screening by Serological Analysis
2.3. Total RNA Extraction
2.4. Molecular Analyses
2.5. Sequence Analyses
3. Results
3.1. Incidence of GFLV and ArMV Infection in Sicilian Vineyards
3.2. Polymerase Chain Reaction and Sequencing
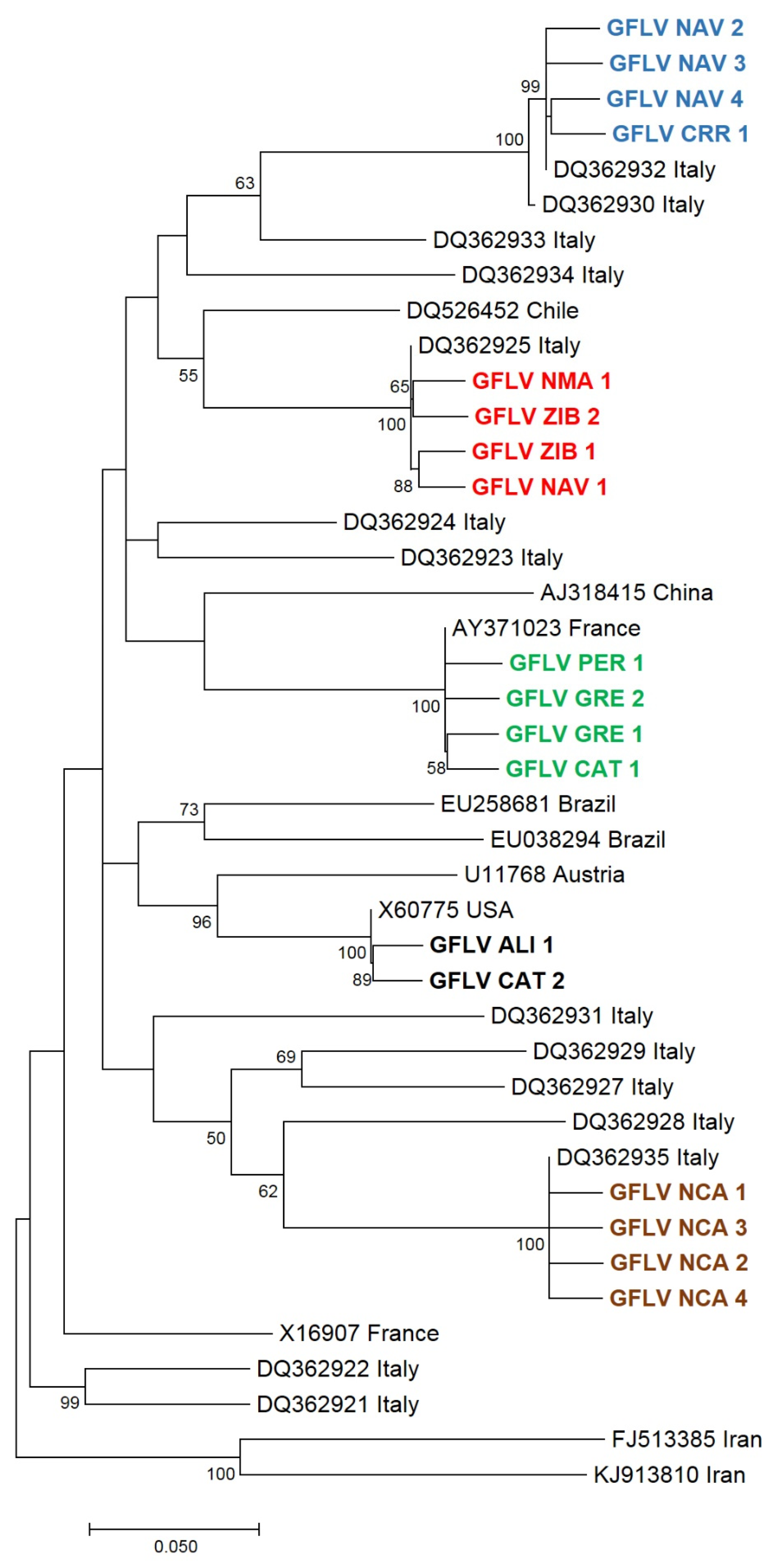
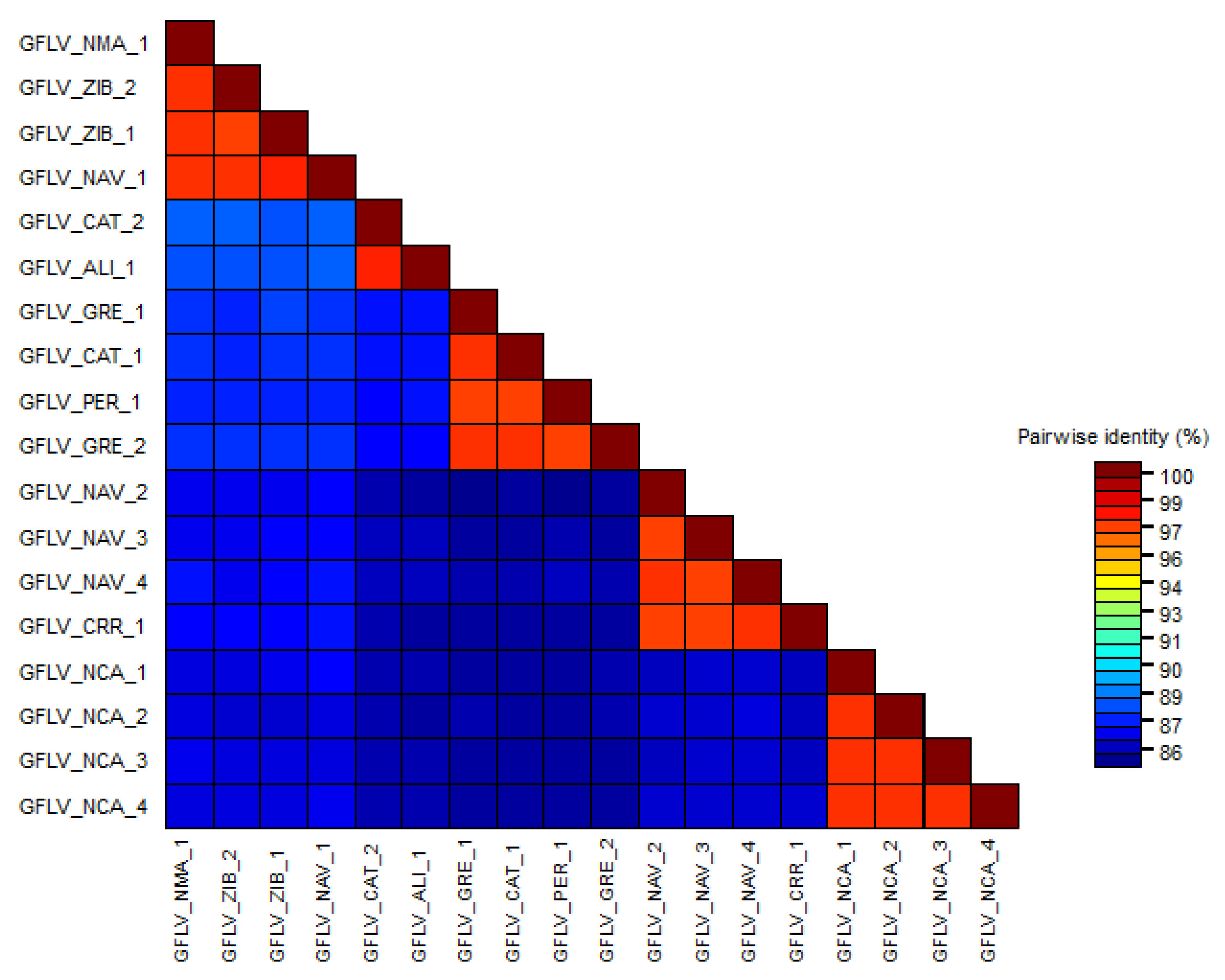
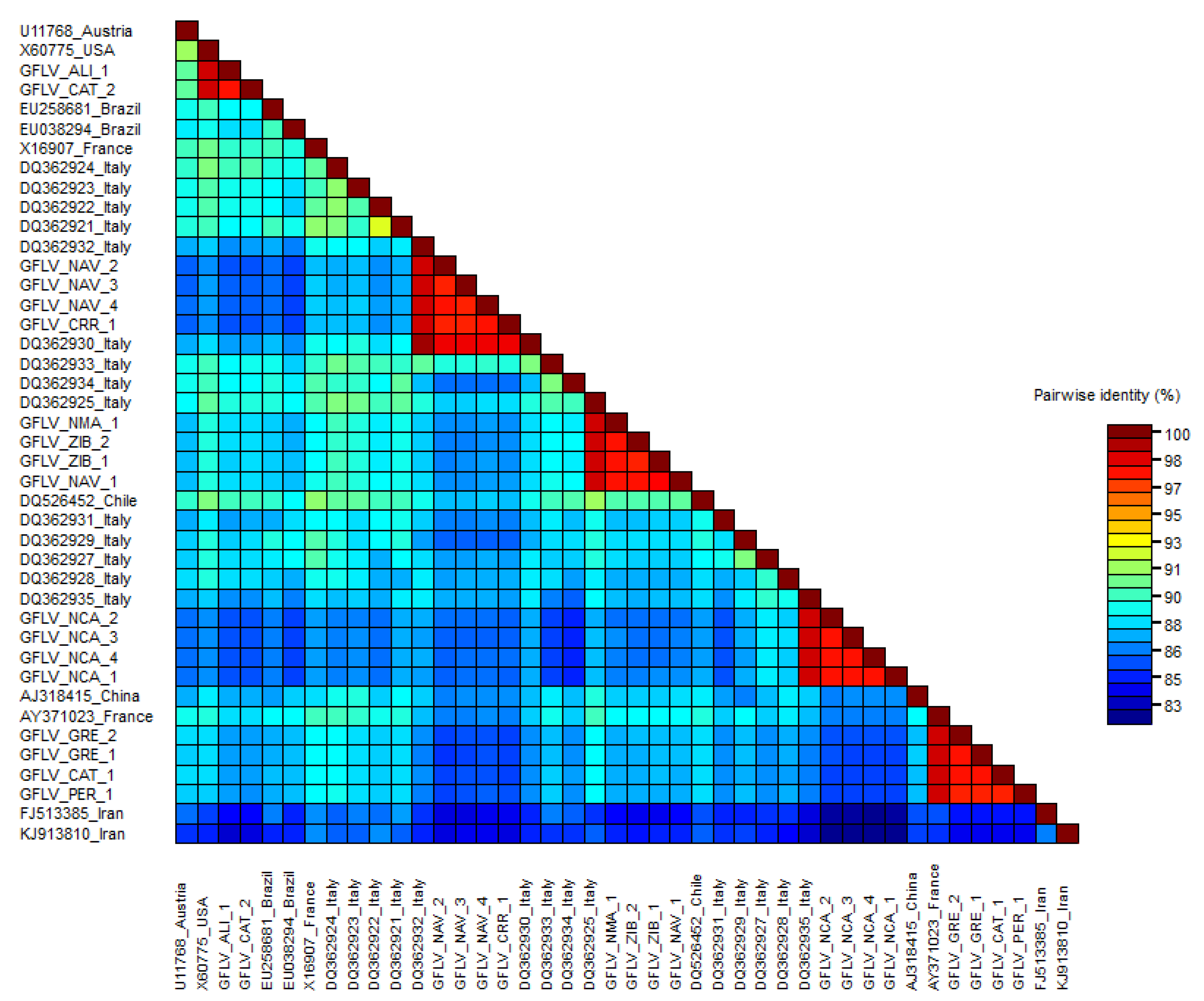
3.3. Phylogenetic Analyses
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Food and Agriculture Organization of the United Nations (FAO). Transforming Our World: The 2030 Agenda for Sustainable Development. Available online: http://www.fao.org/ (accessed on 20 January 2021).
- ISTAT—Istituto Nazionale di Statistica. Available online: www.dati.istat.it (accessed on 1 April 2021).
- Martelli, G.P. Directory of virus and virus-like diseases of the grapevine and their agents. J. Plant Pathol. 2014, 96, 1–136. [Google Scholar]
- Maliogka, V.I.; Martelli, G.P.; Fuchs, M.; Katis, N.I. Control of Viruses Infecting Grapevine. Adv. Virus Res. 2015, 91, 175–227. [Google Scholar] [CrossRef]
- Fuchs, M. Grapevine viruses: A multitude of diverse species with simple but overall poorly adopted management solutions in the vineyard. J. Plant Pathol. 2020, 102, 643–653. [Google Scholar] [CrossRef]
- Panno, S.; Caruso, A.G.; Troiano, E.; Luigi, M.; Manglli, A.; Vatrano, T.; Iacono, G.; Marchione, S.; Bertin, S.; Tomassoli, L.; et al. Emergence of tomato leaf curl New Delhi virus in Italy: Estimation of incidence and genetic diversity. Plant Pathol. 2018, 68, 601–608. [Google Scholar] [CrossRef]
- Sabella, E.; Pierro, R.; Luvisi, A.; Panattoni, A.; D’Onofrio, C.; Scalabrelli, G.; Nutricati, E.; Aprile, A.; De Bellis, L.; Materazzi, A. Phylogenetic analysis of viruses in Tuscan Vitis vinifera sylvestris (Gmeli) Hegi. PLoS ONE 2018, 13, e0200875. [Google Scholar] [CrossRef] [PubMed]
- Martelli, G.P. Grapevine virology highlights: 2010–2012. In Proceedings of the 17th Congress of ICVG, Davis, CA, USA, 7–14 October 2012; pp. 13–31. [Google Scholar]
- Guta, I.-C.; Buciumeanu, E.-C. Grapevine Pinot gris virus infecting grapevines in Romania—Short Communicaiton. Hortic. Sci. 2021, 48, 47–50. [Google Scholar] [CrossRef]
- Martelli, G.P.; Savino, V. Fanleaf degeneration. In Compendium of Grape Diseases; Pearson, R., Goheen, A.C., Eds.; APS Press: Saint Paul, MN, USA, 1990; pp. 48–49. [Google Scholar]
- Oliver, J.E.; Fuchs, M.F. Fanleaf Degeneration/Decline Disease of Grapevines; NYS Integrated Pest Management Program; Cornell University, NYSAES: Geneva, NY, USA, 2011; pp. 1–3. [Google Scholar]
- Basso, M.F.; Fajardo, T.V.M.; Saldarelli, P. Grapevine Virus Diseases: Economic Impact and Current Advances in Viral Prospection and Management. Rev. Bras. Frutic. 2017, 39. [Google Scholar] [CrossRef] [Green Version]
- Andret-Link, P.; Laporte, C.; Valat, L.; Ritzenthaler, C.; Demangeat, G.; Vigne, E.; Fuchs, M. Grapevine fanleaf virus: Still a major threat to the grapevine industry. J. Plant Pathol. 2004, 183–195. [Google Scholar]
- Bertolini, E.; García, J.; Yuste, A.; Olmos, A. High prevalence of viruses in table grape from Spain detected by real-time RT-PCR. Eur. J. Plant Pathol. 2010, 128, 283–287. [Google Scholar] [CrossRef]
- Poljua, D.; Sladonja, B.; Bubola, M. Incidence of viruses infecting grapevine varieties in Istria (Croatia). J. Food Agric. Environ. 2010, 8, 166–169. [Google Scholar]
- Fiore, N.; Zamorano, A.; Rivera, L.; González, F.; Aballay, E.; Montealegre, J.; Pino, A.M. Grapevine Viruses in the Atacama Region of Chile. J. Phytopathol. 2011, 159, 743–750. [Google Scholar] [CrossRef]
- Reynard, J.S.; Gugerli, P. Current status of major grapevine viruses in La Cote vineyards of Switzerland. In Proceedings of the 17th ICVG Congress, Davis, CA, USA, 7–14 October 2012; pp. 7–14. [Google Scholar]
- Mahfoudhi, N.; Harbi-Ben, S.M.; Elair, M.; Selmi, I.; Ben Hamda, H. Prevalence of viruses infecting autochthonous grapevines in Tunisia. Tunis. J. Plant Prot. 2014, 9, 111–118. [Google Scholar]
- Rizzo, D.; Materazzi, A.; Stefani, L.; Farina, P.; Vanarelli, S.; Panattoni, A.; Luvisi, A. Distribution of regulated viruses in cv. Sangiovese vineyards in Tuscany. J. Plant Pathol. 2015, 972, 131–135. [Google Scholar]
- Poojari, S.; Moreau, D.L.; Kahl, D.; Ritchie, M.; Ali, S.; Úrbez-Torres, J.R. Disease incidence and genetic variability of eco-nomically important grapevine viruses in Nova Scotia. Can. J. Plant Pathol. 2020, 42, 584–594. [Google Scholar] [CrossRef]
- Vigne, E.; Bergdoll, M.; Guyader, S.; Fuchs, M. Population structure and genetic variability within isolates of Grapevine fanleaf virus from a naturally infected vineyard in France: Evidence for mixed infection and recombination. J. Gen. Virol. 2004, 85, 2435–2445. [Google Scholar] [CrossRef]
- Sokhandan-Bashir, N.; Melcher, U. Population genetic analysis of grapevine fanleaf virus. Arch. Virol. 2012, 157, 1919–1929. [Google Scholar] [CrossRef]
- Wetzel, T.; Chisholm, J.; Bassler, A.; Sanfaçon, H. Characterization of proteinase cleavage sites in the N-terminal region of the RNA1-encoded polyprotein from Arabis mosaic virus (subgroup A nepovirus). Virology 2008, 375, 159–169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vigne, E.; Marmonier, A.; Fuchs, M. Multiple interspecies recombination events within RNA2 of Grapevine fanleaf virus and Arabis mosaic virus. Arch. Virol. 2008, 153, 1771–1776. [Google Scholar] [CrossRef] [PubMed]
- Zarghani, S.N.; Dupuis-Maguiraga, L.; Bassler, A.; Wetzel, T. Mapping of the exchangeable and dispensable domains of the RNA 2-encoded 2AHP protein of arabis mosaic nepovirus. Virology 2014, 458–459, 106–113. [Google Scholar] [CrossRef] [PubMed]
- Mekuria, T.; Martin, R.R.; Naidu, R.A. First Report of the Occurrence of Grapevine fanleaf virus in Washington State Vineyards. Plant Dis. 2008, 92, 1250. [Google Scholar] [CrossRef]
- Raski, D.J.; Goheen, A.C.; Lider, L.A.; Meredith, C.P. Strategies Against Grapevine Fanleaf Virus and Its Nematode Vector. Plant Dis. 1983, 67, 335. [Google Scholar] [CrossRef]
- Leopold, S.; Borroto-Fernandez, E.; Schartl, A.; Laimer, M. Identification of Xiphinema index in an Austrian vineyard. Vitis 2007, 46, 49–50. [Google Scholar]
- Demangeat, G.; Komar, V.; Van-Ghelder, C.; Voisin, R.; Lemaire, O.; Esmenjaud, D.; Fuchs, M. Transmission Competency of Single-Female Xiphinema index Lines for Grapevine fanleaf virus. Phytopathology 2010, 100, 384–389. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garcia, S.; Hily, J.-M.; Komar, V.; Gertz, C.; Demangeat, G.; Lemaire, O.; Vigne, E. Detection of Multiple Variants of Grapevine Fanleaf Virus in Single Xiphinema index Nematodes. Viruses 2019, 11, 1139. [Google Scholar] [CrossRef] [Green Version]
- Gottwald, T.R.; Hughes, G. A new survey method for Citrus tristeza virus disease assessment. In Proceedings of the XIV International Organization of Citrus Virologists (IOCV), Sao Paulo, Brazil, 14–21 July 2000; pp. 77–87. [Google Scholar]
- Davino, S.; Panno, S.; Arrigo, M.; La Rocca, M.; Caruso, A.G.; Lo Bosco, G. Planthology: An Application System for Plant Diseases Management. Chem. Eng. Trans. 2017, 58, 619–624. [Google Scholar]
- Clark, M.F.; Adams, A.N.; Graham, F.L.; Smiley, J.; Russell, W.C.; Nairn, R. Characteristics of the Microplate Method of Enzyme-Linked Immunosorbent Assay for the Detection of Plant Viruses. J. Gen. Virol. 1977, 34, 475–483. [Google Scholar] [CrossRef]
- Crnogorac, A.; Panno, S.; Mandić, A.; Gašpar, M.; Caruso, A.G.; Noris, E.; Davino, S.; Matić, S. Survey of five major grapevine viruses infecting Blatina and Žilavka cultivars in Bosnia and Herzegovina. PLoS ONE 2021, 16, e0245959. [Google Scholar] [CrossRef]
- Larkin, M.A.; Blackshields, G.; Brown, N.P.; Chenna, R.; Mc Gettigan, P.A.; McWilliam, H.; Valentin, F.; Wallace, I.M.; Wilm, A.; Lopez, R.; et al. ClustalW and Clustal X version 2.0. Bioinformatics 2007, 23, 2947–2948. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nei, M.; Kumar, S. Molecular Evolution and Phylogenetics; Oxford University Press: Oxford, UK, 2000; pp. 147–164. [Google Scholar]
- Efron, B.; Halloran, E.; Holmes, S. Bootstrap confidence levels for phylogenetic trees. Proc. Natl. Acad. Sci. USA 1996, 93, 7085–7090. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Martin, D.P.; Murrell, B.; Golden, M.; Khoosal, A.; Muhire, B. RDP4: Detection and analysis of recombination patterns in virus genomes. Virus Evol. 2015, 1, vev003. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jukes, T.H.; Cantor, C.R. Evolution of Protein Molecules. In Mammalian Protein Metabolism; Munro, H., Ed.; Academic Press: New York, NY, USA, 1969; Volume III, Chapter 24; pp. 21–132. [Google Scholar]
- Pamilo, P.; Bianchi, N. Evolution of the Zfx and Zfy genes: Rates and interdependence between the genes. Mol. Biol. Evol. 1993, 10, 271–281. [Google Scholar] [CrossRef] [Green Version]
- Muhire, B.M.; Varsani, A.; Martin, D.P. SDT: A Virus Classification Tool Based on Pairwise Sequence Alignment and Identity Calculation. PLoS ONE 2014, 9, e108277. [Google Scholar] [CrossRef]
- Davino, S.; Willemsen, A.; Panno, S.; Davino, M.; Catara, A.; Elena, S.F.; Rubio, L. Emergence and Phylodynamics of Citrus tristeza virus in Sicily, Italy. PLoS ONE 2013, 8, e66700. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Panno, S.; Caruso, A.G.; Davino, S. The nucleotide sequence of a recombinant tomato yellow leaf curl virus strain frequently detected in Sicily isolated from tomato plants carrying the Ty-1 resistance gene. Arch. Virol. 2017, 163, 795–797. [Google Scholar] [CrossRef] [PubMed]
- Panno, S.; Caruso, A.; Blanco, G.; Davino, S. First report of Tomato brown rugose fruit virus infecting sweet pepper in Italy. New Dis. Rep. 2020, 41, 20. [Google Scholar] [CrossRef] [Green Version]
- Garcia-Arenal, F.; Fraile, A.; Malpica, J.M. Variability and genetic structure of plant virus populations. Annu. Rev. Phytopathol. 2001, 39, 157–186. [Google Scholar] [CrossRef] [PubMed]
- Marmonier, A.; Schellenberger, P.; Esmenjaud, D.; Schmitt-Keichinger, C.; Ritzenthaler, C.; Andret-Link, P.; Lemaire, O.; Fuchs, M.; Demangeat, G. The coat protein determines the specificity of virus transmission by Xiphinema diversicaudatum. J. Plant Pathol. 2010, 92, 275–279. [Google Scholar]
- Davino, S.; Panno, S.; Iacono, G.; Sabatino, L.; D’Anna, F.; Iapichino, G.; Olmos, A.; Scuderi, G.; Rubio, L.; Tomassoli, L.; et al. Genetic variation and evolutionary analysis of Pepino mosaic virus in Sicily: Insights into the dispersion and epidemiology. Plant Pathol. 2017, 66, 368–375. [Google Scholar] [CrossRef] [Green Version]
- Panno, S.; Caruso, A.G.; Barone, S.; Bosco, G.L.; Rangel, E.A.; Davino, S. Spread of Tomato Brown Rugose Fruit Virus in Sicily and Evaluation of the Spatiotemporal Dispersion in Experimental Conditions. Agronomy 2020, 10, 834. [Google Scholar] [CrossRef]
- Borges, D.F.; Preising, S.; Ambrósio, M.M.D.Q.; da Silva, W.L. Detection of multiple grapevine viruses in New England vineyards. Crop. Prot. 2020, 132, 105143. [Google Scholar] [CrossRef]
- Malenica, N.; Jagić, M.; Pavletić, B.; Bauer, N.; Vončina, D.; Zdunić, G.; Leljak Levanić, D. Somatic embryogenesis as a tool for virus elimination in croatian indigenous grapevine cultivars. Acta Bot. Croat. 2020, 79. [Google Scholar] [CrossRef]
- Rubio, B.; Lalanne-Tisné, G.; Voisin, R.; Tandonnet, J.P.; Portier, U.; Van Ghelder, C.; Lafargue, M.; Petit, J.; Donnart, M.; Joubard, B.; et al. Characterization of genetic determinants of the re-sistance to phylloxera, Daktulosphaira vitifoliae, and the dagger nematode Xiphinema index from muscadine background. BMC Plant Biol. 2020, 20, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, V.C.; Tandonnet, J.-P.; Khallouk, S.; Van Ghelder, C.; Portier, U.; LaFargue, M.-D.-D.; Banora, M.Y.; Ollat, N.; Esmenjaud, D. Grapevine Resistance to the Nematode Xiphinema index Is Durable in Muscadine-Derived Plants Obtained from Hardwood Cuttings but Not from In Vitro. Phytopathology 2020, 110, 1565–1571. [Google Scholar] [CrossRef] [PubMed]
- Pacifico, D.; Caciagli, P.; Palmano, S.; Mannini, F.; Marzachì, C. Quantitation of Grapevine leafroll associated virus-1 and-3, Grapevine virus A, Grapevine fanleaf virus and Grapevine fleck virus in field-collected Vitis vinifera L. ‘Nebbiolo’ by real-time reverse transcription-PCR. J. Virol. Methods 2011, 172, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Čepin, U.; Gutiérrez-Aguirre, I.; Balažic, L.; Pompe-Novak, M.; Gruden, K.; Ravnikar, M. A one-step reverse transcription real-time PCR assay for the detection and quantitation of Grapevine fanleaf virus. J. Virol. Methods 2010, 170, 47–56. [Google Scholar] [CrossRef] [PubMed]
- Panno, S.; Ruiz-Ruiz, S.; Caruso, A.G.; Alfaro-Fernandez, A.; Ambrosio, M.I.F.S.; Davino, S. Real-time reverse transcription polymerase chain reaction development for rapid detection of Tomato brown rugose fruit virus and comparison with other techniques. Peer J. 2019, 7, e7928. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Almasi, M. Establishment and Application of a Reverse Transcription Loop-mediated Isothermal Amplification Assay for Detection of Grapevine Fanleaf Virus. Mol. Biol. 2015, 4. [Google Scholar] [CrossRef]
- Panno, S.; Matić, S.; Tiberini, A.; Caruso, A.G.; Bella, P.; Torta, L.; Stassi, R.; Davino, A.S. Loop Mediated Isothermal Amplification: Principles and Applications in Plant Virology. Plants 2020, 9, 461. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cultivar | No. of Samples Analyzed | ArMV/GFLV Positive Samples |
|---|---|---|
| Grillo | 114 | 0 |
| Nerello mascalese | 43 | 1 |
| Zibibbo | 106 | 7 |
| Nero d’Avola | 64 | 10 |
| Carricante | 30 | 4 |
| Nerello Cappuccio | 24 | 11 |
| Grecanico | 64 | 5 |
| Perricone | 74 | 1 |
| Catarratto | 66 | 8 |
| Alicante | 21 | 1 |
| Moscato | 11 | 0 |
| Total | 617 | 48 |
| Virus | Gene | Primer Name | Sequence (5′-3′) | Amplicon Size (bp) |
|---|---|---|---|---|
| GFLV | Coat protein | EV00N1 * | GACTATCTAGACACATATATACACTTGGGTCTTTTAA | 1573 |
| EV00N3 * | ACTGTCTAGAGGATTRGCYGGYAGAGGAGT | |||
| ArMV | Polyprotein | ArMV 2BF | AGGGTCGCTTCTAGTACAGC | 962 |
| ArMV 2BR | ATCCGAGGAAGAGCAACTCC |
| Cultivar | Number of Samples Analyzed | ArMV | GFLV |
|---|---|---|---|
| No. Positive Samples | No. Positive Samples | ||
| Grillo | 114 | 0 | 0 |
| Nerello mascalese | 1 | 0 | 1 |
| Zibibbo | 7 | 0 | 7 |
| Nero d’Avola | 10 | 0 | 10 |
| Carricante | 4 | 0 | 4 |
| Nerello Cappuccio | 11 | 0 | 11 |
| Grecanico | 5 | 0 | 5 |
| Perricone | 1 | 0 | 1 |
| Catarratto | 8 | 0 | 8 |
| Alicante | 1 | 0 | 1 |
| Moscato | 11 | 0 | 0 |
| Total | 173 | 0 | 48 |
| Cultivar | No. of Samples Collected | % of GFLV Incidence |
|---|---|---|
| Grillo | 114 | - |
| Nerello mascalese | 43 | 2.32 |
| Zibibbo | 106 | 6.60 |
| Nero d’Avola | 64 | 13.33 |
| Carricante | 30 | 13.33 |
| Nerello Cappuccio | 24 | 45.83 |
| Grecanico | 64 | 7.81 |
| Perricone | 74 | 1.35 |
| Catarratto | 66 | 12.12 |
| Alicante | 21 | 4.76 |
| Moscato | 11 | - |
| n b | France | Italy | China | Chile | USA | Austria | Brazil | Iran | |
|---|---|---|---|---|---|---|---|---|---|
| France | 2 | 0.1062±0.500 | 0.1166 ± 0.0188 | 0.1227 ± 0.0614 | 0.1024 ± 0.0521 | 0.1062 ± 0.0535 | 0.1119 ± 0.0560 | 0.1153 ± 0.0579 | 0.1531 ± 0.0768 |
| Italy | 32 | 0.1258±0.008 | 0.1339± 0.0234 | 0.1180 ± 0.0208 | 0.1164 ± 0.0216 | 0.1318± 0.0232 | 0.1354 ± 0.0208 | 0.1726± 0.0263 | |
| China | 1 | n.c. | 0.1238 ± 0.0000 | 0.1246 ± 0.0000 | 0.1372 ± 0.0000 | 0.1360 ± 0.0681 | 0.1593 ± 0.0797 | ||
| Chile | 1 | n.c. | 0.0963 ± 0.0000 | 0.1107 ± 0.0000 | 0.1153 ± 0.0578 | 0.1565 ± 0.0782 | |||
| USA | 1 | n.c. | 0.0881 ± 0.0000 | 0.1123 ± 0.0562 | 0.1651 ± 0.0826 | ||||
| Austria | 1 | n.c. | 0.1207 ± 0.0604 | 0.1561 ± 0.0782 | |||||
| Brazil | 2 | 0.1092±0.500 | 0.1639 ± 0.0822 | ||||||
| Iran | 2 | 0.1476 ± 0.500 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Panno, S.; Caruso, A.G.; Bertacca, S.; Pisciotta, A.; Lorenzo, R.D.; Marchione, S.; Matić, S.; Davino, S. Genetic Structure and Molecular Variability of Grapevine Fanleaf Virus in Sicily. Agriculture 2021, 11, 496. https://doi.org/10.3390/agriculture11060496
Panno S, Caruso AG, Bertacca S, Pisciotta A, Lorenzo RD, Marchione S, Matić S, Davino S. Genetic Structure and Molecular Variability of Grapevine Fanleaf Virus in Sicily. Agriculture. 2021; 11(6):496. https://doi.org/10.3390/agriculture11060496
Chicago/Turabian StylePanno, Stefano, Andrea Giovanni Caruso, Sofia Bertacca, Antonino Pisciotta, Rosario Di Lorenzo, Serafino Marchione, Slavica Matić, and Salvatore Davino. 2021. "Genetic Structure and Molecular Variability of Grapevine Fanleaf Virus in Sicily" Agriculture 11, no. 6: 496. https://doi.org/10.3390/agriculture11060496
APA StylePanno, S., Caruso, A. G., Bertacca, S., Pisciotta, A., Lorenzo, R. D., Marchione, S., Matić, S., & Davino, S. (2021). Genetic Structure and Molecular Variability of Grapevine Fanleaf Virus in Sicily. Agriculture, 11(6), 496. https://doi.org/10.3390/agriculture11060496