
Weed Densities in Perennial Flower Mixtures Cropped for Greater Arable Biodiversity


Abstract
:1. Introduction
2. Materials and Methods
3. Results
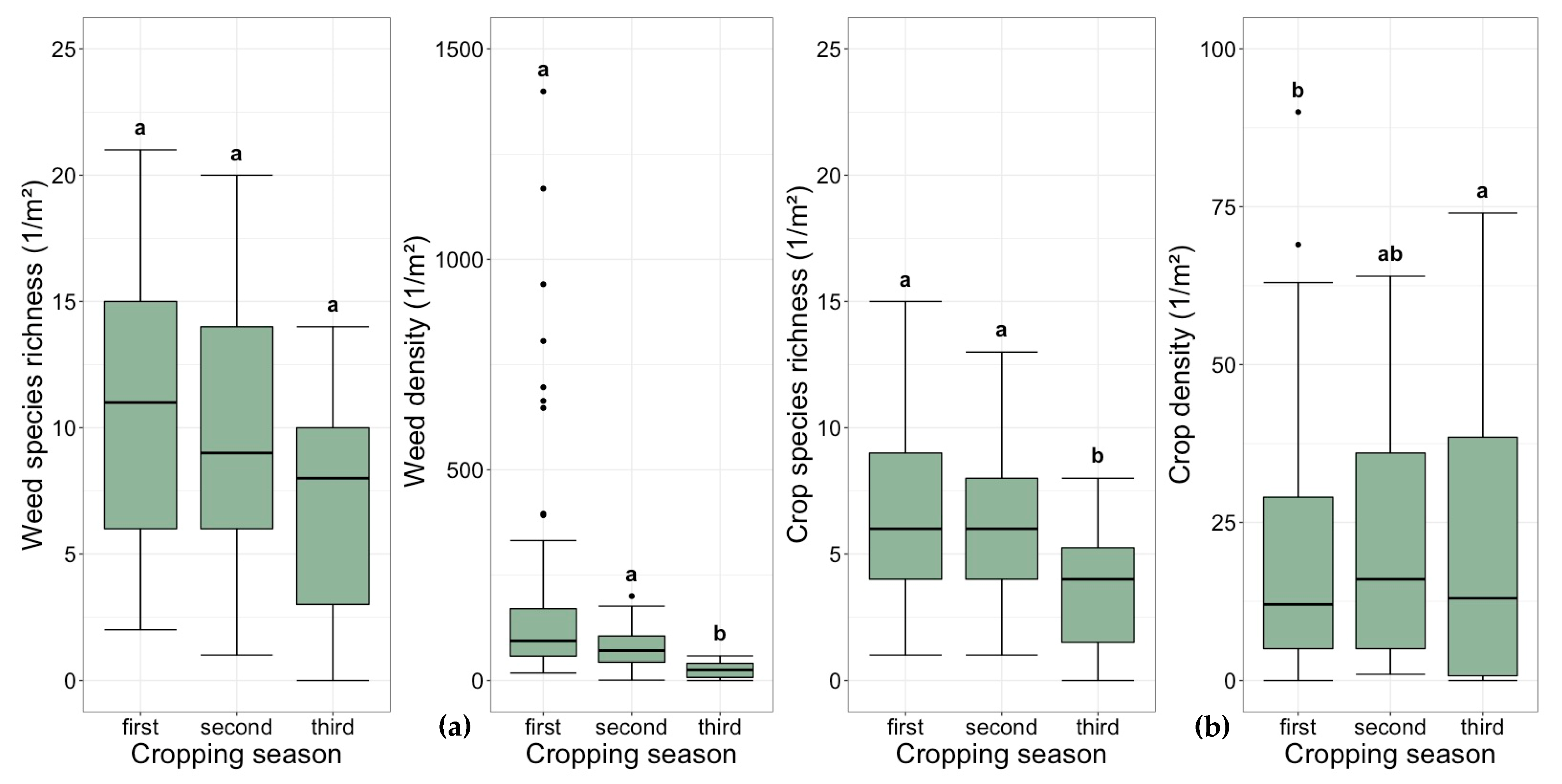
3.1. Weed Species
3.2. Weed Density and Diversity
3.3. Crop Density and Diversity
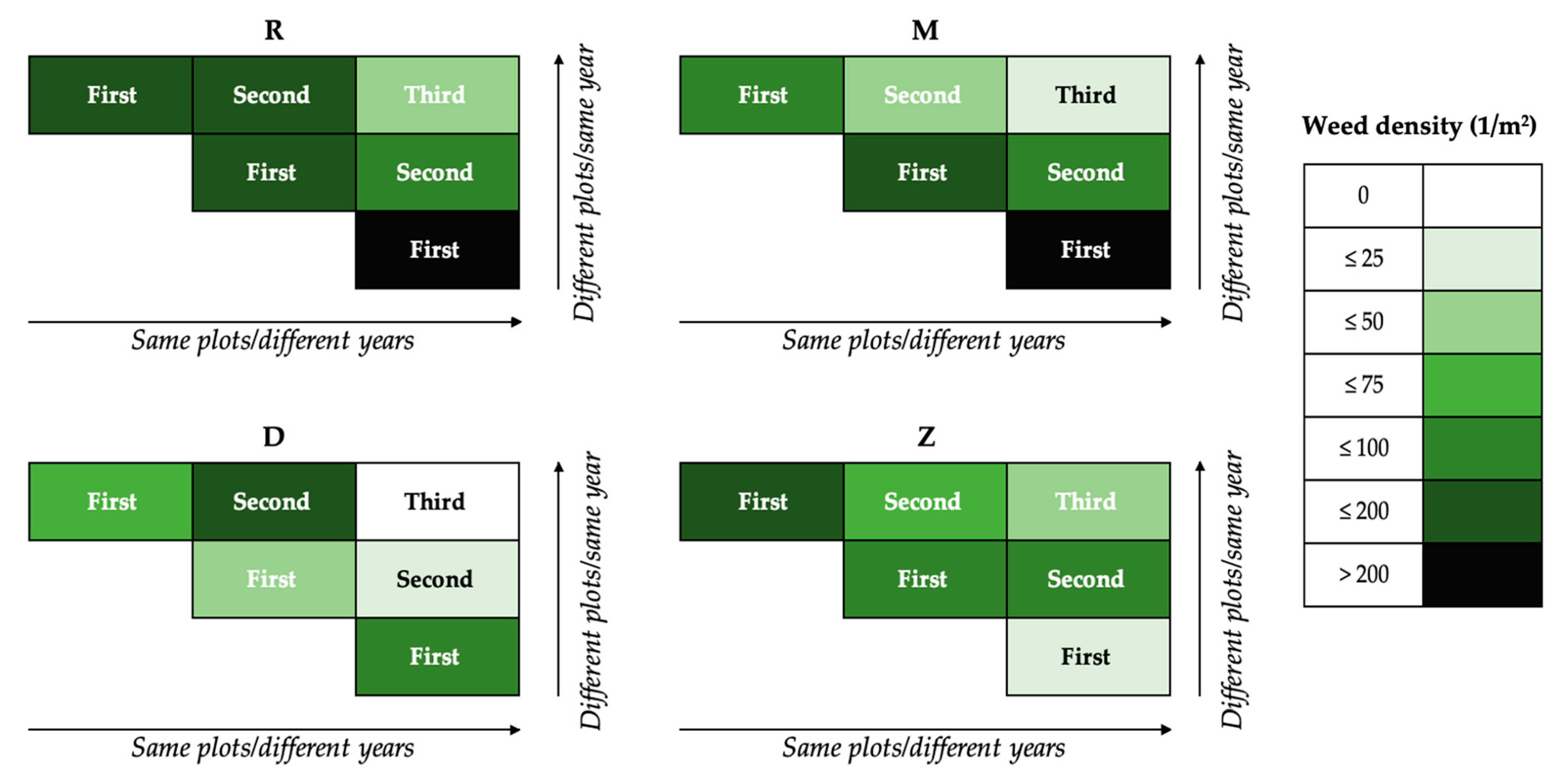
3.4. Three-Year Weed Densities in the Two Set-Ups
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
| Weed Species | First Cropping Season | Second Cropping Season | Third Cropping Season |
|---|---|---|---|
| Achillea millefolium L. | Z | R, Z | Z |
| Agrostis stolonifera L. | M | M | M |
| Amaranthus retroflexus L. | D, Z | Z | |
| Anagallis arvensis L. | M | R | |
| Anchusa arvensis M. Bieb. | M, R, D | ||
| Anthemis arvensis L. | R | ||
| Anthoxanthum odoratum | R | R, Z | R, Z |
| Apera spica-venti Beauv. | R | R, D, Z | Z |
| Aphanes arvensis L. | M, R | R, D | |
| Arabidopsis thaliana Heynh. | M, R | M, R | M, R |
| Arenaria serpyllifolia L. | M | M, Z | M, Z |
| Borago officinalis L. | R | ||
| Brassica napus L. | R, D | M | |
| Brassica nigra W. D. J. Koch | D | ||
| Bromus hordeaceus L. | D | ||
| Capsella bursa-pastoris Medik. | R, D, Z | R, Z | |
| Centaurea cyanus L. | R | M, R, Z | |
| Cerastium arvense L. | R, M | M, R | M, R |
| Cereale ssp. | M, R | R | M |
| Chenopodium album L. | M, R, D, Z | M, R, D, Z | R |
| Cirsium arvense Scop. | M, R, D, Z | M, R | M, R |
| Crepis biennis L. | R | R, Z | |
| Crepis tectorum L. | Z | Z | Z |
| Echinochloa crus-galli P. Beauv. | D, Z | D, Z | |
| Elymus repens Gould | R, D, Z | R, D, Z | R, Z |
| Epilobium parviflorum Schreb. | R | M | M |
| Erigeron canadensis L. | M, D | M, D, Z | M, Z |
| Erophila verna Besser | R | ||
| Euphorbia helioscopia L. | M | ||
| Fallopia convolvulus A. Löve | M, R, D, Z | M, R, D, Z | Z |
| Festuca rubra L. | M, R | ||
| Fumaria officinalis L. | M | M | |
| Galinsoga ssp. | M, R | M | |
| Galium aparine L. | M | M | |
| Geranium pusillum L. | R, D | D, Z | |
| Glechoma hederacea L. | M | ||
| Gnaphalium uliginosum L. | M, R, D | ||
| Hieracium ssp. | R | R, Z | |
| Holcus lanatus L. | M | M, R | M, R |
| Hypericum perforatum L. | M, R | M | |
| Hypochaeris glabra L. | D, Z | ||
| Hypochaeris radicata L. | R | R, Z | R, Z |
| Lactuca serriola L. | M | M, R | M, Z |
| Lamium amplexicaule L. | M, R, D | M | |
| Lamium purpureum L. | M, R | M, R | M |
| Leontodon autumnalis L. | Z | ||
| Lolium perenne L. | R | ||
| Lotus corniculatus L. | R | ||
| Lupinus ssp. | M | M | |
| Matricaria chamomilla L. | R | R, D | R |
| Matricaria discoidea DC. | D | D | |
| Matricaria inodora L. | D, Z | D, Z | Z |
| Medicago lupulina L. | M, R | M, R, Z | R |
| Myosotis sylvatica Ehrh. ex Hoffm. | R | M | |
| Oenothera biennis L. | R, D | Z | Z |
| Papaver rhoeas L. | M, R | M, R, Z | Z |
| Persicaria maculosa Gray | M, R, D | ||
| Plantago lanceolata L. | M, R | M, R | M, R |
| Plantago major L. | M, R, D | M, R, D | |
| Plantago media L. | D | ||
| Poa annua L. | M, R, D | M, R, D | M, R |
| Poa trivialis L. | R, M | M, R, Z | M, R |
| Polygonum aviculare L. | M, R, D, Z | M, D, Z | |
| Polygonum lapathifolium L. | M, R | M, R | |
| Ranunculus repens L. | M | ||
| Rorippa sylvestris Besser | M | ||
| Rumex acetosa L. | M, R, Z | R, Z | R, Z |
| Rumex acetosella L. | Z | Z | |
| Rumex crispus L. | M | M, R, Z | M, R |
| Rumex obtusifolius L. | M, R | M, Z | |
| Secale cereale L. | R | Z | |
| Senecio vulgaris L. | M | M | |
| Setaria viridis P. Beauv. | Z | Z | |
| Sinapis arvensis L. | M | ||
| Sisymbrium officinale Scop. | M | ||
| Solanum nigrum L. | M | ||
| Solidago canadensis L. | Z | Z | |
| Sonchus arvensis L. | M, R | R | |
| Spergula arvensis L. | M, R, D | M, D, Z | |
| Stellaria media Vill. | M, R, D, Z | M, R, Z | M, R |
| Taraxacum officinale F. H. Wigg. | M, R, D, Z | M, R, D, Z | M, R, D |
| Thlaspi arvense L. | M, D, Z | M | |
| Tordylium officinale Reichb. | D | ||
| Trifolium ssp. | M, R, D, Z | M, R, D, Z | M, R, D, Z |
| Urtica dioica L. | M | M | M |
| Veronica arvensis L. | M, R, D | M | M |
| Veronica hederifolia L. | M, R | M, R | R |
| Veronica persica Poir. | M, R | M, R | R |
| Vicia ssp. | M, R, D, Z | M, R, D, Z | M, R, Z |
| Viola arvensis Murray | M, R, D, Z | M, R, D, Z | R |
References
- Lüker-Jans, N.; Simmering, D.; Otte, A. The impact of biogas plants on regional dynamics of permanent grassland and maize area—The example of Hesse, Germany (2005–2010). Agric. Ecosyst. Environ. 2017, 241, 24–38. [Google Scholar] [CrossRef]
- German Maize Committee. Fakten—Statistik—Deutschland-Maisanbauflächen in Deutschland. Available online: https://www.maiskomitee.de/fakten.aspx/statistik/deutschland (accessed on 9 December 2020).
- Meissle, M.; Mouron, P.; Musa, T.; Bigler, F.; Pons, X.; Vasileiadis, V.P.; Otto, S.; Antichi, D.; Kiss, J.; Pálinkás, Z.; et al. Pests, pesticide use and alternative options in European maize production: Current status and future prospects. J. Appl. Entomol. 2010, 134, 357–375. [Google Scholar] [CrossRef]
- Pedroli, B.; Elbersen, B.; Frederiksen, P.; Grandin, U.; Heikkilä, R.; Krogh, P.H.; Izakovičová, Z.; Johansen, A.; Meiresonne, L.; Spijker, J. Is energy cropping in Europe compatible with biodiversity?—Opportunities and threats to biodiversity from land-based production of biomass for bioenergy purposes. Biomass Bioenergy 2010, 55, 73–86. [Google Scholar] [CrossRef]
- von Redwitz, C.; Glemnitz, M.; Hoffmann, J.; Brose, R.; Verch, G.; Barkusky, D.; Bellingrath-Kimura, S. Microsegregation in maize cropping—A chance to improve farmland biodiversity. Gesunde Pflanz. 2019, 71, 87–102. [Google Scholar] [CrossRef]
- Kiesel, A.; Wagner, M.; Lewandowski, I. Environmental performance of Miscanthus, switchgrass and maize: Can C4 perenni-als increase the sustainability of biogas production? Sustainability 2017, 9, 5. [Google Scholar] [CrossRef] [Green Version]
- Von Cossel, M.; Lewandowski, I. Perennial wild plant mixtures for biomass production: Impact of species composition dynamics on yield performance over a five-year cultivation period in southwest Germany. Eur. J. Agron. 2016, 79, 74–89. [Google Scholar] [CrossRef]
- Lewandowski, I. The role of perennial biomass crops in a growing bioeconomy. In Perennial Biomass Crops for a Resource-con-Strained World; Barth, S., Murphy-Bokern, D., Kalinina, O., Taylor, G., Jones, M., Eds.; Springer: Cham, Switzerland, 2016; Volume 319, pp. 3–13. [Google Scholar]
- Börjesson, P. Environmental effects of energy crop cultivation in Swedend: Identification and quantification. Biomass Bioenergy 1999, 16, 137–154. [Google Scholar] [CrossRef]
- Netzwerk Lebensraum Feldflur. Energie aus Wildpflanzen Praxisempfehlung für den Anbau von Wildpflanzen zur Biomasseproduktion. Available online: https://www.saaten-zeller.de/rel/images/biogas/praxisratgeber_wildpflanzen_energie.pdf (accessed on 1 April 2021).
- Emmerling, C. Impact of land-use change towards perennial energy crops on earthworm population. Appl. Soil Ecol. 2014, 84, 12–15. [Google Scholar] [CrossRef]
- Emmerling, C.; Schmidt, A.; Ruf, T.; von Francken-Welz, H.; Thielen, S. Impact of newly introduced perennial bioenergy crops on soil quality parameters at three different locations in W-Germany. J. Plant. Nutr. Soil Sci. 2017, 180, 759–767. [Google Scholar] [CrossRef]
- Zhang, Y.; Li, Y.; Jiang, L.; Tain, C.; Li, J.; Xiao, Z. Potential of perennial crops on environmental sustainability of agriculture. Procedia Environ. Sci. 2011, 10, 1141–1147. [Google Scholar] [CrossRef] [Green Version]
- Haughton, A.J.; Bohan, D.A.; Clark, S.J.; Mallott, M.D.; Mallot, V.; Sage, R.; Karp, A. Dedicated biomass crops can enhance biodiversity in the arable landscape. GCB Bioenergy 2016, 8, 1071–1081. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Werling, B.P.; Dickson, T.L.; Isaacs, R.; Gaines, H.; Gratton, C.; Gross, K.L.; Liere, H.; Malmstrom, C.M.; Meehan, T.D.; Ruan, L.; et al. Perennial grasslands enhance biodiversity and multiple ecosystem services in bioenergy landscapes. Proc. Natl. Acad. Sci. USA 2014, 111, 1652–1657. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Benton, T.G.; Vickery, J.A.; Wilson, J.D. Farmland biodiversity: Is habitat heterogeneity the key? Trends Ecol. Evol. 2003, 18, 182–188. [Google Scholar] [CrossRef]
- Wagner, C.; Schmidt, C. Blühflächen erhöhen die Tierartenvielfalt in der Feldflur. In Bayerische Landesanstalt für Landwirtschaft (LfL) (ed) Wildtiere in der Agrarlandschaft 14; Kulturlandschaftstag: Freising, Germany; Bayerische Landesanstalt für Landwirtschaft (LfL): Freising-Weihenstephan, Germany, 2016; pp. 53–62. [Google Scholar]
- Vollrath, B.; Werner, A.; Degenbeck, M.; Illies, I.; Zeller, J.; Marzini, K. Energetische Verwertung von Kräuterreichen Ansaaten in der Agrarlandschaft und im Siedlungsbereich-eine Ökologische und Wirtschaftliche Alternative bei der Biogasproduktion. Energie aus Wildpflanzen-Schlussbericht. Available online: https://www.lwg.bayern.de/mam/cms06/landespflege/dateien/energie_aus_wildpflanzhen_fnr_abschlussbericht_22005308_in.pdf (accessed on 1 April 2021).
- Uyttenbroeck, R.; Hatt, S.; Piqueray, J.; Paul, A.; Bodson, B.; Francis, F.; Monty, A. Creating perennial flower strips: Think functional! Agric. Agric. Sci. Procedia. 2015, 6, 95–101. [Google Scholar] [CrossRef] [Green Version]
- Albrecht, H. Suitability of arable weeds as indicator organisms to evaluate species conservation effects of management in agricultural ecosystems. Agric. Ecosyst. Environ. 2003, 98, 201–211. [Google Scholar] [CrossRef]
- Marshall, E.J.P.; Brown, V.K.; Boatman, N.D.; Lutman, P.J.W.; Squire, G.R.; Ward, L.K. The role of weeds in supporting biological diversity within crop fields. Weed Res. 2003, 43, 77–89. [Google Scholar] [CrossRef] [Green Version]
- Kulkarni, S.S.; Dosdall, L.M.; Spence, J.R.; Willenborg, C.J. Field density and distribution of weeds are associated with spatial dynamics of omnivorous ground beetles (Coleoptera: Carabidae). Agric. Ecosyst. Environ. 2017, 236, 134–141. [Google Scholar] [CrossRef]
- Schumacher, M.; Dieterich, M.; Gerhards, R. Effects of weed biodiversity on the ecosystem service of weed seed predation along a farming intensity gradient. Glob. Ecol. Conserv. 2020, 24, e01316. [Google Scholar] [CrossRef]
- Vollrath, B.; Marzini, K. Mehr Vielfalt und rentable Biogasproduktion -mit den richtigen Blühmischungen ist beides möglich. LWG 2016, 5, 1–5. [Google Scholar]
- Von Cossel, M.; Steberl, K.; Hartung, J.; Pereira, L.A.; Kiesel, A.; Lewandowski, I. Methane yield and species diversity dynamics of perennial wild plant mixtures established alone, under cover crop maize (Zea mays L.), and after spring barley (Hordeum vulgare L.). Glob. Chang. Biol. Bioenergy 2019, 11, 1376–1391. [Google Scholar] [CrossRef] [Green Version]
- Feledyn-Szewczyk, B.; Matyka, M.; Staniak, M. Comparison of the Effect of Perennial Energy Crops and Agricultural Crops on Weed Flora Diversity. Agronomy 2019, 9, 695. [Google Scholar] [CrossRef] [Green Version]
- Tørresen, K.S.; Skuterud, R.; Tandsaether, H.J.; Hagemo, M.B. Long-term experiments with reduced tillage in spring cereals. I. Effects on weed flora, weed seedbank and grain yield. Crop Prot. 2003, 22, 185–200. [Google Scholar] [CrossRef]
- Wo´zniak, A.; Soroka, M. Biodiversity of weeds in pea cultivated in various tillage systems. Rom. Agric. Res. 2015, 32, 231–237. [Google Scholar]
- Energie aus Wildpflanzen: Biogas-Mischungen für Innovative Landwirte. Available online: https://www.saaten-zeller.de/landwirtschaft/biogas-i#bg (accessed on 5 November 2020).
- R Development Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2020; ISBN 3-900051-07-0. Available online: http://www.R-project.org (accessed on 12 October 2020).
- Bates, D.; Mächler, M.; Bolker, B.; Walker, S. Fitting Linear Mixed-Effects Models Using lme4. J. Stat. Softw. 2015, 67, 1–48. [Google Scholar] [CrossRef]
- Lenth, R. Emmeans: Estimated Marginal Means, aka Least-Squares Means. R Package Version 1.4.8. Available online: https://CRAN.R-project.org/package=emmeans (accessed on 15 December 2020).
- Plückers, C.; Rascher, U.; Scharr, H.; von Gillhaussen, P.; Beierkuhnlein, C.; Temperton, V.M. Sowing different mixtures in dry acidic grassland produced priority effects of varying strength. Acta Oecol. 2013, 53, 110–116. [Google Scholar] [CrossRef]
- Schmidt, A.; Kirmer, A.; Kiehl, K.; Tischew, S. Seed mixture strongly affects species-richness and quality of perennial flower strips on fertile soil. Basic Appl. Ecol. 2019, 42, 62–72. [Google Scholar] [CrossRef]
- Bonin, C.L.; Fidel, R.B.; Banik, C.; Laird, D.A.; Mitchell, R.; Heaton, E.A. Perennial biomass crop establishment, community characteristics, and productivity in the upper US Midwest: Effects of cropping systems seed mixtures and biochar applications. Eur. J. Agron. 2018, 101, 121–128. [Google Scholar] [CrossRef] [Green Version]
- Redwitz, C.; Gerowitt, B. Which factors support the occurrence of Chenopodium album in maize fields in Northern Germany? In Proceedings of the 26th German Conference on Weed Biology and Weed Control, Braunschweig, Germany, 11–14 March 2014; pp. 165–171. [Google Scholar]
- Brust, J.; Claupein, W.; Gerhards, R. Growth and weed suppression ability of common and new cover crops in Germany. J. Crop Prot. 2014, 63, 1–8. [Google Scholar] [CrossRef]
- Williams, T. A Study of the competitive ability of Chenopodium album L. Weed Res. 1964, 4, 283–295. [Google Scholar] [CrossRef]
- Westerman, P.R.; Hofman, A.; Vet, L.E.M.; van der Werf, W. Relative importance of vertebrates and invertebrates in epigeaic weed seed predation in organic cereal filds. Agric. Ecosyst. Environ. 2003, 95, 417–425. [Google Scholar] [CrossRef]
- Meiss, H.; Le Lagadec, L.; Munier-Jolain, N.; Waldhardt, R.; Petit, S. Weed seed predation increases with vegetation cover in perennial forage crops. Agric. Ecosyst. Environ. 2010, 138, 10–16. [Google Scholar] [CrossRef]
- Van der Putten, W.H.; Mortimer, S.R.; Hedlund, K.; Van Dijk, C.; Brown, V.K.; Lepš, J.; Rodriguez-Barrueco, C.; Roy, J.; Diaz Len, T.A.; Gormsen, D.; et al. Plant species diversity as a driver of early succession in abandoned fields: A multi-site approach. Oecologia. 2000, 124, 91–99. [Google Scholar] [CrossRef]
- Froud-Williams, R.J.; Drennan, D.S.H.; Chancellor, R.J. Influence of Cultivation Regime on Weed Floras of Arable Cropping Systems. J. Appl. Ecol. 1983, 20, 187–197. [Google Scholar] [CrossRef]
- Brandsæter, L.O.; Mangerud, K.; Helgheim, M.; Berge, T.W. Control of perennial weeds in spring cereals through stubble cultivation and mouldboard ploughing during autumn or spring. Crop Prot. 2017, 98, 16–23. [Google Scholar] [CrossRef]


| Factor | R | M | D | Z |
|---|---|---|---|---|
| Temperature 1 | 10.5 °C | 9.9 °C | 9.4 °C | 9.9 °C |
| Precipitation 1 | 584 mm | 517 mm | 531 mm | 520 mm |
| Soil type | loamy sand | sandy loam | sandy loam | loamy sand |
| Soil classification 2 | 45 | 56 | 42 | 25 |
| Plot sizes | 30 m2 | 18 m2 | 48 m2 | 64 m2 |

| Annual Species | Perennial Species | ||
| Fagopyron esculentum Moench | 8.0 | Althaea officinalis L. | 5.5 |
| Helianthus annuus L. | 8.0 | Anthemis tinctoria L. | 0.1 |
| Malva verticillata L. | 7.5 | Artemisia vulgaris L. | 1.0 |
| Centaurea nigra L. | 20.0 | ||
| Biennial species | Cichorium intybus L. | 2.0 | |
| Daucus carota L. | 0.1 | Foeniculum vulgare Mill. | 4.0 |
| Dipsacus sylvestris Huds. | 0.5 | Inula helenium L. | 5.0 |
| Echium vulgare L. | 0.5 | Malva alcea L. | 0.6 |
| Melilotus albus Medik. | 3.5 | Malva sylvestris L. | 7.0 |
| Melilotus officinalis Lam. | 7.0 | Medicago sativa L. | 2.0 |
| Reseda luteola L. | 0.3 | Onobrychis viciifolia Scop. | 9.0 |
| Verbascum thapsus L. | 0.5 | Silene dioica Clairv. | 0.2 |
| Tanacetum vulgare L. | 5.0 | ||
| Fixed Effects | Random Effects | ||||
|---|---|---|---|---|---|
| Crop. Seas. | Seed Rate | Exp. Year | Site | Plot (Block, Site) | |
| Weeds | |||||
| Density | <0.001 *** | 0.938 | 1.000 | <0.001 *** | 0.542 |
| Species richness | 0.137 | 0.739 | <0.001 *** | <0.001 *** | 0.049 * |
| Shannon’s H | <0.001 *** | 0.553 | <0.001 *** | <0.001 *** | 1.000 |
| Shannon’s E | <0.001 *** | 0.339 | <0.001 *** | <0.001 *** | 0.721 |
| Berger-Parker index | <0.001 *** | 0.583 | <0.001 *** | <0.001 *** | 1.000 |
| Crops | |||||
| Density | <0.043 * | <0.01 ** | <0.032 * | <0.001 *** | <0.01 ** |
| Species richness | <0.049 * | 0.607 | <0.001 *** | <0.001 *** | <0.01 ** |
| Shannon’s H | <0.026 * | 0.769 | <0.001 *** | <0.001 *** | <0.001 *** |
| Shannon’s E | 0.626 | 0.432 | 0.015 * | <0.01 ** | <0.01 ** |
| Berger-Parker index | 0.209 | 0.920 | <0.001 *** | <0.01 ** | 0.184 |
| First Crop. Seas. | Second Crop. Seas. | Third Crop. Seas. | |
|---|---|---|---|
| Weeds | |||
| Shannon’s H | 1.27 ± 0.281 a | 1.455 ± 0.065 b | 1.77 ± 0.092 c |
| Shannon’s E | 0.53 ± 0.087 a | 0.59 ± 0.026 a | 0.80 ± 0.038 b |
| BPH index | 2.10 ± 0.434 a | 2.50 ± 0.136 b | 3.00 ± 0.194 c |
| Sample size | N = 96 | N = 56 | N = 24 |
| Crops | |||
| Shannon’s H | 1.50 ± 0.199 a | 1.55 ± 0.047 a | 1.42 ± 0.086 a |
| Shannon’s E | 0.78 ± 0.035 a | 0.79 ± 0.018 a | 0.77 ± 0.030 a |
| BPH index | 2.74 ± 0.348 a | 2.77 ± 0.113 a | 2.48 ± 0.185 a |
| Sample size | N = 76 | N = 57 | N = 19 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tamms, L.; de Mol, F.; Glemnitz, M.; Gerowitt, B. Weed Densities in Perennial Flower Mixtures Cropped for Greater Arable Biodiversity. Agriculture 2021, 11, 501. https://doi.org/10.3390/agriculture11060501
Tamms L, de Mol F, Glemnitz M, Gerowitt B. Weed Densities in Perennial Flower Mixtures Cropped for Greater Arable Biodiversity. Agriculture. 2021; 11(6):501. https://doi.org/10.3390/agriculture11060501
Chicago/Turabian StyleTamms, Laura, Friederike de Mol, Michael Glemnitz, and Bärbel Gerowitt. 2021. "Weed Densities in Perennial Flower Mixtures Cropped for Greater Arable Biodiversity" Agriculture 11, no. 6: 501. https://doi.org/10.3390/agriculture11060501
APA StyleTamms, L., de Mol, F., Glemnitz, M., & Gerowitt, B. (2021). Weed Densities in Perennial Flower Mixtures Cropped for Greater Arable Biodiversity. Agriculture, 11(6), 501. https://doi.org/10.3390/agriculture11060501