
Effects of Plant Protection Products on Biochemical Markers in Honey Bees


Abstract
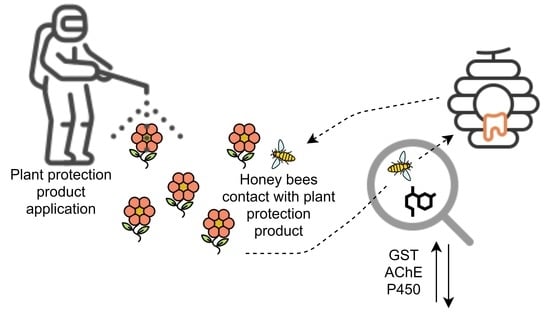
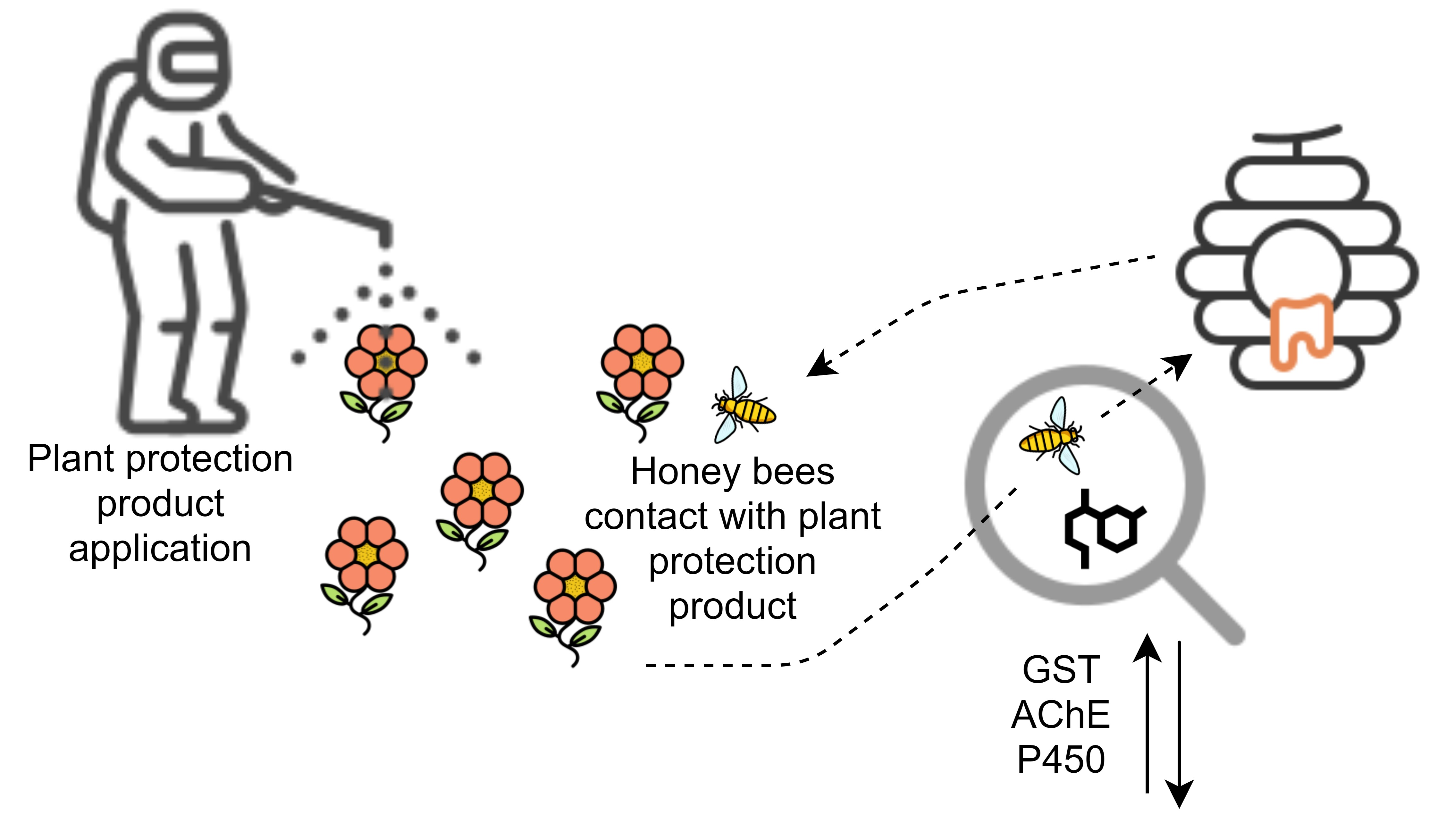
:
1. Introduction
2. Effects of Insecticides Used in Plant Protection on Biochemical Markers in Honey Bees
2.1. Effects of Neonicotinoids on Biochemical Markers in Honey Bees
2.1.1. Introduction
2.1.2. Imidacloprid
Effects of Imidacloprid on Detoxification Enzymes in Honey Bees
Effects of Imidacloprid on Antioxidants and Immune-Related Biochemical Mechanisms in Honey Bees
Effects of Imidacloprid on Immunity, Detoxification, and Antioxidation-Related Genes Expression in Honey Bees
2.1.3. Other Neonicotinoids
Effects of Other Neonicotinoids on Detoxification Enzymes in Honey Bees
Effects of Other Neonicotinoids on Antioxidants and Immune-Related Biochemical Mechanisms in Honey Bees

{kind=link}
| Pesticide | Dose (ng/bee)/Concentration (ppb) | Exposure Type and Duration | Effect on Honey Bee Physiology | Sample for Enzyme Activity Analysis | References | |
|---|---|---|---|---|---|---|
| Neonicotinoids | Imidacloprid(I) | 0.1, 1, and 10 ppb | Orally, 10 and 20 days | ⇿ | Head | [26] |
| 4.3 ng/bee | Orally, 2, 24, and 48 h | ↓ after 48 h | Tissue homogenate | [24] | ||
| 1 ng/bee | Orally, 2, 24, and 48 h | ↓ after 2, 24 h ↑ after 48 h in Apis cerana | Tissue homogenate | [24] | ||
| Thiamethoxam(I) | 300 ppb | Orally, acute | ↑ | Head | [20] | |
| 0.00001, 0.001, and 1.44 ppb | Larvae feeding, 6 days | ↑ | Head | [43] | ||
| 51.16, 5.12, and 2.56 ng/bee | Topically, acute | ⇿ | Head | [42] | ||
| Clothianidin(I) | 1 ng/bee | Orally, 2, 24 and 48 h | ⇿ | Tissue homogenate | [24] | |
| 0.3 ng/bee | Orally, 2, 24 and 48 h | ↓ in Apis cerana | Tissue homogenate | [24] | ||
| Acetamiprid(I) | 3660 pbb | Orally, 1, 5, and 10 days | ⇿ in Apis cerana | Head | [46] | |
| Pyrethroids | Lamda-cyhalothrin(I) | 26,400,000 ppb | Orally, acute | ↑ | Head | [20] |
| 62,200 ppb | Spraying | ↓ | Head and thorax | [30] | ||
| Beta-cyhalothrin(I) | 10,400,000 ppb | Orally, acute | ↓ | Head | [20] | |
| Oxamyl(I) | 68,000 ppb | Spraying | ⇿ | Head and thorax | [30] | |
| Acephate(I) | 168 ppb | Orally, 6, 12, 24, 48, 72 h | ↓ after 48, 72 h | Head and thorax | [44] | |
| 88,300 ppb | Spraying | ↓ | Head and thorax | [30] | ||
| Sulfoxaflor(I) | 58,500 ppb | Spraying | ↑ | Head and thorax | [30] | |
| Abamectin(I) | 33 ppb | Orally, acute | ↓ | Head | [20] | |
| Propiconazole(F) | 24,000 and 60,000 ppb | Orally, 1, 5, and 10 days | ⇿ in Apis cerana | Head | [46] | |
| Difenoconazole(F) | 0.1, 1, and 10 ppb | Orally, 10 and 20 days | ⇿ | Head | [26] | |
| Tetraconazole(F) | 512,500 ppb | Spraying | ⇿ | Head and thorax | [30] | |
| Glyphosate(H) | 0.1, 1, and 10 ppb | Orally, 10 and 20 days | ↑ after 20 days (0.1 ppb) | Head | [26] | |
| 1,217,500 ppb | Spraying | ⇿ | Head and thorax | [30] | ||
| Pesticide | Dose (ng/bee)/Concentration (pbb) | Exposure Type and Duration | Effect on Honey Bee Physiology | Sample for Enzyme Activity Analysis | References | |
|---|---|---|---|---|---|---|
| Neonicotinoids | Imidacloprid(I) | 0.1, 1, and 10 ppb | Orally, 10 and 20 days | ↓ μg/L in abdomen after 10 days (1 ppb) and 20 days | Head, midgut, or abdomen | [26] |
| 968 ppb | Orally, 1 h, 2 h, 4 h, 8 h, 12 h, 16 h, and 20 h. | ⇿ in Apis cerana | Abdomen | [34] | ||
| 4.3 ng/bee | Orally, 2, 24, and 48 h | ↑ after 48 h | Tissue homogenate | [24] | ||
| 1 ng/bee | Orally, 2, 24, and 48 h | ⇿ in Apis cerana | Tissue homogenate | [24] | ||
| 7 ppb | Orally, 8 days | ↑ in queen | Midgut or head | [25] | ||
| Thiamethoxam(I) | 300 ppb | Orally, acute | ↑ | Abdomen | [20] | |
| 0.00001, 0.001, and 1.44 ppb | Larvae feeding, 6 days | ⇿ | Head | [43] | ||
| 51.16, 5.12, and 2.56 ng/bee | Topically, acute | ⇿ | Midgut | [42] | ||
| Clothianidin(I) | 11,000 ppb | Orally, 3 weeks | ↑ | Midgut | [44] | |
| 1 ng/bee | Orally, 2, 24, and 48 h | ⇿ | Tissue homogenate | [24] | ||
| 0.3 ng/bee | Orally, 2, 24, and 48 h | ↑ in Apis cerana after 24 and 48 h | Tissue homogenate | [24] | ||
| Acetamiprid(I) | 3660 ppb | Orally, 1, 5, and 10 days | ↓ after 10 days in Apis cerana | Midgut | [46] | |
| Pyrethroids | Lamda-cyhalothrin(I) | 26,400 ppb | Orally, acute | ↑ | Abdomen | [20] |
| 7300 ppb | Orally, 3 weeks | ↑ | Midgut | [44] | ||
| 62,200 ppb | Spraying | ↓ | Head and thorax | [30] | ||
| Beta-cyhalothrin(I) | 10,400 ppb | Orally, acute | ↓ | Abdomen | [20] | |
| Abamectin(I) | 33,000 ppb | Orally, acute | ↑ | Abdomen | [20] | |
| Oxamyl(I) | 179 ppb | Orally, 3 weeks | ↑ | Midgut | [44] | |
| 68,000 | Spraying | ⇿ | Head and thorax | [30] | ||
| Sulfoxaflor(I) | 58,500 ppb | Spraying | ⇿ | Head and thorax | [30] | |
| Chlorpyrifos(I) | 1860 ppb | Orally, 3 weeks | ⇿ | Midgut | [44] | |
| Acephate(I) | 168 ppb | Orally, 6, 12, 24, 48, and 72 h; 3 weeks | ↓ after 48 and 72 h ⇿after 3 weeks | Midgut | [44] | |
| Tetraconazole(F) | 84 ppb | Orally, 3 weeks | ⇿ | Midgut | [44] | |
| 512,500 ppb | Spraying | ⇿ | Head and thorax | [30] | ||
| Difenoconazole(F) | 0.1, 1 and 10 ppb | Orally, 10 and 20 days | ↓ in midgut after 10 day (1 ppb) | Head, midgut, or abdomen | [26] | |
| Propiconazole(F) | 24,000 ppb | Orally, 1, 5, and 10 days | ⇿in Apis cerana | Midgut | [46] | |
| Glyphosate(H) | 0.1, 1 and 10 ppb | Orally, 10 and 20 days | ↓ in head and abdomen after 10 days and in midgut after 20 days (1 ppb) | Head, midgut, or abdomen | [26] | |
| 35,000 ppb | Orally, 3 weeks | ⇿ | Midgut | [44] | ||
| 1,217,500 ppb | Spraying | ⇿ | Head and thorax | [30] | ||
| Pesticide | Dose (ng/bee)/Concentration (pbb) | Exposure Type and Duration | Effect on Honey bee Physiology | Sample for Enzyme Activity Analisis | References | |
|---|---|---|---|---|---|---|
| Neonicotinoids | Imidacloprid(I) | 968 ppb | Orally, 1 h, 2 h, 4 h, 8 h, 12 h, 16 h, and 20 h. | ↑ after 4 h ↓ after 8–20 h in Apis cerana | Midgut | [34] |
| 39.5 ng/bee | Orally, acute | ↑ 24 and 72 h after application | Bee homogenate | [28] | ||
| 27.7 ng/bee | Topically, acute | ↑ 72 h after aplication | Bee homogenate | [28] | ||
| 4300 ppb | Spraying | ↑ | Head and thorax | [30] | ||
| 0.02 ppb | Topically, acute | ↓ in queens ↓ in workers | Abdomen | [29] | ||
| Clothianidin(I) | 14.7 ng/bee | Orally, acute | ↑ 24 and 72 h after application | Bee homogenate | [28] | |
| 0.63 ng/bee | Topically, acute | ↑ 72 h after aplication | Bee homogenate | [28] | ||
| Acetamiprid(I) | 3660 ppb | Orally, 1, 5, and 10 days | ↑ after 5 days in Apis cerana | Midgut | [46] | |
| Carbaryl(I) | 205 ng/bee | Orally, acute | ↑ 24 and 72 h after application | Bee homogenate | [28] | |
| 112 ng/bee | Topically, acute | ↑ 72 h after aplication | Bee homogenate | [28] | ||
| Acephate(I) | 168 ppb | Orally, 6, 12, 24, 48, and 72 h; 3 weeks | ↓ after 48 ⇿ after 3 weeks | Midgut | [44] | |
| Propiconazole(F) | 2400 ppb | Orally, 1, 5, and 10 days | ⇿ in Apis cerana | Midgut | [46] | |
2.2. Effects of Other Insecticides on Biochemical Markers in Honey Bees
2.2.1. Introduction
2.2.2. Effects of Other Insecticides on Detoxification Enzymes in Honey Bees
2.2.3. Effects of Other Insecticides on Antioxidants and Immune-Related Biochemical Mechanisms in Honey Bees
2.2.4. Effects of Other Insecticides on Immunity, Detoxification, and Antioxidation-Related Gene Expression in Honey Bees
3. Biochemical Effects of Fungicides and Herbicides in Honey Bees
3.1. Introduction
3.2. Fungicides and Herbicides Used in Plant Protection
4. Biochemical Effects of Plant Protection Products Mixtures in Honey Bees
4.1. Introduction
4.2. Mixtures of Plant Protection Products
5. Potential Effects of Pesticides on Honey Bee Biology
5.1. Introduction
5.2. Effects on Detoxification and Antioxidation
5.3. Effects on Immunity
5.4. Effects on Behavior
5.5. Effects on Individual Development
5.6. Effects on Colony Strength
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Bloch, D.; Marx-Stoelting, P.; Martin, S. Towards a tiered test strategy for plant protection products to address mixture toxicity by alternative approaches in human health assessment. Pest. Manag. Sci. 2020, 76, 3326–3332. [Google Scholar] [CrossRef]
- Razaq, M.; Shah, F.M.; Ahmad, S.; Afzal, M. Pest Management for Agronomic Crops. In Agronomic Crops; Springer: Singapore, 2019; pp. 365–384. [Google Scholar]
- Carlile, W.R. Pesticide Selectivity, Health and the Environment; Cambridge University Press: Cambridge, UK, 2006. [Google Scholar]
- Diamand, E. Breaking the pesticide chain. Pestic. Outlook 2003, 14, 153. [Google Scholar] [CrossRef]
- Stanley, D.A.; Russell, A.L.; Morrison, S.J.; Rogers, C.; Raine, N.E. Investigating the impacts of field-realistic exposure to a neonicotinoid pesticide on bumblebee foraging, homing ability and colony growth. J. Appl. Ecol. 2016, 53, 1440–1449. [Google Scholar] [CrossRef]
- Magesh, V.; Zhu, Z.; Tang, T.; Chen, S.; Li, L.; Wang, L. Toxicity of Neonicotinoids to honey bees and detoxification mechanism in honey bees. J. Environ. Sci. Toxicol. Food Technol. 2017, 11, 102–110. [Google Scholar] [CrossRef]
- Migdał, P.; Roman, A.; Popiela-Pleban, E.; Kowalska-Góralska, M.; Opaliński, S. The Impact of Selected Pesticides on Honey Bees. Pol. J. Environ. Stud. 2018, 27, 787–792. [Google Scholar] [CrossRef]
- Decourtye, A.; Devillers, J.; Cluzeau, S.; Charreton, M.; Pham-Delègue, M.-H. Effects of imidacloprid and deltamethrin on associative learning in honeybees under semi-field and laboratory conditions. Ecotoxicol. Environ. Saf. 2004, 57, 410–419. [Google Scholar] [CrossRef]
- Farooqui, T. A potential link among biogenic amines-based pesticides, learning and memory, and colony collapse disorder: A unique hypothesis. Neurochem. Int. 2013, 62, 122–136. [Google Scholar] [CrossRef]
- Paleolog, J.; Wilde, J.; Miszczak, A.; Gancarz, M.; Strachecka, A. Antioxidation Defenses of Apis mellifera Queens and Workers Respond to Imidacloprid in Different Age-Dependent Ways: Old Queens Are Resistant, Foragers Are Not. Anim. Open Access J. MDPI 2021, 11, 1246. [Google Scholar]
- Paleolog, J.; Wilde, J.; Siuda, M.; Bąk, B.; Wójcik, Ł.; Strachecka, A. Imidacloprid markedly affects hemolymph proteolysis, biomarkers, DNA global methylation, and the cuticle proteolytic layer in western honeybees. Apidologie 2020, 51, 620–630. [Google Scholar] [CrossRef] [Green Version]
- Grünewald, B.; Siefert, P. Acetylcholine and Its Receptors in Honeybees: Involvement in Development and Impairments by Neonicotinoids. Insects 2019, 10, 420. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Schuler, M.A.; Berenbaum, M.R. Molecular mechanisms of metabolic resistance to synthetic and natural xenobiotics. Annu. Rev. Entomol. 2007, 52, 231–253. [Google Scholar] [CrossRef]
- Festucci-Buselli, R.A.; Carvalho-Dias, A.S.; Oliveira-Andrade, M.; de Caixeta-Nunes, C.; Li, H.-M.; Stuart, J.J.; Muir, W.; Scharf, M.E.; Pittendrigh, B.R. Expression of Cyp6g1 and Cyp12d1 in DDT resistant and susceptible strains of Drosophila melanogaster. Insect Mol. Biol. 2005, 14, 69–77. [Google Scholar] [CrossRef]
- Mitchell, S.N.; Stevenson, B.J.; Müller, P.; Wilding, C.S.; Egyir-Yawson, A.; Field, S.G.; Hemingway, J.; Paine, M.J.I.; Ranson, H.; Donnelly, M.J. Identification and validation of a gene causing cross-resistance between insecticide classes in Anopheles gambiae from Ghana. Proc. Natl. Acad. Sci. USA 2012, 109, 6147–6152. [Google Scholar] [CrossRef] [Green Version]
- Mao, W.; Schuler, M.A.; Berenbaum, M.R. CYP9Q-mediated detoxification of acaricides in the honey bee (Apis mellifera). Proc. Natl. Acad. Sci. USA 2011, 108, 12657–12662. [Google Scholar] [CrossRef] [Green Version]
- Johnson, R.M.; Wen, Z.; Schuler, M.A.; Berenbaum, M.R. Mediation of Pyrethroid Insecticide Toxicity to Honey Bees (Hymenoptera: Apidae) by Cytochrome P450 Monooxygenases. J. Econ. Entomol. 2006, 99, 1046–1050. [Google Scholar] [CrossRef]
- Abdelkader, F.B.; Guillaume, K.; Tchamitchan, S.; Bonnet, M.; Cousin, M.; Barbouche, N.; Belzunces, L.P.; Brunet, J.L. Effects of clothianidin exposure on semen parameters of honey bee drones. J. New Sci. 2018, 59, 3791–3798. [Google Scholar]
- Abdelkader, F.B.; Kairo, G.; Bonnet, M.; Barbouche, N.; Belzunces, L.P.; Brunet, J.L. Effects of clothianidin on antioxidant enzyme activities and malondialdehyde level in honey bee drone semen. J. Apic. Res. 2019, 58, 740–745. [Google Scholar] [CrossRef]
- Wang, Y.; Zhang, W.; Shi, T.; Xu, S.; Lu, B.; Qin, H.; Yu, L. Synergistic toxicity and physiological impact of thiamethoxam alone or in binary mixtures with three commonly used insecticides on honeybee. Apidologie 2020, 51, 395–405. [Google Scholar] [CrossRef] [Green Version]
- Cerenius, L.; Söderhäll, K. The prophenoloxidase-activating system in invertebrates. Immunol. Rev. 2004, 198, 116–126. [Google Scholar] [CrossRef]
- Zhang, Y.; Chen, D.; Du, M.; Ma, L.; Li, P.; Qin, R.; Yang, J.; Yin, Z.; Wu, X.; Xu, H. Insights into the degradation and toxicity difference mechanism of neonicotinoid pesticides in honeybees by mass spectrometry imaging. Sci. Total Environ. 2021, 774, 145170. [Google Scholar] [CrossRef]
- Simon-Delso, N.; Amaral-Rogers, V.; Belzunces, L.P.; Bonmatin, J.M.; Chagnon, M.; Downs, C.; Furlan, L.; Gibbons, D.W.; Giorio, C.; Girolami, V.; et al. Systemic insecticides (neonicotinoids and fipronil): Trends, uses, mode of action and metabolites. Environ. Sci. Pollut. Res. 2015, 22, 5–34. [Google Scholar] [CrossRef]
- Li, Z.; Li, M.; He, J.; Zhao, X.; Chaimanee, V.; Huang, W.-F.; Nie, H.; Zhao, Y.; Su, S. Differential physiological effects of neonicotinoid insecticides on honey bees: A comparison between Apis mellifera and Apis cerana. Pestic. Biochem. Physiol. 2017, 140, 1–8. [Google Scholar] [CrossRef]
- Dussaubat, C.; Maisonnasse, A.; Crauser, D.; Tchamitchian, S.; Bonnet, M.; Cousin, M.; Kretzschmar, A.; Brunet, J.-L.; Le Conte, Y. Combined neonicotinoid pesticide and parasite stress alter honeybee queens’ physiology and survival. Sci. Rep. 2016, 6, 31430. [Google Scholar] [CrossRef]
- Almasri, H.; Tavares, D.A.; Pioz, M.; Sené, D.; Tchamitchian, S.; Cousin, M.; Brunet, J.-L.; Belzunces, L.P. Mixtures of an insecticide, a fungicide and a herbicide induce high toxicities and systemic physiological disturbances in winter Apis mellifera honey bees. Ecotoxicol. Environ. Saf. 2020, 203, 111013. [Google Scholar] [CrossRef]
- Wilde, J.; Frączek, R.J.; Siuda, M.; Bąk, B.; Hatjina, F.; Miszczak, A. The influence of sublethal doses of imidacloprid on protein content and proteolytic activity in honey bees ( Apis mellifera L.). J. Apic. Res. 2016, 55, 212–220. [Google Scholar] [CrossRef] [Green Version]
- Tarek, H.; Hamiduzzaman, M.M.; Morfin, N.; Guzman-Novoa, E. Research Article Sub-lethal doses of neonicotinoid and carbamate insecticides reduce the lifespan and alter the expression of immune health and detoxification related genes of honey bees (Apis mellifera). Genet. Mol. Res. 2018, 17, 17. [Google Scholar] [CrossRef]
- Chaimanee, V.; Evans, J.D.; Chen, Y.; Jackson, C.; Pettis, J.S. Sperm viability and gene expression in honey bee queens (Apis mellifera) following exposure to the neonicotinoid insecticide imidacloprid and the organophosphate acaricide coumaphos. J. Insect Physiol. 2016, 89, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Zhu, Y.C.; Yao, J.; Adamczyk, J.; Luttrell, R. Synergistic toxicity and physiological impact of imidacloprid alone and binary mixtures with seven representative pesticides on honey bee (Apis mellifera). PLoS ONE 2017, 12, e0176837. [Google Scholar] [CrossRef]
- Manzi, C.; Vergara-Amado, J.; Franco, L.M.; Silva, A.X. The effect of temperature on candidate gene expression in the brain of honey bee Apis mellifera (Hymenoptera: Apidae) workers exposed to neonicotinoid imidacloprid. J. Therm. Biol. 2020, 93, 102696. [Google Scholar] [CrossRef]
- Vergara-Amado, J.; Manzi, C.; Franco, L.M.; Contecha, S.C.; Marquez, S.J.; Solano-Iguaran, J.J.; Haro, R.E.; Silva, A.X. Effects of residual doses of neonicotinoid (imidacloprid) on metabolic rate of queen honey bees Apis mellifera (Hymenoptera: Apidae). Apidologie 2020, 51, 1091–1099. [Google Scholar] [CrossRef]
- Ciereszko, A.; Wilde, J.; Dietrich, G.J.; Siuda, M.; Bąk, B.; Judycka, S.; Karol, H. Sperm parameters of honeybee drones exposed to imidacloprid. Apidologie 2017, 48, 211–222. [Google Scholar] [CrossRef] [Green Version]
- Gao, J.; Jin, S.-S.; He, Y.; Luo, J.-H.; Xu, C.-Q.; Wu, Y.-Y.; Hou, C.-S.; Wang, Q.; Diao, Q.-Y. Physiological Analysis and Transcriptome Analysis of Asian Honey Bee (Apis cerana cerana) in Response to Sublethal Neonicotinoid Imidacloprid. Insects 2020, 11, 753. [Google Scholar] [CrossRef]
- Balieira, K.V.B.; Mazzo, M.; Bizerra, P.F.V.; Guimarães, A.R.d.J.S.; Nicodemo, D.; Mingatto, F.E. Imidacloprid-induced oxidative stress in honey bees and the antioxidant action of caffeine. Apidologie 2018, 49, 562–572. [Google Scholar] [CrossRef] [Green Version]
- Gauthier, M.; Aras, P.; Paquin, J.; Boily, M. Chronic exposure to imidacloprid or thiamethoxam neonicotinoid causes oxidative damages and alters carotenoid-retinoid levels in caged honey bees (Apis mellifera). Sci. Rep. 2018, 8, 16274. [Google Scholar] [CrossRef]
- Christen, V.; Schirrmann, M.; Frey, J.E.; Fent, K. Global Transcriptomic Effects of Environmentally Relevant Concentrations of the Neonicotinoids Clothianidin, Imidacloprid, and Thiamethoxam in the Brain of Honey Bees (Apis mellifera). Environ. Sci. Technol. 2018, 52, 7534–7544. [Google Scholar] [CrossRef] [PubMed]
- Tesovnik, T.; Zorc, M.; Gregorc, A.; Rinehart, T.; Adamczyk, J.; Narat, M. Immune gene expression in developing honey bees ( Apis mellifera L.) simultaneously exposed to imidacloprid and Varroa destructor in laboratory conditions. J. Apic. Res. 2019, 58, 730–739. [Google Scholar] [CrossRef]
- De Smet, L.; Hatjina, F.; Ioannidis, P.; Hamamtzoglou, A.; Schoonvaere, K.; Francis, F.; Meeus, I.; Smagghe, G.; de Graaf, D.C. Stress indicator gene expression profiles, colony dynamics and tissue development of honey bees exposed to sub-lethal doses of imidacloprid in laboratory and field experiments. PLoS ONE 2017, 12, e0171529. [Google Scholar] [CrossRef] [Green Version]
- Gregorc, A.; Alburaki, M.; Rinderer, N.; Sampson, B.; Knight, P.R.; Karim, S.; Adamczyk, J. Effects of coumaphos and imidacloprid on honey bee (Hymenoptera: Apidae) lifespan and antioxidant gene regulations in laboratory experiments. Sci. Rep. 2018, 8, 15003. [Google Scholar] [CrossRef]
- Wu, Y.-Y.; Luo, Q.-H.; Hou, C.-S.; Wang, Q.; Dai, P.-L.; Gao, J.; Liu, Y.-J.; Diao, Q.-Y. Sublethal effects of imidacloprid on targeting muscle and ribosomal protein related genes in the honey bee Apis mellifera L. Sci. Rep. 2017, 7, 15943. [Google Scholar] [CrossRef] [PubMed]
- Badiou-Bénéteau, A.; Carvalho, S.M.; Brunet, J.-L.; Carvalho, G.A.; Buleté, A.; Giroud, B.; Belzunces, L.P. Development of biomarkers of exposure to xenobiotics in the honey bee Apis mellifera: Application to the systemic insecticide thiamethoxam. Ecotoxicol. Environ. Saf. 2012, 82, 22–31. [Google Scholar] [CrossRef] [PubMed]
- Tavares, D.A.; Dussaubat, C.; Kretzschmar, A.; Carvalho, S.M.; Silva-Zacarin, E.C.M.; Malaspina, O.; Bérail, G.; Brunet, J.-L.; Belzunces, L.P. Exposure of larvae to thiamethoxam affects the survival and physiology of the honey bee at post-embryonic stages. Environ. Pollut. 2017, 229, 386–393. [Google Scholar] [CrossRef]
- Yao, J.; Zhu, Y.C.; Adamczyk, J.; Luttrell, R. Influences of acephate and mixtures with other commonly used pesticides on honey bee (Apis mellifera) survival and detoxification enzyme activities. Comp. Biochem. Physiol. Part. C Toxicol. Pharmacol. 2018, 209, 9–17. [Google Scholar] [CrossRef]
- Shi, T.; Burton, S.; Wang, Y.; Xu, S.; Zhang, W.; Yu, L. Metabolomic analysis of honey bee, Apis mellifera L. response to thiacloprid. Pestic. Biochem. Physiol. 2018, 152, 17–23. [Google Scholar] [CrossRef]
- Han, W.; Yang, Y.; Gao, J.; Zhao, D.; Ren, C.; Wang, S.; Zhao, S.; Zhong, Y. Chronic toxicity and biochemical response of Apis cerana cerana (Hymenoptera: Apidae) exposed to acetamiprid and propiconazole alone or combined. Ecotoxicology 2019, 28, 399–411. [Google Scholar] [CrossRef]
- Gajger, I.T.; Sakač, M.; Gregorc, A. Impact of Thiamethoxam on Honey Bee Queen (Apis mellifera carnica) Reproductive Morphology and Physiology. Bull. Environ. Contam Toxicol. 2017, 99, 297–302. [Google Scholar] [CrossRef] [PubMed]
- Ben Abdelkader, F.; Çakmak, İ.; Çakmak, S.S.; Nur, Z.; İncebıyık, E.; Aktar, A.; Erdost, H. Toxicity assessment of chronic exposure to common insecticides and bee medications on colony development and drones sperm parameters. Ecotoxicology 2021, 30, 806–817. [Google Scholar] [CrossRef]
- Annoscia, D.; Di Prisco, G.; Becchimanzi, A.; Caprio, E.; Frizzera, D.; Linguadoca, A.; Nazzi, F.; Pennacchio, F. Neonicotinoid Clothianidin reduces honey bee immune response and contributes to Varroa mite proliferation. Nat. Commun. 2020, 11, 5887. [Google Scholar] [CrossRef] [PubMed]
- Renzi, M.T.; Rodríguez-Gasol, N.; Medrzycki, P.; Porrini, C.; Martini, A.; Burgio, G.; Maini, S.; Sgolastra, F. Combined effect of pollen quality and thiamethoxam on hypopharyngeal gland development and protein content in Apis mellifera. Apidologie 2016, 47, 779–788. [Google Scholar] [CrossRef] [Green Version]
- Domingues, C.E.C.; Abdalla, F.C.; Balsamo, P.J.; Pereira, B.V.R.; Hausen, M.d.A.; Costa, M.J.; Silva-Zacarin, E.C.M. Thiamethoxam and picoxystrobin reduce the survival and overload the hepato-nephrocitic system of the Africanized honeybee. Chemosphere 2017, 186, 994–1005. [Google Scholar] [CrossRef]
- Brandt, A.; Gorenflo, A.; Siede, R.; Meixner, M.; Büchler, R. The neonicotinoids thiacloprid, imidacloprid, and clothianidin affect the immunocompetence of honey bees (Apis mellifera L.). J. Insect Physiol. 2016, 86, 40–47. [Google Scholar] [CrossRef]
- Cook, S.C. Compound and Dose-Dependent Effects of Two Neonicotinoid Pesticides on Honey Bee (Apis mellifera) Metabolic Physiology. Insects 2019, 10, 18. [Google Scholar] [CrossRef] [Green Version]
- Zhu, L.; Qi, S.; Xue, X.; Niu, X.; Wu, L. Nitenpyram disturbs gut microbiota and influences metabolic homeostasis and immunity in honey bee (Apis mellifera L.). Environ. Pollut. 2020, 258, 113671. [Google Scholar] [CrossRef] [PubMed]
- Brandt, A.; Grikscheit, K.; Siede, R.; Grosse, R.; Meixner, M.D.; Büchler, R. Immunosuppression in Honeybee Queens by the Neonicotinoids Thiacloprid and Clothianidin. Sci. Rep. 2017, 7, 4673. [Google Scholar] [CrossRef] [Green Version]
- Meurer-Grimes, B.; Casida, J.E.; Quistad, G.B. Pyrethrum Flowers. Production, Chemistry, Toxicology, and Uses. Brittonia 1996, 48, 613. [Google Scholar] [CrossRef] [Green Version]
- Grdiša, M.; Carović-Stanko, K.; Kolak, I.; Šatović, Z. Morphological and Biochemical Diversity of Dalmatian Pyrethrum (Tanacetum cinerariifolium (Trevir.) Sch. Bip.). Agric. Conspec. Sci. 2009, 74, 73–80. [Google Scholar]
- Naumann, K. Formation of the Pyrethroid-Ester-Linkage. In Synthetic Pyrethroid Insecticides: Chemistry and Patents; Springer: Berlin/Heidelberg, Germany, 1990; pp. 129–143. [Google Scholar]
- Christen, V.; Joho, Y.; Vogel, M.; Fent, K. Transcriptional and physiological effects of the pyrethroid deltamethrin and the organophosphate dimethoate in the brain of honey bees (Apis mellifera). Environ. Pollut. 2019, 244, 247–256. [Google Scholar] [CrossRef]
- Gunasekara, A.S.; Rubin, A.L.; Goh, K.S.; Spurlock, F.C.; Tjeerdema, R.S. Environmental Fate and Toxicology of Carbaryl. In Reviews of Environmental Contamination and Toxicology; Springer: New York, NY, USA, 2008; Volume 196, pp. 95–121. [Google Scholar]
- Di Prisco, G.; Cavaliere, V.; Annoscia, D.; Varricchio, P.; Caprio, E.; Nazzi, F.; Gargiulo, G.; Pennacchio, F. Neonicotinoid clothianidin adversely affects insect immunity and promotes replication of a viral pathogen in honey bees. Proc. Natl. Acad. Sci. USA 2013, 110, 18466–18471. [Google Scholar] [CrossRef] [Green Version]
- Shafiq-ur-Rehman; Rehman, S.; Waliullah, M.I.S. Chlorpyrifos-induced neuro-oxidative damage in bee. Toxicol. Environ. Health Sci. 2012, 4, 30–36. [Google Scholar] [CrossRef]
- Wolstenholme, A.J.; Rogers, A.T. Glutamate-gated chloride channels and the mode of action of the avermectin/milbemycin anthelmintics. Parasitology 2005, 131, S85–S95. [Google Scholar] [CrossRef]
- Pilling, E.D.; Jepson, P.C. Synergism between EBI fungicides and a pyrethroid insecticide in the honeybee (Apis mellifera). Pestic. Sci. 1993, 39, 293–297. [Google Scholar] [CrossRef]
- Łoś, A.; Strachecka, A. Fast and Cost-Effective Biochemical Spectrophotometric Analysis of Solution of Insect “Blood” and Body Surface Elution. Sensors 2018, 18, 1494. [Google Scholar] [CrossRef] [Green Version]
- Zhu, Y.C.; Caren, J.; Reddy, G.V.P.; Li, W.; Yao, J. Effect of age on insecticide susceptibility and enzymatic activities of three detoxification enzymes and one invertase in honey bee workers (Apis mellifera). Comp. Biochem. Physiol. Part. C Toxicol. Pharmacol. 2020, 238, 108844. [Google Scholar] [CrossRef]
- Wilson-Rich, N.; Bonoan, R.E.; Taylor, E.; Lwanga, L.; Starks, P.T. An improved method for testing invertebrate encapsulation response as shown in the honey bee. Insect. Soc. 2019, 66, 91–105. [Google Scholar] [CrossRef]
- Liu, T.; Zhang, L.; Joo, D.; Sun, S.-C. NF-κB signaling in inflammation. Sig. Transduct. Target. 2017, 2, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Strachecka, A.; Olszewski, K.; Paleolog, J. Varroa treatment with bromfenvinphos markedly suppresses honeybee biochemical defence levels. Entomol. Exp. Appl. 2016, 160, 57–71. [Google Scholar] [CrossRef]
- Malone, L.A.; Gatehouse, H.S. Effects ofNosema apisInfection on Honey Bee (Apis mellifera) Digestive Proteolytic Enzyme Activity. J. Invertebr. Pathol. 1998, 71, 169–174. [Google Scholar] [CrossRef]
- Strachecka, A.; Demetraki-Paleolog, J. System proteolityczny powierzchni ciała Apis mellifera w zachowaniu zdrowotności rodzin pszczelich. Kosmos 2011, 60, 43–51. [Google Scholar]
- Asgari, S.; Zhang, G.; Zareie, R.; Schmidt, O. A serine proteinase homolog venom protein from an endoparasitoid wasp inhibits melanization of the host hemolymph. Insect Biochem. Mol. Biol. 2003, 33, 1017–1024. [Google Scholar] [CrossRef]
- Shapira, M.; Thompson, C.K.; Soreq, H.; Robinson, G.E. Changes in Neuronal Acetylcholinesterase Gene Expression and Division of Labor in Honey Bee Colonies. J. Mol. Neurosci. 2001, 17, 1–12. [Google Scholar] [CrossRef]
- Williamson, S.M.; Baker, D.D.; Wright, G.A. Acute exposure to a sublethal dose of imidacloprid and coumaphos enhances olfactory learning and memory in the honeybee Apis mellifera. Invert. Neurosci. 2013, 13, 63–70. [Google Scholar] [CrossRef] [Green Version]
- Gauthier, M.; Belzunces, L.P.; Zaoujal, A.; Colin, M.E.; Richard, D. Modulatory effect of learning and memory on honey bee brain acetylcholinesterase activity. Comp. Biochem. Physiol. Part. C Comp. Pharmacol. 1992, 103, 91–95. [Google Scholar] [CrossRef]
- Navarro-Roldán, M.A.; Amat, C.; Bau, J.; Gemeno, C. Extremely low neonicotinoid doses alter navigation of pest insects along pheromone plumes. Sci. Rep. 2019, 9, 8150. [Google Scholar] [CrossRef] [PubMed]
- Henry, M.; Béguin, M.; Requier, F.; Rollin, O.; Odoux, J.-F.; Aupinel, P.; Aptel, J.; Tchamitchian, S.; Decourtye, A. A common pesticide decreases foraging success and survival in honey bees. Science 2012, 336, 348–350. [Google Scholar] [CrossRef] [PubMed]
- Williamson, S.M.; Moffat, C.; Gomersall, M.A.E.; Saranzewa, N.; Connolly, C.N.; Wright, G.A. Exposure to acetylcholinesterase inhibitors alters the physiology and motor function of honeybees. Front. Physiol. 2013, 4, 13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Evans, J.D.; Aronstein, K.; Chen, Y.P.; Hetru, C.; Imler, J.-L.; Jiang, H.; Kanost, M.; Thompson, G.J.; Zou, Z.; Hultmark, D. Immune pathways and defence mechanisms in honey bees Apis mellifera. Insect Mol. Biol. 2006, 15, 645–656. [Google Scholar] [CrossRef] [Green Version]
- Wessler, I.; Gärtner, H.-A.; Michel-Schmidt, R.; Brochhausen, C.; Schmitz, L.; Anspach, L.; Grünewald, B.; Kirkpatrick, C.J. Honeybees Produce Millimolar Concentrations of Non-Neuronal Acetylcholine for Breeding: Possible Adverse Effects of Neonicotinoids. PLoS ONE 2016, 11, e0156886. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, Y.H.; Kim, J.H.; Kim, K.; Lee, S.H. Expression of acetylcholinesterase 1 is associated with brood rearing status in the honey bee, Apis mellifera. Sci. Rep. 2017, 7, 39864. [Google Scholar] [CrossRef] [Green Version]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Murawska, A.; Migdał, P.; Roman, A. Effects of Plant Protection Products on Biochemical Markers in Honey Bees. Agriculture 2021, 11, 648. https://doi.org/10.3390/agriculture11070648
Murawska A, Migdał P, Roman A. Effects of Plant Protection Products on Biochemical Markers in Honey Bees. Agriculture. 2021; 11(7):648. https://doi.org/10.3390/agriculture11070648
Chicago/Turabian StyleMurawska, Agnieszka, Paweł Migdał, and Adam Roman. 2021. "Effects of Plant Protection Products on Biochemical Markers in Honey Bees" Agriculture 11, no. 7: 648. https://doi.org/10.3390/agriculture11070648
APA StyleMurawska, A., Migdał, P., & Roman, A. (2021). Effects of Plant Protection Products on Biochemical Markers in Honey Bees. Agriculture, 11(7), 648. https://doi.org/10.3390/agriculture11070648