
Neofusicoccum batangarum Causing Dieback of Mango (Mangifera indica) in Florida



{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Pathogen Isolation and DNA Extraction
2.2. Pathogen Identification
2.3. Temperature Growth Response
2.4. Inoculation Study
3. Results
3.1. Pathogen Identification
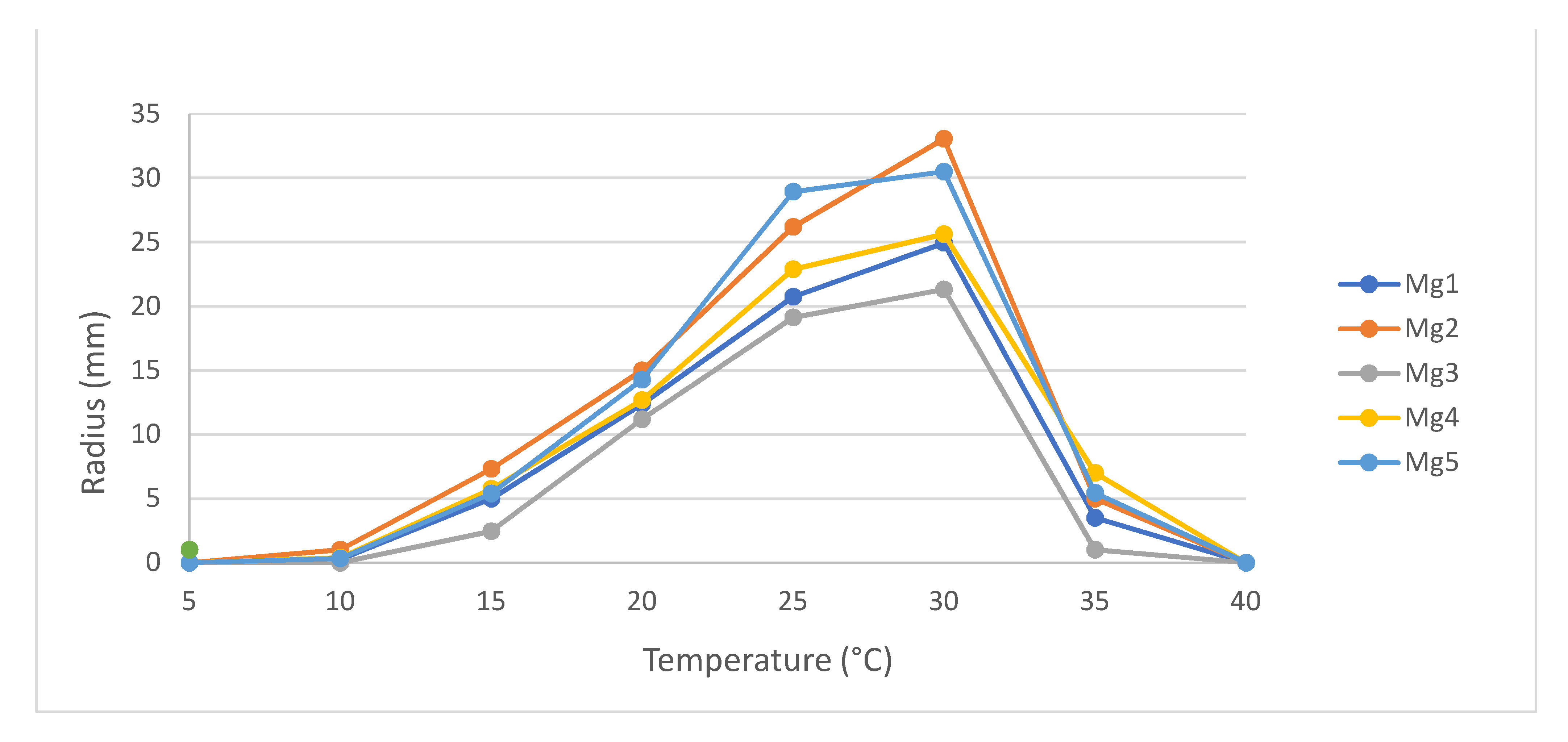
3.2. Temperature Growth Effect
3.3. Inoculation Study
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- De Oliveira Costa, V.S.; Michereff, S.J.; Martins, R.B.; Gava, C.A.T.; Mizubuti, E.S.G.; Câmara, M.P.S. Species of Botryosphaeriaceae associated on mango in Brazil. Eur. J. Plant. Pathol. 2010, 127, 509–519. [Google Scholar] [CrossRef] [Green Version]
- Ablormeti, F.; Coleman, S.; Honger, J.; Owusu, E.; Bedu, I.; Aidoo, O.; Cornelius, E.; Odamtten, G. Management of Lasiodiplodia theobromae, the causal agent of mango tree decline disease in Ghana. Afr. Crop. Sci. J. 2021, 29, 193–207. [Google Scholar] [CrossRef]
- Al Adawi, A.; Deadman, M.; Al Rawahi, A.; Al Maqbali, Y.; Al Jahwari, A.; Al Saadi, B.; Al Amri, I.; Wingfield, M. Aetiology and causal agents of mango sudden decline disease in the Sultanate of Oman. Eur. J. Plant. Pathol. 2006, 116, 247–254. [Google Scholar] [CrossRef]
- Khanzada, M.A.; Lodhi, A.M.; Shahzad, S. Chemical control of Lasiodiplodia theobromae, the causal agent of mango decline in Sindh. Pak. J. Bot. 2005, 37, 1023. [Google Scholar]
- Sakalidis, M.L.; Ray, J.D.; Lanoiselet, V.; Hardy, G.E.S.; Burgess, T.I. Pathogenic Botryosphaeriaceae associated with Mangifera indica in the Kimberley region of Western Australia. Eur. J. Plant. Pathol. 2011, 130, 379–391. [Google Scholar] [CrossRef] [Green Version]
- Sandoval-Sánchez, M.; Nieto-Ángel, D.; Sandoval-Islas, J.S.; Téliz-Ortiz, D.; Orozco-Santos, M.; Silva-Rojas, H. Fungi associated to stem-end rot and dieback of mango (Mangifera indica L.). Agrociencia 2013, 47, 61–73. [Google Scholar]
- Rodríguez-Gálvez, E.; Guerrero, P.; Barradas, C.; Crous, P.W.; Alves, A. Phylogeny and pathogenicity of Lasiodiplodia species associated with dieback of mango in Peru. Fungal Biol. 2017, 121, 452–465. [Google Scholar] [CrossRef] [PubMed]
- Saeed, E.E.; Sham, A.; AbuZarqa, A.; A Al Shurafa, K.; S Al Naqbi, T.; Iratni, R.; El-Tarabily, K.; F AbuQamar, S. Detection and management of mango dieback disease in the United Arab Emirates. Int. J. Mol. Sci. 2017, 18, 2086. [Google Scholar] [CrossRef] [Green Version]
- Crane, J. Tropical fruit production in Florida: Trials, tribulations and opportunities. Proc. Fla. State Hort. Soc 2018. [Google Scholar]
- Ledesma, N. Economic feasibility of small-scale specialty mango production in South Florida. In Proceedings of the Florida State Horticultural Society, Saint Augustine, FL, USA, 31 May–2 June 2015; pp. 25–27. [Google Scholar]
- Ramos, L.J.; Davenport, T.L.; McMillan Jr, R.T.; Lara, S.P. The resistance of mango (Mangifera indica) cultivars to tip dieback disease in Florida. Plant. Dis. 1997, 81, 509–514. [Google Scholar] [CrossRef] [Green Version]
- Ploetz, R.C.; Benscher, D.; Vazquez, A.; Colls, A.; Nagel, J.; Schaffer, B. A reexamination of mango decline in Florida. Plant. Dis. 1996, 80, 664–668. [Google Scholar] [CrossRef]
- Burgess, T.I.; Tan, Y.P.; Garnas, J.; Edwards, J.; Scarlett, K.A.; Shuttleworth, L.A.; Daniel, R.; Dann, E.K.; Parkinson, L.E.; Dinh, Q. Current status of the Botryosphaeriaceae in Australia. Australas. Plant. Pathol. 2019, 48, 35–44. [Google Scholar] [CrossRef]
- Slippers, B.; Johnson, G.I.; Crous, P.W.; Coutinho, T.A.; Wingfield, B.D.; Wingfield, M.J. Phylogenetic and morphological re-evaluation of the Botryosphaeria species causing diseases of Mangifera indica. Mycologia 2005, 97, 99–110. [Google Scholar] [CrossRef]
- Phillips, A.; Alves, A.; Abdollahzadeh, J.; Slippers, B.; Wingfield, M.J.; Groenewald, J.; Crous, P.W. The Botryosphaeriaceae: Genera and species known from culture. Stud. Mycol. 2013, 76, 51–167. [Google Scholar] [CrossRef] [Green Version]
- Sakalidis, M.L.; Hardy, G.E.S.J.; Burgess, T.I. Use of the Genealogical Sorting Index (GSI) to delineate species boundaries in the Neofusicoccum parvum–Neofusicoccum ribis species complex. Mol. Phylogenetics Evol. 2011, 60, 333–344. [Google Scholar] [CrossRef] [Green Version]
- Sakalidis, M.; Slippers, B.; Wingfield, B.D.; Hardy, G.S.J.; Burgess, T. The challenge of understanding the origin, pathways and extent of fungal invasions: Global populations of the Neofusicoccum parvum–N. ribis species complex. Divers. Distrib. 2013, 19, 873–883. [Google Scholar] [CrossRef]
- Begoude, B.D.; Slippers, B.; Wingfield, M.J.; Roux, J. Botryosphaeriaceae associated with Terminalia catappa in Cameroon, South Africa and Madagascar. Mycol. Prog. 2010, 9, 101–123. [Google Scholar] [CrossRef] [Green Version]
- Marques, M.W.; Lima, N.B.; de Morais, M.A.; Michereff, S.J.; Phillips, A.J.; Câmara, M.P. Botryosphaeria, Neofusicoccum, Neoscytalidium and Pseudofusicoccum species associated with mango in Brazil. Fungal Divers. 2013, 61, 195–208. [Google Scholar] [CrossRef]
- Ismail, A.; Cirvilleri, G.; Lombard, L.; Crous, P.; Groenewald, J.; Polizzi, G. Characterisation of Neofusicoccum species causing mango dieback in Italy. J. Plant. Pathol. 2013, 95, 549–557. [Google Scholar]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. PCR Protoc. A Guide Methods Appl. 1990, 18, 315–322. [Google Scholar]
- Glass, N.L.; Donaldson, G.C. Development of primer sets designed for use with the PCR to amplify conserved genes from filamentous ascomycetes. Appl. Environ. Microbiol. 1995, 61, 1323–1330. [Google Scholar] [CrossRef] [Green Version]
- Carbone, I.; Kohn, L.M. A method for designing primer sets for speciation studies in filamentous ascomycetes. Mycologia 1999, 91, 553–556. [Google Scholar] [CrossRef]
- Zhang, W.; Groenewald, J.; Lombard, L.; Schumacher, R.; Phillips, A.; Crous, P. Evaluating species in Botryosphaeriales. Pers.-Mol. Phylogeny Evol. Fungi 2020, 46, 63–115. [Google Scholar]
- Ramos, L.; Lara, S.; McMillan Jr, R.; Narayanan, K. Tip dieback of mango (Mangifera indica) caused by Botryosphaeria ribis. Plant. Dis. 1991, 75, 315–318. [Google Scholar] [CrossRef]
- Rêgo, T.J.; Silva, F.J.; Santos, K.M.; Câmara, M.P.; Correia, K.C.; Michereff, S.J. First report of Neofusicoccum batangarum causing grapevine dieback in Brazil. J. Plant. Pathol. 2020, 102, 973. [Google Scholar] [CrossRef] [Green Version]
- Netto, M.S.; Lima, W.G.; Correia, K.C.; Da Silva, C.F.; Thon, M.; Martins, R.B.; Miller, R.N.; Michereff, S.J.; Câmara, M.P. Analysis of phylogeny, distribution, and pathogenicity of Botryosphaeriaceae species associated with gummosis of Anacardium in Brazil, with a new species of Lasiodiplodia. Fungal Biol. 2017, 121, 437–451. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Diniz, I.; Batista, D.; Pena, A.R.; Rodrigues, A.S.B.; Reis, P.; Baldé, A.; Indjai, B.; Catarino, L.; Monteiro, F. First Report of Dieback Caused by Neofusicoccum batangarum in Cashew in Guinea-Bissau. Plant. Dis. 2021, 105, 1215. [Google Scholar] [CrossRef] [PubMed]
- Serrato-Diaz, L.; Aviles-Noriega, A.; Soto-Bauzó, A.; Rivera-Vargas, L.; Goenaga, R.; Bayman, P. Botryosphaeriaceae fungi as causal agents of dieback and corky bark in rambutan and longan. Plant. Dis. 2020, 104, 105–115. [Google Scholar] [CrossRef] [PubMed]
- Francesco, A.; Giambra, S.; Schena, L.; Surico, G.; Antonella, P.; Gusella, G.; Stracquadanio, C.; Burruano, S.; Cacciola, S.O. New insights into scabby canker of Opuntia ficus-indica, caused by Neofusicoccum batangarum. Phytopathol. Mediterr. 2020, 59, 269–284. [Google Scholar]
- Shetty, K.G.; Minnis, A.M.; Rossman, A.Y.; Jayachandran, K. The Brazilian peppertree seed-borne pathogen, Neofusicoccum batangarum, a potential biocontrol agent. Biol. Control. 2011, 56, 91–97. [Google Scholar] [CrossRef]
- Arjona-Girona, I.; López-Herrera, C. First report of branch dieback in mango trees caused by Neofusicoccum parvum in Spain. Plant. Dis. 2016, 100, 2529. [Google Scholar] [CrossRef]
- U.S. Climate Data. Available online: https://www.usclimatedata.com (accessed on 5 August 2021).
- Zhou, Y.; Gong, G.; Cui, Y.; Zhang, D.; Chang, X.; Hu, R.; Liu, N.; Sun, X. Identification of Botryosphaeriaceae species causing kiwifruit rot in Sichuan Province, China. Plant. Dis. 2015, 99, 699–708. [Google Scholar] [CrossRef] [PubMed]
- Úrbez-Torres, J.; Battany, M.; Bettiga, L.; Gispert, C.; McGourty, G.; Roncoroni, J.; Smith, R.; Verdegaal, P.; Gubler, W. Botryosphaeriaceae species spore-trapping studies in California vineyards. Plant. Dis. 2010, 94, 717–724. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amponsah, N.; Jones, E.; Ridgway, H.; Jaspers, M. Rainwater dispersal of Botryosphaeria conidia from infected grapevines. N. Z. Plant. Prot. 2009, 62, 228–233. [Google Scholar] [CrossRef] [Green Version]
- Amponsah, N.T.; Jones, E.E.; Ridgway, H.J.; Jaspers, M.V. Identification, potential inoculum sources and pathogenicity of botryosphaeriaceous species associated with grapevine dieback disease in New Zealand. Eur. J. Plant. Pathol. 2011, 131, 467–482. [Google Scholar] [CrossRef]
- Amponsah, N.T.; Jones, E.E.; Ridgway, H.J.; Jaspers, M.V. Susceptibility of grapevine tissues to Neofusicoccum luteum conidial infection. Plant. Pathol. 2012, 61, 719–729. [Google Scholar] [CrossRef]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Puig, A.S.; Winterstein, M.C. Neofusicoccum batangarum Causing Dieback of Mango (Mangifera indica) in Florida. Agriculture 2021, 11, 853. https://doi.org/10.3390/agriculture11090853
Puig AS, Winterstein MC. Neofusicoccum batangarum Causing Dieback of Mango (Mangifera indica) in Florida. Agriculture. 2021; 11(9):853. https://doi.org/10.3390/agriculture11090853
Chicago/Turabian StylePuig, Alina S., and Mike C. Winterstein. 2021. "Neofusicoccum batangarum Causing Dieback of Mango (Mangifera indica) in Florida" Agriculture 11, no. 9: 853. https://doi.org/10.3390/agriculture11090853
APA StylePuig, A. S., & Winterstein, M. C. (2021). Neofusicoccum batangarum Causing Dieback of Mango (Mangifera indica) in Florida. Agriculture, 11(9), 853. https://doi.org/10.3390/agriculture11090853