
Virulence Diversity of Puccinia striiformis f. sp. Tritici in Common Wheat in Russian Regions in 2019–2021



Abstract
:1. Introduction
2. Materials and Methods
2.1. Yellow Rust Sampling and Spore Multiplication
2.2. Virulence Characterization
2.3. Data Analysis
3. Results
3.1. Virulence Characterization
3.2. Pathotypes Composition
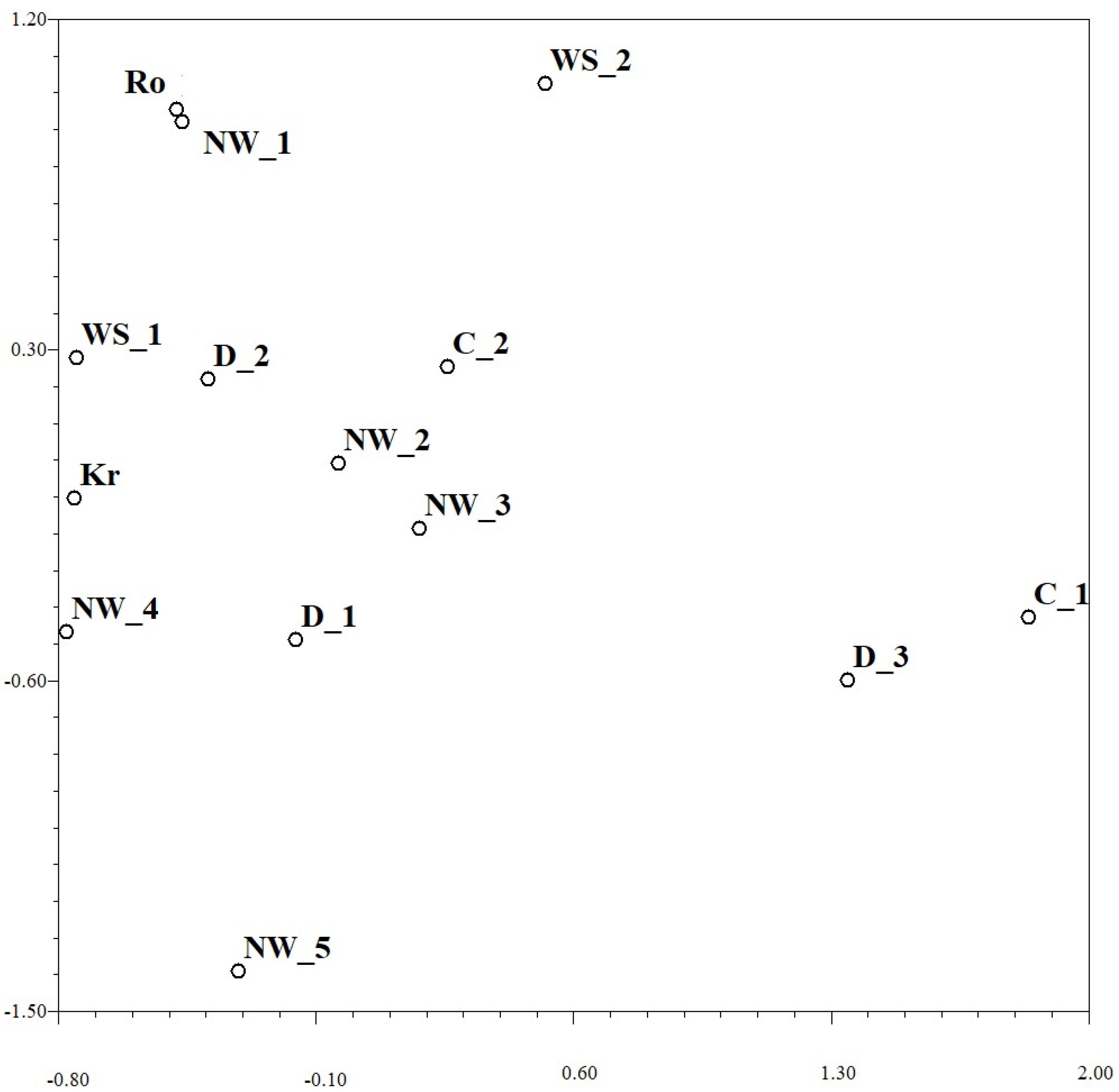
3.3. Variability within and among Regional Collections of Pst Pathotypes
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Listman, M.; Ordóñez, R. Imagine a World without Maize and Wheat. CIMMYT Newsletter World Food Day 2019. Available online: https://www.cimmyt.org/news/ten-things-you-should-know-about-maize-and-wheat/ (accessed on 25 August 2022).
- Samborski, D.J. Wheat Leaf Rust. In The Cereal Rusts; Roelfs, A.P., Bushnell, W.R., Eds.; Volume II Diseases, Distribution, Epidemiology, and Control; Academic Press, Inc. (Harcourt Brace Jovanovich, Publishers): Orlando, FL, USA; San Diego, CA, USA; New York, NY, USA; London, UK; Toronto, ON, Canada; Montreal, QC, Canada; Sydney, NSW, Australia; Tokyo, Japan, 1985. [Google Scholar]
- Wellings, C.R. Global status of stripe rust: A review of historical and current threats. Euphytica 2011, 179, 129–141. [Google Scholar] [CrossRef]
- Chen, W.; Wellings, C.; Chen, X.; Kang, Z.; Liu, T. Wheat stripe (yellow) rust caused by Puccinia striiformis f. sp. tritici. Mol. Plant Pathol. 2014, 15, 433–446. [Google Scholar] [CrossRef]
- Cheng, P.; Chen, X.M.; See, D.R. Grass hosts harbor more diverse isolates of Puccinia striiformis than cereal crops. Phytopathology 2016, 106, 362–371. [Google Scholar] [CrossRef] [Green Version]
- Jin, Y.; Szabo, L.J.; Carson, M. Century-old mystery of Puccinia striiformis life history solved with the identification of Berberis as an alternate host. Phytopathology 2010, 100, 432–435. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, M.N.; Chen, X.M. First report of Oregon grape (Mahonia aquifolium) as an alternate host for the wheat stripe rust pathogen (Puccinia striiformis f. sp. tritici) under artificial inoculation. Plant Dis. 2013, 97, 839. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Zhang, H.; Yao, J.; Huang, L.; Kang, Z. Confirmation of Berberis spp. as alternate hosts of Puccinia striiformis f. sp. tritici on wheat in China. Mycosystema 2011, 30, 895–900. [Google Scholar]
- Zhao, J.; Wang, L.; Wang, Z.; Chen, X.; Zhang, H.; Yao, J.; Zhan, G.; Chen, W.; Huang, L.; Kang, Z. Identification of eighteen Berberis species as alternate hosts of Puccinia striiformis f. sp. tritici and virulence variation in the pathogen isolates from natural infection of barberry plants in China. Phytopathology 2013, 103, 927–934. [Google Scholar] [CrossRef] [Green Version]
- Hovmøller, M.S.; Sørensen, C.K.; Walter, S.; Justesen, A.F. Diversity of Puccinia striiformis on cereals and grasses. Annu. Rev. Phytopathol. 2011, 49, 197–217. [Google Scholar] [CrossRef]
- Sinha, P.; Chen, X. Potential infection risks of the wheat stripe rust and stem rust pathogens on Barberry in Asia and Southeastern Europe. Plants 2021, 10, 957. [Google Scholar] [CrossRef]
- Milus, E.A.; Kristensen, K.; Hovmøller, M.S. Evidence for increased aggressiveness in a recent widespread strain of Puccina striiformis f. sp. tritici causing stripe rust of wheat. Phytopathology 2009, 99, 89–94. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, X.M. Epidemiology and control of stripe rust [Puccinia striiformis f. sp. tritici] on wheat. Can. J. Plant Pathol. 2005, 27, 314–337. [Google Scholar] [CrossRef]
- Koyshibaev, M. About the international conference on yellow rust. Mikol. Fitopatol. 2002, 36, 83–85. (In Russian) [Google Scholar]
- Chen, X.M. Integration of cultivar resistance and fungicide application for control of wheat stripe rust. Can. J. Plant Pathol. 2014, 36, 311–326. [Google Scholar] [CrossRef]
- Chen, X.; Wang, M.; Wan, A.; Bai, Q.; Li, M.; López, P.F.; Maccaferri, M.; Mastrangelo, M.; Barnes, C.; Fabricio, D.; et al. Virulence characterization of Puccinia striiformis f. sp. tritici collections from six countries in 2013 to 2020. Can. J. Plant Pathol. 2021, 43, 308–322. [Google Scholar] [CrossRef]
- Brar, G.S.; Kutcher, H.R. Race characterization of Puccinia striiformis f. sp. tritici, the cause of wheat stripe rust, in Saskatchewan and Southern Alberta, Canada and virulence comparison with races from the United States. Plant Dis. 2016, 100, 1744–1753. [Google Scholar] [CrossRef] [Green Version]
- Wellings, C.R. Puccinia striiformis in Australia: A review of the incursion, evolution and adaptation of stripe rust in the period 1979–2006. Aust. J. Agric. Res. 2007, 58, 567–575. [Google Scholar] [CrossRef]
- Zhou, X.; Fang, T.; Li, K.; Huang, K.; Ma, C.; Zhang, M.; Li, X.; Yang, S.; Ren, R.; Zhang, P. Yield losses associated with different levels of stripe rust resistance of commercial wheat cultivars in China. Phytopathology 2022, 112, 1244–1254. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.M.; Moore, M.; Milus, E.A.; Long, D.L.; Line, R.F.; Marshall, D.; Jackson, L. Wheat stripe rust epidemics and races of Puccinia striiformis f.sp. tritici in the United States in 2000. Plant Dis. 2002, 86, 39–46. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ali, S.; Rodriguez-Algaba, J.; Thach, T.; Sørensen, C.K.; Hansen, J.G.; Lassen, P.; Nazari, K.; Hodson, D.P.; Justesen, A.F.; Hovmøller, M.S. Yellow rust epidemics Worldwide were caused by pathogen races from divergent genetic lineages. Front. Plant Sci. 2017, 8, 1057. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walter, S.; Ali, S.; Kemen, E.; Nazari, K.; Bahri, B.A.; Enjalbert, J.; Hansen, J.G.; Brown, J.K.M.; Sicheritz-Pontén, T.; Jones, J. Molecular markers for tracking the origin and worldwide distribution of invasive strains of Puccinia striiformis. Ecol. Evol. 2016, 6, 2790–2804. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hovmøller, M.S.; Walter, S.; Bayles, R.A.; Hubbard, A.; Flath, K.; Sommerfeldt, N.; Leconte, M.; Czembor, P.; Rodriguez-Algaba, J.; Thach, T.; et al. Replacement of the European wheat yellow rust population by new races from the centre of diversity in the near-Himalayan region. Plant Pathol. 2016, 65, 402–411. [Google Scholar] [CrossRef] [Green Version]
- Hubbard, A.; Lewis, C.; Yoshida, K.; Ramirez-Gonzalez, R.; de Vallavieille-Pope, C.; Thomas, J.; Kamoun, S.; Bayles, R.; Uauy, C.; Saunders, D.G. Field pathogenomics reveals the emergence of a diverse wheat yellow rust population. Genome Biol. 2015, 16, 23. [Google Scholar] [CrossRef] [PubMed]
- Bouvet, L.; Holdgate, S.; James, L.; Thomas, J.; Mackay, I.J.; Cockram, J. The evolving battle between yellow rust and wheat: Implications for global food security. Theor. Appl. Genet. 2022, 135, 741–753. [Google Scholar] [CrossRef]
- Hovmøller, M.S.; Patpour, M.; Rodriguez-Algaba, J.; Thach, T.; Sørensen, C.K.; Justesen, A.F.; Hansen, J.G. GRRC Report of Yellow and Stem Rust Genotyping and Race Analyses 2021. Available online: https://agro.au.dk/fileadmin/www.grcc.au.dk/International_Services/Pathotype_YR_results/GRRC_Annual_Report2021.pdf (accessed on 25 August 2022).
- Sharma-Poudyal, D.; Chen, X.M.; Wan, A.M.; Zhan, G.M.; Kang, Z.S.; Cao, S.Q.; Jin, S.L.; Morgounov, A.; Akin, B.; Mert, Z.; et al. Virulence characterization of international collections of the wheat stripe rust pathogen, Puccinia striiformis f. sp. tritici. Plant Dis. 2013, 97, 379–386. [Google Scholar] [CrossRef] [Green Version]
- Sanin, S.S. Agricultural plant disease control—The main factor of the crop production intensification. Plant Prot. News 2010, 1, 3–14. (In Russian) [Google Scholar]
- Ivanova, Y.N.; Rosenfread, K.K.; Stasyuk, A.I.; Skolotneva, E.S.; Silkova, O.G. Raise and characterization of a bread wheat hybrid line (Tulaykovskaya 10 × Saratovskaya 29) with chromosome 6Agi2 introgressed from Thinopyrum intermedium. Vavilov J. Genet. Breed. 2021, 25, 7701712. [Google Scholar] [CrossRef]
- Zeleneva, Y.V.; Shaydayuk, E.L. Resistance of winter soft wheat varieties to leaf and yellow rust in the conditions of the Central Chernozem region of Russia. E3S Web Conf. 2021, 254, 01017. [Google Scholar] [CrossRef]
- Druzhin, A.E. Effect of climate change on the structure of populations of spring wheat pathogens in the Volga region. Agrar. Rep. South-East 2010, 1, 31–36. (In Russian) [Google Scholar]
- Volkova, G.V.; Kudinova, O.A.; Matveeva, I.P. Virulence and diversity of Puccinia striiformis in South Russia. Phytopathol. Mediterr. 2021, 60, 119–127. [Google Scholar] [CrossRef]
- Hovmøller, M.S.; Rodriguez-Algaba, J.; Thach, T.; Sørensen, C.K. Race Typing of Puccinia striiformis on Wheat. In Methods in Molecular Biology; Periyannan, S., Ed.; Springer Science+Business Media LLC: Berlin Germany, 2017; Volume 1659, pp. 29–40. [Google Scholar] [CrossRef]
- Gultyaeva, E.; Shaydayuk, E.; Gannibal, P.; Kosman, E. Analysis of host-specific differentiation of Puccinia striiformis in the South and North-West of the European Part of Russia. Plants 2021, 10, 2497. [Google Scholar] [CrossRef] [PubMed]
- Gassner, G.; Straib, W. Untersuchungen Über die Infektions bedingungen von Puccinia glumarum und Puccinia graminis. Arb. Biol. Reichsanst. Land-Forst- wirtsch Berlin-Dahlem 1932, 16, 609–629. [Google Scholar]
- McIntosh, R.A.; Wellings, C.R.; Park, R.F. Wheat Rusts: An Atlas of Resistance Genes; CSIRO: Canberra, Australia, 1995. [Google Scholar]
- Kosman, E. Measure of multilocus correlation as a new parameter for study of plant pathogen populations. Phytopathology 2003, 93, 1464–1470. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kosman, E. Difference and diversity of plant pathogen populations: A new approach for measuring. Phytopathology 1996, 86, 1152–1155. [Google Scholar]
- Kosman, E.; Leonard, K.J. Conceptual analysis of methods applied to assessment of diversity within and distance between populations with asexual or mixed mode of reproduction. New Phytol. 2007, 174, 683–696. [Google Scholar] [CrossRef] [PubMed]
- Kosman, E. Measuring diversity: From individuals to populations. Eur. J. Plant Pathol. 2014, 138, 467–486. [Google Scholar] [CrossRef]
- Czajowski, G.; Kosman, E.; Słowacki, P.; Park, R.F.; Czembor, P. Assessing new SSR markers for utility and informativeness in genetic studies of brown rust fungi on wheat, triticale and rye. Plant Pathol. 2021, 70, 1110–1122. [Google Scholar] [CrossRef]
- Kosman, E.; Ben-Yehuda, P.; Manisterski, J. Diversity of virulence phenotypes among annual populations of wheat leaf rust in Israel from 1993 to 2008. Plant Pathol. 2014, 63, 563–571. [Google Scholar] [CrossRef]
- Scheiner, S.M.; Kosman, E.; Presley, S.J.; Willig, M.R. Decomposing functional diversity. Methods Ecol. Evol. 2017, 97, 809–820. [Google Scholar] [CrossRef] [Green Version]
- Kosman, E.; Chen, X.; Dreiseitl, A.; McCallum, B.; Lebeda, A.; Ben-Yehuda, P.; Gultyaeva, E.; Manisterski, J. Functional variation of plant-pathogen interactions: New concept and methods for virulence data analyses. Phytopathology 2019, 109, 1324–1330. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.; Kosman, E.; Sharon, O.; Ezrati, S.; Sharon, A. Significant host- and environment-dependent differentiation among highly sporadic fungal endophyte communities in cereal crops-related wild grasses. Environ. Microbiol. 2020, 22, 3357–3374. [Google Scholar] [CrossRef]
- Kosman, E.; Burgio, K.R.; Presley, S.J.; Willig, M.R.; Scheiner, S.M. Conservation prioritization based on trait-based metrics illustrated with global parrot distributions. Divers. Distrib. 2019, 25, 1156–1165. [Google Scholar] [CrossRef] [Green Version]
- Kosman, E.; Dinoor, A.; Herrmann, A.; Schachtel, G.A. Virulence Analysis Tool (VAT). User Manual. 2008. Available online: http://www.tau.ac.il/lifesci/departments/plant_s/members/kosman/VAT.html (accessed on 25 August 2022).
- Schachtel, G.A.; Dinoor, A.; Herrmann, A.; Kosman, E. Comprehensive evaluation of virulence and resistance data: A new analysis tool. Plant Dis. 2012, 96, 1060–1063. [Google Scholar] [CrossRef] [Green Version]
- Morgounov, A.; Ablova, I.; Babayants, O.; Babayants, L.; Bespalova, L.; Khudokormova, Z.; Litvinenko, N.; Shamanin, V.; Syukov, V. Genetic protection of wheat from rusts and development of resistant varieties in Russia and Ukraine. Euphytica 2011, 179, 297–311. [Google Scholar] [CrossRef]
- Kosman, E.; Ben-Yehuda, P.; Manisterski, J.; Anikster, Y.; Sela, H. Virulence survey of Puccinia striiformis in Israel revealed considerable changes in the pathogen population during the period 2001 to 2019. Plant Dis. 2022, 106, 2823–2830. [Google Scholar] [CrossRef] [PubMed]
- Wan, A.M.; Chen, D.X.M.; He, Z.H. Wheat stripe rust in China. Aust. J. Agric. Res. 2007, 58, 605–619. [Google Scholar] [CrossRef]
- Lagudah, E.S. Molecular genetics of race non-specific rust resistance in wheat. Euphytica 2011, 179, 81–91. [Google Scholar] [CrossRef]
- Mago, R.; Tabe, L.; McIntosh, R.A.; Pretorius, Z.; Kota, R.; Paux, E.; Wicker, T.; Breen, J.; Lagudah, E.S.; Ellis, J.G.; et al. A multiple resistance locus on chromosome arm 3BS in wheat confers resistance to stem rust (Sr2), leaf rust (Lr27) and powdery mildew. Theor. Appl. Genet. 2011, 123, 615–623. [Google Scholar] [CrossRef]
- Herrera-Foessel, S.A.; Singh, R.P.; Lillemo, M.; Huerta-Espino, J.; Bhavani, S.; Singh, S.; Lan, C.; Calvo-Salazar, V.; Lagudah, E.S. Lr67/Yr46 confers adult plant resistance to stem rust and powdery mildew in wheat. Theor. Appl. Genet. 2014, 127, 781–789. [Google Scholar] [CrossRef]
- Guo, Q.; Zhang, Z.J.; Xu, Y.B.; Li, G.H.; Feng, J.; Zhou, Y. Quantitative trait loci for high-temperature adult-plant and slow-rusting resistance to Puccinia striiformis f. sp. tritici in wheat cultivars. Phytopathology 2008, 98, 803–809. [Google Scholar] [CrossRef] [Green Version]
- State Register for Selection Achievements Admitted for Usage (National List). Vol. 1.“Plant Varieties” (Official Publication); FGBNU «Rosinformagrotech»: Moscow, Russia, 2022; 646p, Available online: https://reestr.gossortrf.ru/ (accessed on 25 August 2022). (In Russian)
- Gultyaeva, E.; Shaydayuk, E.; Gannibal, P. Leaf rust resistance genes in wheat cultivars registered in Russia and their influence on adaptation processes in pathogen populations. Agriculture 2021, 11, 319. [Google Scholar] [CrossRef]
- Coriton, O.; Jahier, J.; Leconte, M.; Huteau, V.; Trotoux, G.; Dedryver, F.; de-Vallavieille-Pope, C.; Miedaner, T. Double dose efficiency of the yellow rust resistance gene Yr17 in bread wheat lines. Plant Breed. 2019, 139, 263–271. [Google Scholar] [CrossRef]
- Bayles, R.A.; Flath, K.; Hovmøller, M.S.; de Vallavieille-Pope, C. Breakdown of the Yr17 resistance to yellow rust of wheat in northern Europe—A case study by the yellow rust sub-group of COST 817. Agronomie 2000, 20, 805–811. [Google Scholar] [CrossRef] [Green Version]
- Shahin, A.A. Occurrence of new races and virulence changes of the wheat stripe rust pathogen (Puccinia striiformis f. sp. tritici) in Egypt. Arch. Phytopathol. Plant Prot. 2020, 53, 552–569. [Google Scholar] [CrossRef]
- Amil, R.E.; Ali, S.; Bahri, B.; Leconte, M.; de Vallavieille-Pope, C.; Nazari, K. Pathotype diversification in the invasive PstS2 clonal lineage of Puccinia striiformis f. sp. tritici causing yellow rust on durum and bread wheat in Lebanon and Syria in 2010–2011. Plant Pathol. 2020, 69, 618–630. [Google Scholar] [CrossRef]
- Gangwar, P.; Kumar, S.; Bhardwaj, S.C.; Prasad, P.; Kashyap, P.L.; Khan, H.; Singh, G.P.; Savadi, S. Virulence and molecular diversity among Puccinia striiformis f. sp. tritici pathotypes identified in India between 2015 and 2019. Crop Prot. 2021, 148, 105717. [Google Scholar] [CrossRef]
- Chuprina, V.P.; Sokolov, M.S.; Anpilogova, L.K.; Kobileva, E.A.; Levashova, G.I.; Pikushova, V.V. Protection of wheat crops against stripe rust, taking into account the epiphytotic and biotrophic characteristics of its pathogen Puccinia striiformis West. Agrochemistry 1999, 7, 81–94. (In Russian) [Google Scholar]
- Ziyaev, Z.M.; Sharma, R.C.; Nazari, K.; Morgounov, A.I.; Amanov, A.A.; Ziyadullaev, Z.F.; Khalikulov, Z.I.; Alikulov, S.M. Improving wheat stripe rust resistance in Central Asia and the Caucasus. Euphytica 2011, 179, 197–207. [Google Scholar] [CrossRef]
- Kokhmetova, A.; Rsaliyev, A.; Malysheva, A.; Atishova, M.; Kumarbayeva, M.; Keishilov, Z. Identification of stripe rust resistance genes in common wheat cultivars and breeding lines from Kazakhstan. Plants 2021, 10, 2303. [Google Scholar] [CrossRef]
- Feodorova-Fedotova, L.; Bankina, B. Occurrence of genetic lineages of Puccinia striiformis in Latvia. Res. Rural. Dev. 2020, 35, 27–32. [Google Scholar] [CrossRef]
- Gultyaeva, E.I.; Bespalova, L.A.; Ablova, I.B.; Shaydayuk, E.L.; Khudokormova, Z.N.; Yakovleva, D.R.; Titova, Y.A. Wild grasses as the reservoirs of infection of rust species for winter soft wheat in the Northern Caucasus. Vavilov J. Genet. Breed. 2021, 25, 638–646. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Region | Location | Year | Samples 1 | Isolates 2 |
|---|---|---|---|---|
| North Caucasus (NC) | Dagestan (D) | 2020 | 5 | 8 |
| 2021 | 19 | 18 | ||
| Total Da | 24 | 26 | ||
| Krasnodar (Kr) | 2019 | 4 | 4 | |
| 2020 | 5 | 8 | ||
| 2021 | 5 | 7 | ||
| Total Kr | 14 | 19 | ||
| Rostov (Ro) | 2021 | 6 | 6 | |
| Total NC | 44 | 51 | ||
| North-West (NW) | Saint Petersburg (SPb) | 2019 | 8 | 8 |
| 2020 | 15 | 23 | ||
| 2021 | 11 | 11 | ||
| Total NW | 34 | 42 | ||
| Low Volga (LV) | Saratov (Sa) | 2020 | 2 | 2 |
| 2021 | 2 | 2 | ||
| Total LV | 4 | 4 | ||
| Central Black Earth region (CBE) | Tambov (Ta) | 2020 | 2 | 2 |
| Volga-Vyatka (VV) | Kirov (Ki) | 2020 | 2 | 4 |
| West Siberia (WS) | Novosibirsk (No) | 2019 | 4 | 4 |
| 2020 | 2 | 4 | ||
| 2021 | 2 | 4 | ||
| Total No | 8 | 12 | ||
| Krasnoyarsk (Kk) | 2021 | 1 | 4 | |
| Total WS | 9 | 16 | ||
| Total Russia | 95 | 117 |
| Yr Genes | Line with the Corresponding Yr Genes | NC 1 | NW | LV | CBE | VV | WS | Overall | |||
|---|---|---|---|---|---|---|---|---|---|---|---|
| D 2 | Kr | Ro | SPb | Sa | Ta | Ki | No | Kk | |||
| Yr1 | Yr1/6*Avocet S, Chinese 166 | 42 | 95 | 100 | 55 | 50 | 100 | 100 | 100 | 100 | 68 |
| Yr2, Yr25, Yr+3 | Heines VII | 96 | 84 | 100 | 100 | 50 | 100 | 100 | 92 | 100 | 94 |
| Yr3, Yr+ | Vilmorin 23 | 69 | 89 | 100 | 79 | 0 | 100 | 100 | 75 | 100 | 78 |
| Yr4, Yr+ | Hybrid 46 | 88 | 95 | 0 | 62 | 50 | 0 | 100 | 83 | 100 | 73 |
| Yr5 | Yr5/6*Avocet S | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Yr6 | Yr6/6*Avocet S | 92 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 100 | 98 |
| Yr7 | Yr7/6*Avocet S | 100 | 95 | 33 | 95 | 50 | 50 | 100 | 50 | 100 | 86 |
| Yr8 | Yr8/6*Avocet S | 100 | 95 | 100 | 98 | 100 | 100 | 0 | 50 | 0 | 88 |
| Yr9 | Yr9/6*Avocet S | 85 | 100 | 100 | 100 | 50 | 100 | 100 | 100 | 100 | 95 |
| Yr10 | Yr10/6*Avocet S | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Yr15 | Yr15/6*Avocet S | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Yr17 | Yr17/6*Avocet S | 0 | 0 | 0 | 12 | 25 | 0 | 0 | 0 | 0 | 5 |
| Yr24 | Yr24/6*Avocet S | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| YrSD, Yr25, Yr+ | Strubes Dickkopf (SD) | 46 | 26 | 50 | 74 | 100 | 0 | 100 | 100 | 100 | 62 |
| Yr27 | Yr27/6*Avocet S | 100 | 63 | 0 | 67 | 100 | 100 | 100 | 83 | 100 | 75 |
| Yr32, Yr25, Yr+ | Carstens V | 58 | 74 | 0 | 67 | 75 | 50 | 100 | 83 | 100 | 66 |
| YrSu, Yr+ | Suwon 92/Omar (Su) | 92 | 100 | 67 | 95 | 100 | 100 | 100 | 100 | 100 | 95 |
| YrSp | YrSP/6*Avocet S | 50 | 58 | 100 | 29 | 0 | 0 | 100 | 67 | 100 | 48 |
| YrND, Yr3 | Nord Desprez (ND) | 15 | 0 | 0 | 17 | 50 | 50 | 100 | 33 | 100 | 21 |
| Yr2,Yr6,Yr25, Yr+ | Heines Peko (HP) | 92 | 100 | 100 | 88 | 50 | 50 | 100 | 100 | 100 | 91 |
| Number of isolates | 26 | 19 | 6 | 42 | 4 | 2 | 2 | 12 | 4 | 117 | |
| Region | Location | Year | Isolates | Pathotypes | Pathotype Composition and Frequency | |
|---|---|---|---|---|---|---|
| NC | D | 2020 | 8 | 6 | P1 1 (38%), P5 (12%), P18 (12%), P19 (12%), P20 (12%), P21 (12%), | |
| 2021 | 18 | 14 | P1 (17%), P2 (5%), P3 (5%), P5 (5%), P6 (11%), P7 (11%), P22 (5%), P23 (5%), P24 (5%), P25 (5%), P26 (5%), P27 (5%), P28 (5%), P29 (5%) | |||
| Kr | 2019 | 3 | 3 | P1 (33%), P30 (33%), P31 (33%) | ||
| 2020 | 8 | 6 | P2 (25%), P8 (25%), P32 (12%), P33 (12%), P34 (12%), P35 (12%) | |||
| 2021 | 7 | 6 | P2 (29%), P1 (14%), P8 (14%), P36 (14%), P37 (14%), P38 (14%) | |||
| R | 2021 | 6 | 6 | P39 (17%), P40 (17%), P41 (17%), P42 (17%), P43 (17%), P44 (17%) | ||
| NW | SPb | 2019 | 8 | 7 | P9 (25%), P1 (12%), P45 (12%), P46 (12%), P47 (12%), P48 (12%), P49 (12%), P50 (12%) | |
| 2020 | 23 | 22 | P9 (9%), P1 (4%), P2 (4%), P3 (4%), P10 (8%), P51 (4%), P52 (4%), P53 (4%), P54 (4%), P55 (4%), P56 (4%), P57 (4%), P58 (4%), P59 (4%), P60 (4%), P61 (4%), P62 (4%), P63 (4%), P66 (4%), P65 (4%), P66 (4%), P67 (4%), P68 (4%) | |||
| 2021 | 11 | 8 | P9 (18%), P11 (18%), P12 (18%), P69 (9%), P70 (9%), P71 (9%), P72 (9%), P73 (9%) | |||
| LW | Sar | 2020 | 2 | 2 | P74 (50%), P75 (50%) | |
| 2021 | 2 | 1 | P13 (100%) | |||
| CBE | Tam | 2020 | 2 | 2 | P76 (50%), P77 (50%) | |
| VV | Kir | 2020 | 2 | 1 | P4 (100%) | |
| WS | Nov | 2019 | 3 | 2 | P14 (67%), P15 (33%) | |
| 2020 | 4 | 2 | P15 (50%), P16 (50%) | |||
| 2021 | 4 | 3 | P17 (50%) P78 (25%), P79 (25%) | |||
| Kr-k | 2020 | 4 | 1 | P4 (100%) | ||
| NC | NW | C | WS | Overall, for Russia | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Parameters | D | Kr | Ro | Total | SPb | Sa | Ta | Ki | Total | No | Kk | Total | |
| Number of isolates | 26 | 19 | 6 | 51 | 42 | 4 | 2 | 2 | 8 | 12 | 4 | 16 | 117 |
| Number of pathotypes | 18 | 11 | 6 | 34 | 36 | 3 | 2 | 1 | 6 | 7 | 1 | 8 | 79 |
| Predominant abundances 1, % | 23.1 | 21.1 | 16.7 | 17.6 | 7.1 | 50 | 50 | 100 | 25 | 25 | 100 | 25 | 9.4 |
| Average virulence complexity of pathotypes | 10.7 | 11.3 | 9.5 | 10.6 | 11.1 | 9.3 | 10 | 14 | 10.3 | 11.8 | 14 | 12.4 | 10.9 |
| Prevailing Pst pathotypes (P#: avirulence/virulence formula of Yr genes) and their abundances (%): | |||||||||||||
| P1. Yr: 5,10,15,17, 24,ND/1,2,3, 4,6,7,8,9,25,27,32,Su,SP,HP | 23 | 16 | 0 | 18 | 5 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 9 |
| P2. Yr: 5,10,15,17,24,25,32,Sp, ND/1,2, 3,4,6,7,8,9,27,Su,HP | 4 | 21 | 0 | 10 | 2 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 5 |
| P3. Yr: 1,5,10,15,17,24,32, SP, ND/2,3,4,6,7,8,9,25,27,Su,HP | 4 | 0 | 0 | 3 | 2 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 2 |
| P4. Yr: 5,8,10,15,17,24/1,2,3,4, 6,7,9,25,27,32,Su,Sp, ND,HP | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 100 | 0 | 4 |
| P5. Yr: 5,9,10,15,17,24,25,ND/1,2,3,4,6,7,8,27,32,Su,SP,HP | 4 | 0 | 0 | 4 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 2 |
| P6. Yr: 1,3,5,10,15,17,24,25, 32,Sp,ND/2,4,6,7,8,9,27,Su, HP | 8 | 0 | 0 | 4 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 2 |
| P7. Yr: 1,3,5,10,15,17,24,32,SP, ND/2,4,6,7,8,9,25,27, Su,HP | 8 | 0 | 0 | 4 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 2 |
| P8. Yr: 5,10,15,17,24,25,SP,ND/ 1,2,3,4,6,7,8,9,27, 32,Su, HP | 0 | 16 | 0 | 6 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 3 |
| P9. Yr: 5,10,15,17,24,SP,ND/1,2,3, 4,6,7,8,9,25,27, 32,Su,HP | 0 | 0 | 0 | 0 | 7 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 3 |
| P10. Yr: 1,4,5,10,15,17,24,SP,ND/2,3,6,7,8,9,25,27,32,Su,HP | 0 | 0 | 0 | 0 | 5 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 2 |
| P11. Yr: 1,5,10,15,17,24,SP,ND/2,3, 4,6,7,8,9,25,27, 32,Su,HP | 0 | 0 | 0 | 0 | 5 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 2 |
| P12. Yr: 4,5,10,15,17,24,SP/1,2,3, 6,7,8,9,25,27,32,Su,ND,HP | 0 | 0 | 0 | 0 | 5 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 2 |
| P13. Yr: 2,3,5,7,10,15,17,24,SP,ND/1,4,6,8,9,25,27,32,Su,HP | 0 | 0 | 0 | 0 | 0 | 50 | 0 | 0 | 0 | 0 | 0 | 0 | 2 |
| P14. Yr: 3,5,8,10,15,17,24,ND/1,2,4,6,7,9,25,27, 32,Su, SP,HP | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 25 | 0 | 0 | 3 |
| P15. Yr: 5,8,10,15,17,24,ND1,2,3, 4,6,7,9,25,27,32,Su,SP,HP | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 17 | 0 | 0 | 2 |
| P16. Yr: 5,7,10,15,17,24/1,2,3,4,6, 8,9,25,27,32,Su,SP,ND,HP | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 17 | 0 | 0 | 2 |
| P17. Yr: 5,7,10,15,17,24,27,SP, ND/1,2,3,4,6,8,9,25,32,Su,HP | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 17 | 0 | 0 | 2 |
| Pst Collection | Pathotypes | KW | ||
|---|---|---|---|---|
| Krasnodar | 11 | 0.21 | 2.99 | 0.12 |
| Rostov | 6 | 0.12 | 1.56 | 0.11 |
| Dagestan | 18 | 0.28 | 5.49 | 0.26 |
| North-West | 36 | 0.31 | 11.37 | 0.3 |
| West Siberia | 8 | 0.27 | 2.77 | 0.25 |
| Central European regions | 6 | 0.36 | 2.41 | 0.35 |
| Regional Population | Abbreviation | Number of Virulence Pathotypes in Five Subgroups | Total | ||||
|---|---|---|---|---|---|---|---|
| 1 | 2 | 3 | 4 | 5 | |||
| Dagestan | D | 11 | 4 | 3 | 18 | ||
| North-West | NW | 4 | 11 | 12 | 7 | 2 | 36 |
| West Siberia | WS | 6 | 2 | 8 | |||
| Central | C | 2 | 3 | 6 | |||
| Kr | Ro | D | NW | WS | C | |
|---|---|---|---|---|---|---|
| Kr a | - | 0.99 | 0.99 | 0.98 | 0.97 | 0.99 |
| Ro b | 0.22 | - | 0.99 | 0.99 | 0.99 | 0.99 |
| D | 0.17 | 0.29 | - | 0.94 | 0.99 | 0.97 |
| NW | 0.16 | 0.24 | 0.14 | - | 0.38 | 0.91 |
| WS | 0.18 | 0.25 | 0.23 | 0.2 | - | 0.89 |
| C b | 0.24 | 0.32 | 0.19 | 0.2 | 0.22 | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gultyaeva, E.; Shaydayuk, E.; Kosman, E. Virulence Diversity of Puccinia striiformis f. sp. Tritici in Common Wheat in Russian Regions in 2019–2021. Agriculture 2022, 12, 1957. https://doi.org/10.3390/agriculture12111957
Gultyaeva E, Shaydayuk E, Kosman E. Virulence Diversity of Puccinia striiformis f. sp. Tritici in Common Wheat in Russian Regions in 2019–2021. Agriculture. 2022; 12(11):1957. https://doi.org/10.3390/agriculture12111957
Chicago/Turabian StyleGultyaeva, Elena, Ekaterina Shaydayuk, and Evsey Kosman. 2022. "Virulence Diversity of Puccinia striiformis f. sp. Tritici in Common Wheat in Russian Regions in 2019–2021" Agriculture 12, no. 11: 1957. https://doi.org/10.3390/agriculture12111957
APA StyleGultyaeva, E., Shaydayuk, E., & Kosman, E. (2022). Virulence Diversity of Puccinia striiformis f. sp. Tritici in Common Wheat in Russian Regions in 2019–2021. Agriculture, 12(11), 1957. https://doi.org/10.3390/agriculture12111957