
Can Mathematical Models Describe Spear Rot Progress in Oil Palm Trees? A Five-Year Black Weevil-Disease Assessment from Ecuador

 and
and
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Geographic Location and Agroclimatic Characteristics of the Study Area
2.2. Field Experiment Conditions
2.3. Quantification of Black Weevil
2.4. Capture and Morphological Characterization of Rhynchophorus palmarum
Sampling
2.5. Characterization of Adult Insects
2.6. Spear Rot Assessment
2.7. Morphological and Molecular Characterization of Microorganism
Sampling
2.8. Isolation of Microorganisms
2.9. Morphological Characterization
2.10. Molecular Characterization
2.11. Fruit Bunches Yield
2.12. Statistical Analyses
3. Results
3.1. Brief Description of The Adult Species
3.2. Characterization of Plant Pathogen
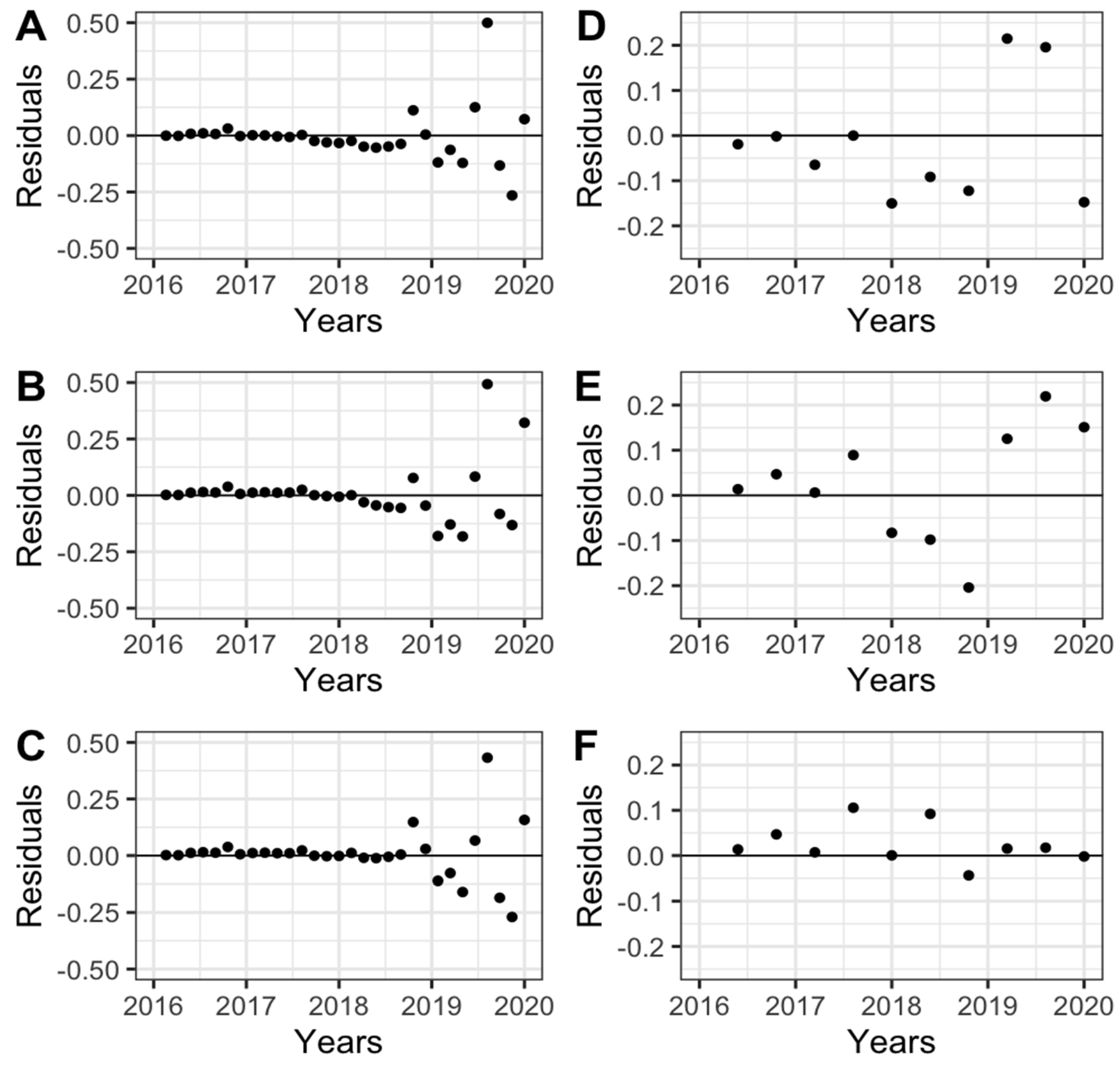
3.3. Temporal Analysis of Black Weevil and Spear Rot
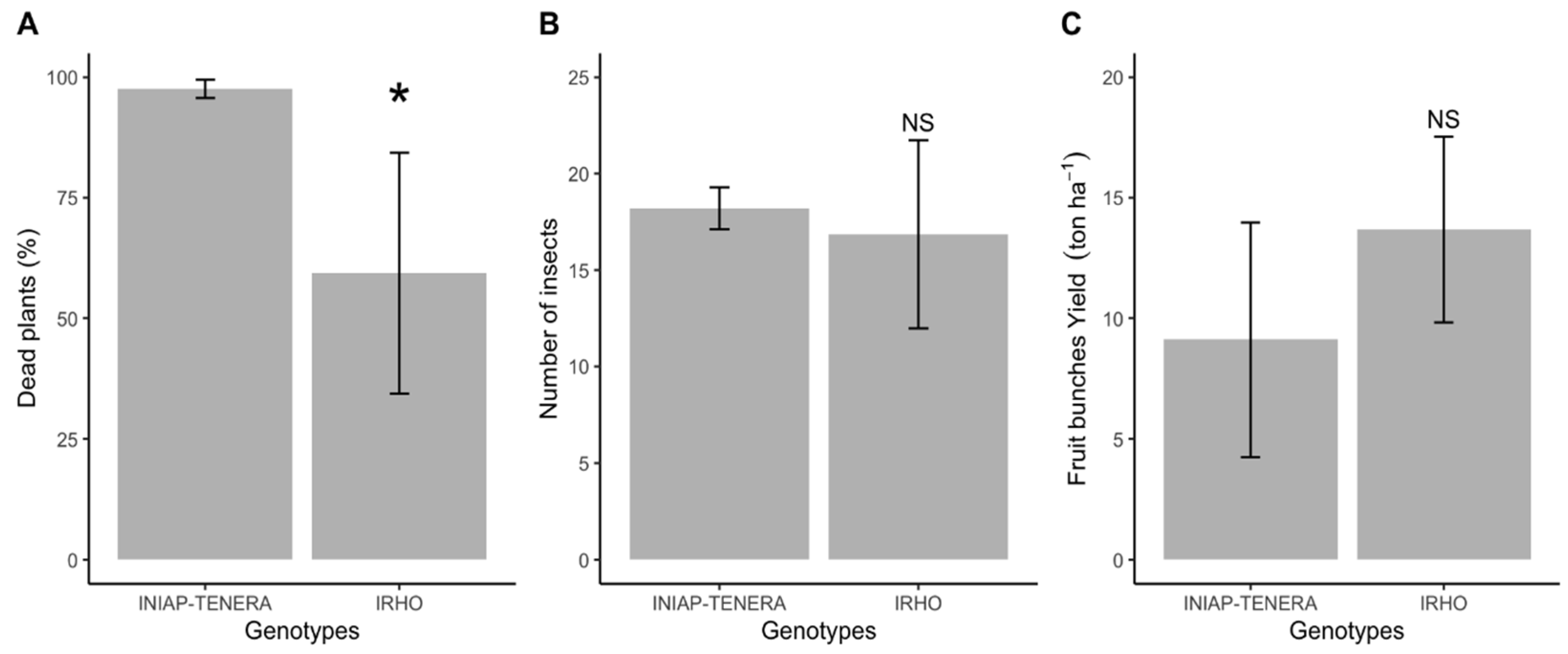
3.4. Dead Plants by Spear Rot, Number of Black Weevil Adults, and Fruit Bunches Yield
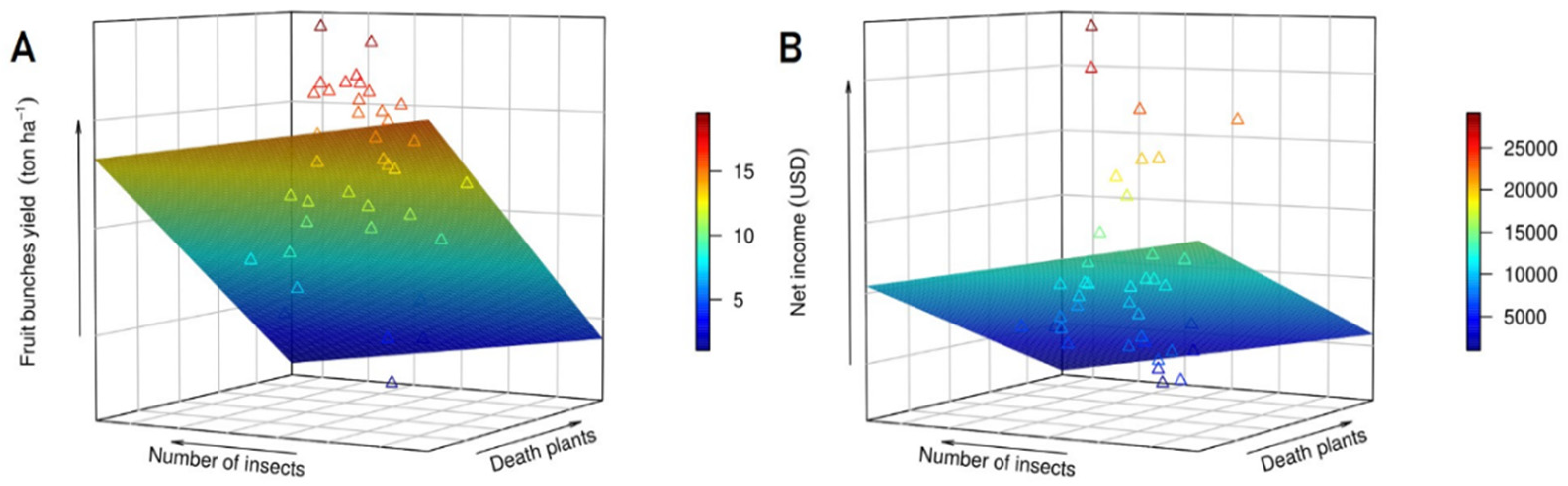
3.5. Relationship and Correlation among Variables
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Acknowledgments
Conflicts of Interest
References
- Nair, P.K. Tree Crops. Harvesting Cash from the World’s Important Cash Crops, 1st ed.; Springer Nature: Cham, Switzerland, 2021; pp. 249–285. [Google Scholar] [CrossRef]
- Instituto Nacional de Estadística y Censos (INEC). Boletín Técnico. Encuesta de Superficie y Producción Agropecuaria Continua, 1st ed.; INEC: Quito, Ecuador, 2020; pp. 1–13. Available online: https://www.ecuadorencifras.gob.ec/documentos/web-inec/Estadisticas_agropecuarias/espac/espac-2019/Boletin%20Tecnico%20ESPAC_2019.pdf (accessed on 14 June 2021).
- Aucique-Pérez, C.E.; Daza, E.S.; Ávila-Diazgranados, R.A.; Romero, H.M. Chlorophyll a fluorescence and leaf temperature are early indicators of oil palm diseases. Sci. Agric. 2020, 77, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Landero-Torres, I.; Presa-Parra, E.; Galindo-Tovar, M.E.; Leyva-Ovalle, O.R.; Murguía-González, J.; Valenzuela-González, J.E.; García-Martínez, M.Á. Variación temporal y espacial de la abundancia del Picudo negro (Rynchophorus palmarum L., Coleoptera: Curculionidae) en cultivos de palmas ornamentales del centro de Veracruz, México. Southwest. Entomol. 2015, 40, 179–188. [Google Scholar] [CrossRef]
- Plata-Rueda, A.; Martínez, L.C.; Fernandes, F.L.; De Sousa Ramalho, F.; Zanuncio, J.C.; Serrão, J.E. Interactions between the bud rot disease of oil palm and Rhynchophorus palmarum (Coleoptera: Curculionidae). J. Econ. Entomol. 2016, 109, 962–965. [Google Scholar] [CrossRef] [PubMed]
- Murguía-González, J.; Landero-Torres, I.; Leyva-Ovalle, O.R.; Galindo-Tovar, M.E.; Llarena-Hernández, R.C.; Presa-Parra, E.; García-Martínez, M.A. Efficacy and cost of trap–bait combinations for capturing Rhynchophorus palmarum L. (Coleoptera: Curculionidae) in ornamental palm polycultures. Neotrop. Entomol. 2018, 47, 302–310. [Google Scholar] [CrossRef]
- Hoddle, M.S.; Hoddle, C.D.; Milosavljević, I. Quantification of the life time flight capabilities of the south American palm weevil, Rhynchophorus palmarum (L.) (Coleoptera: Curculionidae). Insects 2021, 12, 126. [Google Scholar] [CrossRef] [PubMed]
- Pinho, R.; Farias, P.R.; Rodrigues, K.C.; Tinôco, R.; Santos, A.V.; Marssena, R.T. Distribuição espacial de Rhynchophorus palmarum em palma de óleo no Estado do Pará, Amazônia. Cienc. Agrar. 2016, 59, 22–31. [Google Scholar] [CrossRef]
- Milosavljević, I.; El-Shafie, H.A.F.; Faleiro, J.R.; Hoddle, C.D.; Lewys, M.; Hoddle, M.S. Palmageddon: The wasting of ornamental palms by invasive palm weevils, Rhynchophorus spp. J. Pest. Sci. 2019, 92, 143–156. [Google Scholar] [CrossRef]
- Saïd, I.; Kaabi, B.; Rochat, D. Evaluation and modeling of synergy to pheromone and plant kairomone in American palm weevil. Chem. Cent. J. 2011, 5, 14. [Google Scholar] [CrossRef] [Green Version]
- Suwandi–Akino, S.; Kondo, N. Common spear rot of oil palm in Indonesia. Plant Dis. 2012, 96, 537–543. [Google Scholar] [CrossRef] [Green Version]
- de Assis Costa, O.Y.; Tupinambá, D.D.; Bergmann, J.C.; Barreto, C.C.; Quirino, B.F. Fungal diversity in oil palm leaves showing symptoms of fatal yellowing disease. PLoS ONE 2018, 13, e0191884. [Google Scholar] [CrossRef] [Green Version]
- Ronquillo-Narváez, M.; de Jensen, C.E.; Bernal, G. Fusarium spp. asociados a la pudrición del cogollo de la palma aceitera (Elaeis guineensis Jacq.) en Ecuador. J. Agrie. Univ. P. R. 2013, 97, 135–148. [Google Scholar]
- De Franqueville, H. Oil palm bud rot in Latin America. Exp. Agric. 2003, 39, 225–240. [Google Scholar] [CrossRef]
- Torres, G.A.; Sarria, G.A.; Varon, F.; Coffey, M.D.; Elliott, M.L.; Martinez, G. First report of Bud rot caused by Phytophthora palmivora on African oil palm in Colombia. Plant Dis. 2010, 94, 1163. [Google Scholar] [CrossRef] [PubMed]
- Van De Lande, H.L.; Zadoks, J.C. Spatial patterns of spear rot in oil palm plantations in Surinam. Plant Pathol. 1999, 48, 189–201. [Google Scholar] [CrossRef]
- Perthuis, B. Association of the soil insect Scaptocoris minor Berg (Heteroptera, Cydnidae) with foci of bud-rot diseases of oil palm in eastern Ecuador. Int. J. Trop. Insect. Sci. 2020, 41, 1883–1887. [Google Scholar] [CrossRef]
- Rivas-Figueroa, F.; Herrera-Isla, L.; Borrás-Hidalgo, O. Molecular diagnosis of Fusarium spp. isolates associated to bud rot of Oil palm in Ecuador. Biotecnol. Apl. 2015, 32, 2221–2223. [Google Scholar]
- Swinburne, T. Fatal yellows, bud rot and spear rot of African oil palm—A comparison of the symptoms of these diseases in Brazil, Ecuador and Colombia. Planter 1993, 69, 15–23. [Google Scholar]
- Avila-Diazgranados, R.A.; Daza, E.S.; Navia, E.; Romero, H.M. Response of various oil palm materials (Elaeis guineensis and Elaeis oleifera × Elaeis guineensis interspecific hybrids) to bud rot disease in the southwestern oil palm-growing area of Colombia. Agron. Colomb. 2016, 34, 74–81. [Google Scholar] [CrossRef]
- Gitau, C.W.; Gurr, G.M.; Dewhurst, C.F.; Fletcher, M.J.; Mitchell, A. Insect pests and insect-vectored diseases of palms. Aust. J. Entomol. 2009, 48, 328–342. [Google Scholar] [CrossRef]
- Bergamin Filho, A.; Amorim, L.; Laranjeira, F.F.; Berger, R.D.; Hau, B. Análise temporal do amarelecimento fatal do dendezeiro como ferramenta para elucidar sua etiologia. Fitopatol. Bras. 1998, 23, 391–396. [Google Scholar]
- Savary, S.; Nelson, A.D.; Djurle, A.; Esker, P.D.; Sparks, A.; Amorim, L.; Bergamin Filho, A.; Caffi, T.; Castilla, N.; Garrett, K.; et al. Concepts, approaches, and avenues for modelling crop health and crop losses. Eur. J. Agron. 2018, 100, 4–18. [Google Scholar] [CrossRef]
- Van De Lande, H.L. Spatio-temporal analysis of spear rot and ‘marchitez sorpresiva’ in African oil palm in Surinam. Neth. J. Plant Pathol. 1993, 99, 129–138. [Google Scholar] [CrossRef]
- Benítez-Sastoque, E.R. Epidemiología de la Pudrición del Cogollo de la Palma de Aceite. Ph.D. Thesis, Universidad Nacional de Colombia, Bogotá, Colombia, 2011. Available online: https://repositorio.unal.edu.co/handle/unal/52477 (accessed on 14 June 2021).
- Köppen, W. Classification of climates according to temperature, precipitation and seasonal cycle. Petermanns. Geogr. Mitt. 1918, 64, 193–203. [Google Scholar]
- Chamorro, M.L. An illustrated synoptic key and comparative morphology of the larvae of Dryophthorinae (Coleoptera, Curculionidae) genera with emphasis on the mouthparts. Diversity 2019, 11, 4. [Google Scholar] [CrossRef] [Green Version]
- Vásquez-Ordóñez, A.A.; Löhr, B.L.; Marvaldi, A.E. Comparative morphology of the larvae of the palm weevils Dynamis borassi (Fabricius) and Rhynchophorus palmarum (Linnaeus) (Curculionidae: Dryophthorinae): Two major pests of peach palms in the Neotropics. Pap. Avulsos. Zool. 2020, 60, e202060.27. [Google Scholar] [CrossRef]
- Martínez-López, G.; Torres, G.A. Presencia de la pudrición del cogollo de palma de aceite (PC) en plantas de vivero. Palmas 2007, 28, 13–20. [Google Scholar]
- Gupta, V.K.; Tuohy, M.G.; Gaur, R. Methods for high-quality DNA extraction from fungi. In Laboratory Protocols in Fungal Biology, 1st ed.; Gupta, V., Tuohy, M., Ayyachamy, M., Turner, K., O’Donovan, A., Eds.; Springer: New York, NY, USA, 2013; pp. 403–406. [Google Scholar] [CrossRef]
- Liu, Y.J.; Whelen, S.; Hall, B.D. Phylogenetic relationships among ascomycetes: Evidence from an RNA polymerse II subunit. Mol. Biol. Evol. 1999, 16, 1799–1808. [Google Scholar] [CrossRef]
- Geiser, D.M.; del Mar Jiménez-Gasco, M.; Kang, S.; Makalowska, I.; Veeraraghavan, N.; Ward, T.J.; Zhang, N.; Kuldau, G.A.; O’donnell, K. FUSARIUM-ID v. 1.0: A DNA sequence database for identifying Fusarium. Eur. J. Plant. Pathol. 2004, 110, 473–479. [Google Scholar] [CrossRef]
- Stecher, G.; Tamura, K.; Kumar, S. Molecular evolutionary genetics analysis (MEGA) for macOS. Mol. Biol. Evol. 2020, 37, 1237–1239. [Google Scholar] [CrossRef] [PubMed]
- Berger, R.D. Comparison of the Gompertz and Logistic equations to describe plant disease progress. Phytopathology 1981, 71, 716–719. [Google Scholar] [CrossRef]
- Zwietering, M.H.; Jongenburger, I.; Rombouts, F.M.; Van’t Riet, K. Modeling of the bacterial growth curve. Appl. Environ. Microbiol. 1990, 56, 1875–1881. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tjørve, K.M.; Tjørve, E. The use of Gompertz models in growth analyses, and new Gompertz-model approach: An addition to the Unified-Richards family. PLoS ONE 2017, 12, e0178691. [Google Scholar] [CrossRef] [PubMed]
- Turner, H.; Firth, D. Generalized Nonlinear Models in R: An Overview of the Gnm Package Generalized Linear Models Preamble, 1st ed.; ESRC National Centre for Research Methods: Coventry, UK, 2020; pp. 1–52. Available online: https://eprints.ncrm.ac.uk/id/eprint/472/1/0607_overview_of_the_gnm_package.pdf (accessed on 14 June 2021).
- Archontoulis, S.V.; Miguez, F.E. Nonlinear Regression models and applications in agricultural research. J. Agron. 2015, 107, 786–798. [Google Scholar] [CrossRef] [Green Version]
- R Studio Team. RStudio. Available online: http://www.rstudio.com/ (accessed on 14 June 2021).
- Wattanapongsiri, A. A Revision to the Genera Rhynchophorus and Dynamis (Coleoptera: Curculionidae). Ph.D. Thesis, Oregon State University, Corvallis, OR, USA, 1966. Available online: https://hdl.handle.net/1957/10673 (accessed on 14 June 2021).
- Monge, J.E.; Chinchilla, C.M.; Wang, A. Studies on the etiology of the crown disease / spear rot syndrome in oil palm. ASD. Oil Palm Papers 1993, 7, 1–16. [Google Scholar]
- Hafizi, R.; Salleh, B.; Latiffah, Z. Morphological and molecular characterization of Fusarium solani and F. oxysporum associated with crown disease of oil palm. Braz. J. Microbiol. 2013, 44, 959–968. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Flood, J.; Keenan, L.; Wayne, S.; Hasan, Y. Studies on oil palm trunks as sources of infection in the field. Mycopathologia 2005, 159, 101–107. [Google Scholar] [CrossRef]
- Chung, G.F. Effect of pests and diseases on oil palm yield. In Palm Oil: Production, Processing, Characterization, and Uses, 1st ed.; Oi-Ming, L., Chin-Ping, T., Casimir, C.A., Eds.; Academic Press and AOCS Press: Beltsville, MD, USA, 2012; pp. 163–210. [Google Scholar] [CrossRef]
- Paterson, R.R.M. Future scenarios for oil palm mortality and infection by Phytophthora palmivora in Colombia, Ecuador and Brazil, extrapolated to Malaysia and Indonesia. Phytoparasitica 2020, 48, 513–523. [Google Scholar] [CrossRef]
- Milosavljević, I.; Hoddle, C.D.; Mafra-Neto, A.; Gómez-Marco, F.; Hoddle, M.S. Use of digital video cameras to determine the efficacy of two trap types for capturing Rhynchophorus palmarum (Coleoptera: Curculionidae). J. Econ. Entomol. 2020, 113, 3028–3031. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Year | Precipitation (mm) | Temperature (°C) | Wind Speed (m∙s−1) | Insolation (MJ m−2 d−1) | RH (%) | ||
|---|---|---|---|---|---|---|---|
| Maximum | Minimum | Average | |||||
| 2016 | 1480.0 | 29.6 | 20.2 | 24.0 | 4.1 | 3.7 | 87.3 |
| 2017 | 1662.1 | 28.7 | 20.0 | 23.5 | 3.6 | 3.9 | 87.1 |
| 2018 | 1257.9 | 29.4 | 19.9 | 23.7 | 3.8 | 3.7 | 86.4 |
| 2019 | 1515.4 | 29.1 | 20.3 | 23.8 | 3.8 | 3.7 | 87.3 |
| 2020 | 1398.3 | 29.1 | 20.0 | 23.7 | 3.7 | 3.7 | 87.1 |
| Average | 1462.7 | 29.2 | 20.1 | 23.7 | 3.8 | 3.7 | 87.0 |
| Model | Y0 | r | R2 | p-Value | Mean Square of the Remainder | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| INIAP TENERA | IRHO | INIAP TENERA | IRHO | INIAP TENERA | IRHO | INIAP TENERA | IRHO | INIAP TENERA | IRHO | |
| Exponential | 0.02111 | 0.00210 * | 0.39984 | 0.20180 * | 0.89765 | 0.79372 | 0.00083 | 0.00000 | 0.18604 | 0.45168 |
| Gompertz | 0.00000 | 0.00000 | 0.51861 | 0.14785 | 0.91875 | 0.76992 | 0.00772 | 0.27990 | 0.18906 | 0.52056 |
| Logistic | 0.00000 | 0.00003 | 2.01840 | 0.39380 * | 0.98962 | 0.80622 | 0.00126 | 0.00001 | 0.02541 | 0.42430 |
| Meteorological Variable | r | p-Value |
|---|---|---|
| Precipitation | −0.2437 | 0.0605 |
| Relative humidity | −0.1084 | 0.4096 |
| Growing degree days | 0.1651 | 0.2075 |
| Maximum temperature | 0.2244 | 0.0847 |
| Minimum temperature | −0.2324 | 0.0739 |
| Mean temperature | 0.0872 | 0.5078 |
| Water surplus | 0.0629 | 0.6333 |
| Solar radiation | −0.1700 | 0.1940 |
| Variables | β0 | β1 | β2 | R2 | p-Value |
|---|---|---|---|---|---|
| Fruit bunches yield | 4.956 | −0.0163 | −0.0147 | 0.61 | 7.967 × 10−7 |
| Net income | 17.44 | −0.1173 | −0.1800 | 0.78 | 5.617 × 10−13 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guamani-Quimis, L.A.; Solís-Bowen, A.L.; Portalanza, D.; Garcés-Fiallos, F.R. Can Mathematical Models Describe Spear Rot Progress in Oil Palm Trees? A Five-Year Black Weevil-Disease Assessment from Ecuador. Agriculture 2022, 12, 257. https://doi.org/10.3390/agriculture12020257
Guamani-Quimis LA, Solís-Bowen AL, Portalanza D, Garcés-Fiallos FR. Can Mathematical Models Describe Spear Rot Progress in Oil Palm Trees? A Five-Year Black Weevil-Disease Assessment from Ecuador. Agriculture. 2022; 12(2):257. https://doi.org/10.3390/agriculture12020257
Chicago/Turabian StyleGuamani-Quimis, Lenin A., Ariolfo Leonardo Solís-Bowen, Diego Portalanza, and Felipe R. Garcés-Fiallos. 2022. "Can Mathematical Models Describe Spear Rot Progress in Oil Palm Trees? A Five-Year Black Weevil-Disease Assessment from Ecuador" Agriculture 12, no. 2: 257. https://doi.org/10.3390/agriculture12020257
APA StyleGuamani-Quimis, L. A., Solís-Bowen, A. L., Portalanza, D., & Garcés-Fiallos, F. R. (2022). Can Mathematical Models Describe Spear Rot Progress in Oil Palm Trees? A Five-Year Black Weevil-Disease Assessment from Ecuador. Agriculture, 12(2), 257. https://doi.org/10.3390/agriculture12020257