
Variability in Cadmium Uptake in Common Wheat under Cadmium Stress: Impact of Genetic Variation and Silicon Supplementation




Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials and Experimental Design
2.2. Biomass and Root Morphology
2.3. Cd Concentration and Subcellular Distribution in Shoot and Root
2.4. Apoplastic Bypass Flow
2.5. Glutathione in Shoot and Root
2.6. Statistical Analysis
3. Results
3.1. Shoot and Root Biomass and Root Morphology
3.2. Cd Concentration and Subcellular Distribution in Shoot and Root
3.3. Glutathione (GSH) and Glutathione Disulfide (GSSG)
3.4. Translocation Factor, Cd Uptake, and Apoplastic Bypass Flow
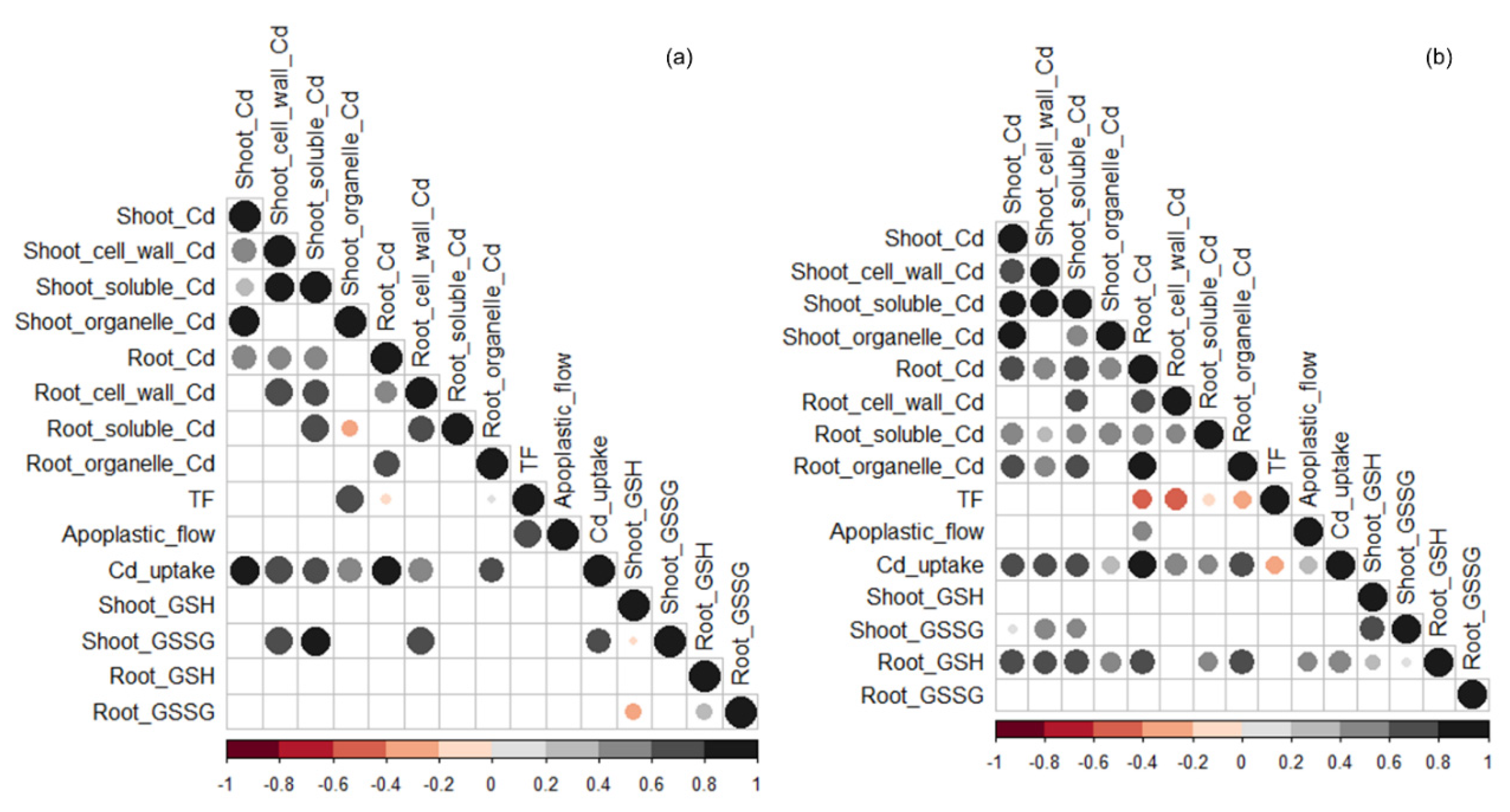
3.5. Correlations
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Centers for Disease Control and Prevention. Cadmium Toxicity; Centers for Disease Control and Prevention: Atlanta, GA, USA, 2011. [Google Scholar]
- Greger, M.; Löfstedt, M. Comparison of Uptake and Distribution of Cadmium in Different Cultivars of Bread and Durum Wheat. Crop Sci. 2004, 44, 501–507. [Google Scholar] [CrossRef]
- Jafarnejadi, A.R.; Homaee, M.; Sayyad, G.; Bybordi, M. Large Scale Spatial Variability of Accumulated Cadmium in the Wheat Farm Grains. Soil Sediment Contam. 2011, 20, 98–113. [Google Scholar] [CrossRef]
- Clemens, S.; Aarts, M.G.; Thomine, S.; Verbruggen, N. Plant Science: The Key to Preventing Slow Cadmium Poisoning. Trends Plant Sci. 2013, 18, 92–99. [Google Scholar] [CrossRef] [PubMed]
- Dong, Q.; Xu, P.; Wang, Z. Differential Cadmium Distribution and Translocation in Roots and Shoots Related to Hyper-Tolerance between Tall Fescue and Kentucky Bluegrass. Front. Plant Sci. 2017, 8, 113. [Google Scholar] [CrossRef] [Green Version]
- Uraguchi, S.; Mori, S.; Kuramata, M.; Kawasaki, A.; Arao, T.; Ishikawa, S. Root-to-Shoot Cd Translocation via the Xylem is the Major Process Determining Shoot and Grain Cadmium Accumulation in Rice. J. Exp. Bot. 2009, 60, 2677–2688. [Google Scholar] [CrossRef] [Green Version]
- Uraguchi, S.; Fujiwara, T. Rice Breaks Ground for Cadmium-Free Cereals. Curr. Opin. Plant Biol. 2013, 16, 328–334. [Google Scholar] [CrossRef]
- Gong, J.-M.; Lee, D.A.; Schroeder, J.I. Long-Distance Root-to-Shoot Transport of Phytochelatins and Cadmium in Arabidopsis. Proc. Natl. Acad. Sci. USA 2003, 100, 10118–10123. [Google Scholar] [CrossRef] [Green Version]
- Xin, J.; Huang, B.; Liu, A.; Zhou, W.; Liao, K. Identification of Hot Pepper Cultivars Containing Low Cd Levels after Growing on Contaminated Soil: Uptake and Redistribution to the Edible Plant Parts. Plant Soil 2013, 373, 415–425. [Google Scholar] [CrossRef]
- Harris, N.S.; Taylor, G.J. Cadmium Uptake and Partitioning in Durum Wheat during Grain Filling. BMC Plant Biol. 2013, 13, 103. [Google Scholar] [CrossRef] [Green Version]
- Perrier, F.; Yan, B.; Candaudap, F.; Pokrovsky, O.S.; Gourdain, E.; Meleard, B.; Bussiere, S.; Coriou, C.; Robert, T.; Nguyen, C. Variability in Grain Cadmium Concentration among Durum Wheat Cultivars: Impact of Aboveground Biomass Partitioning. Plant Soil 2016, 404, 307–320. [Google Scholar] [CrossRef] [Green Version]
- Lu, M.; Yu, S.; Lian, J.; Wang, Q.; He, Z.; Feng, Y.; Yang, X. Physiological and Metabolomics Responses of Two Wheat (Triticum Aestivum L.) Genotypes Differing in Grain Cadmium Accumulation. Sci. Total Environ. 2021, 769, 145345. [Google Scholar] [CrossRef] [PubMed]
- Parrotta, L.; Guerriero, G.; Sergeant, K.; Cai, G.; Hausman, J.-F. Target or Barrier? The Cell Wall of Early-and Later-Diverging Plants vs Cadmium Toxicity: Differences in the Response Mechanisms. Front. Plant Sci. 2015, 6, 133. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wei, W.; Peng, H.; Xie, Y.; Wang, X.; Huang, R.; Chen, H.; Ji, X. The Role of Silicon in Cadmium Alleviation by Rice Root Cell Wall Retention and Vacuole Compartmentalization under Different Durations of Cd Exposure. Ecotoxicol. Environ. Saf. 2021, 226, 112810. [Google Scholar] [CrossRef] [PubMed]
- Hu, X.; Li, T.; Xu, W.; Chai, Y. Distribution of Cadmium in Subcellular Fraction and Expression Difference of Its Transport Genes among Three Cultivars of Pepper. Ecotoxicol. Environ. Saf. 2021, 216, 112182. [Google Scholar] [CrossRef] [PubMed]
- Luo, J.-S.; Zhang, Z. Mechanisms of Cadmium Phytoremediation and Detoxification in Plants. Crop J. 2021, 9, 521–529. [Google Scholar] [CrossRef]
- Farooq, M.A.; Detterbeck, A.; Clemens, S.; Dietz, K.-J. Silicon-Induced Reversibility of Cadmium Toxicity in Rice. J. Exp. Bot. 2016, 67, 3573–3585. [Google Scholar] [CrossRef]
- Nakamura, S.; Suzui, N.; Yin, Y.-G.; Ishii, S.; Fujimaki, S.; Kawachi, N.; Rai, H.; Matsumoto, T.; Sato-Izawa, K.; Ohkama-Ohtsu, N. Effects of Enhancing Endogenous and Exogenous Glutathione in Roots on Cadmium Movement in Arabidopsis Thaliana. Plant Sci. 2020, 290, 110304. [Google Scholar] [CrossRef]
- Li, G.-Z.; Chen, S.-J.; Li, N.-Y.; Wang, Y.-Y.; Kang, G.-Z. Exogenous Glutathione Alleviates Cadmium Toxicity in Wheat by Influencing the Absorption and Translocation of Cadmium. Bull. Environ. Contam. Toxicol. 2021, 107, 320–326. [Google Scholar] [CrossRef]
- Hasanuzzaman, M.; Nahar, K.; Anee, T.I.; Fujita, M. Exogenous Silicon Attenuates Cadmium-Induced Oxidative Stress in Brassica Napus L. by Modulating AsA-GSH Pathway and Glyoxalase System. Front. Plant Sci. 2017, 8, 1061. [Google Scholar] [CrossRef]
- Jian, M.; Zhang, D.; Wang, X.; Wei, S.; Zhao, Y.; Ding, Q.; Han, Y.; Ma, L. Differential Expression Pattern of the Proteome in Response to Cadmium Stress Based on Proteomics Analysis of Wheat Roots. BMC Genom. 2020, 21, 343. [Google Scholar] [CrossRef]
- Wu, J.; Geilfus, C.-M.; Pitann, B.; Mühling, K.-H. Silicon-Enhanced Oxalate Exudation Contributes to Alleviation of Cadmium Toxicity in Wheat. Environ. Exp. Bot. 2016, 131, 10–18. [Google Scholar] [CrossRef]
- Greger, M.; Landberg, T. Role of Rhizosphere Mechanisms in Cd Uptake by Various Wheat Cultivars. Plant Soil 2008, 312, 195–205. [Google Scholar] [CrossRef]
- Shi, X.; Zhang, C.; Wang, H.; Zhang, F. Effect of Si on the Distribution of Cd in Rice Seedlings. Plant Soil 2005, 272, 53–60. [Google Scholar] [CrossRef]
- Shi, G.; Cai, Q.; Liu, C.; Wu, L. Silicon Alleviates Cadmium Toxicity in Peanut Plants in Relation to Cadmium Distribution and Stimulation of Antioxidative Enzymes. Plant Growth Regul. 2010, 61, 45–52. [Google Scholar] [CrossRef]
- Farooq, M.A.; Ali, S.; Hameed, A.; Ishaque, W.; Mahmood, K.; Iqbal, Z. Alleviation of Cadmium Toxicity by Silicon Is Related to Elevated Photosynthesis, Antioxidant Enzymes; Suppressed Cadmium Uptake and Oxidative Stress in Cotton. Ecotoxicol. Environ. Saf. 2013, 96, 242–249. [Google Scholar] [CrossRef]
- Greger, M.; Kabir, A.H.; Landberg, T.; Maity, P.J.; Lindberg, S. Silicate Reduces Cadmium Uptake into Cells of Wheat. Environ. Pollut. 2016, 211, 90–97. [Google Scholar] [CrossRef]
- Hussain, I.; Ashraf, M.A.; Rasheed, R.; Asghar, A.; Sajid, M.A.; Iqbal, M. Exogenous Application of Silicon at the Boot Stage Decreases Accumulation of Cadmium in Wheat (Triticum Aestivum L.) Grains. Braz. J. Bot. 2015, 38, 223–234. [Google Scholar] [CrossRef]
- Naeem, A.; Ghafoor, A.; Farooq, M. Suppression of Cadmium Concentration in Wheat Grains by Silicon Is Related to Its Application Rate and Cadmium Accumulating Abilities of Cultivars. J. Sci. Food Agric. 2015, 95, 2467–2472. [Google Scholar] [CrossRef]
- Liang, X.; Strawn, D.G.; Chen, J.; Marshall, J. Variation in Cadmium Accumulation in Spring Wheat Cultivars: Uptake and Redistribution to Grain. Plant Soil 2017, 421, 219–231. [Google Scholar] [CrossRef]
- Zadoks, J.C.; Chang, T.T.; Konzak, C.F. A Decimal Code for the Growth Stages of Cereals. Weed Res. 1974, 14, 415–421. [Google Scholar] [CrossRef]
- Cornu, J.-Y.; Bakoto, R.; Bonnard, O.; Bussiere, S.; Coriou, C.; Sirguey, C.; Sterckeman, T.; Thunot, S.; Visse, M.I.; Nguyen, C. Cadmium Uptake and Partitioning during the Vegetative Growth of Sunflower Exposed to Low Cd2+ Concentrations in Hydroponics. Plant Soil 2016, 404, 263–275. [Google Scholar] [CrossRef]
- Huang, C.-Y.L.; Schulte, E.E. Digestion of Plant Tissue for Analysis by ICP Emission Spectroscopy. Commun. Soil Sci. Plant Anal. 1985, 16, 943–958. [Google Scholar] [CrossRef]
- Weigel, H.J.; Jäger, H.J. Subcellular Distribution and Chemical Form of Cadmium in Bean Plants. Plant Physiol. 1980, 65, 480–482. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yeo, A.R.; Yeo, M.E.; Flowers, T.J. The Contribution of an Apoplastic Pathway to Sodium Uptake by Rice Roots in Saline Conditions. J. Exp. Bot. 1987, 38, 1141–1153. [Google Scholar] [CrossRef]
- Dempsey, R.W.; Merchant, A.; Tausz, M. Differences in Ascorbate and Glutathione Levels as Indicators of Resistance and Susceptibility in Eucalyptus Trees Infected with Phytophthora Cinnamomi. Tree Physiol. 2012, 32, 1148–1160. [Google Scholar] [CrossRef] [Green Version]
- R Core Team. R: A Language and Environment for Statistical Computing; Vienna R Core Team: Vienna, Austria, 2013. [Google Scholar]
- Courbot, M.; Willems, G.; Motte, P.; Arvidsson, S.; Roosens, N.; Saumitou-Laprade, P.; Verbruggen, N. A Major Quantitative Trait Locus for Cadmium Tolerance in Arabidopsis Halleri Colocalizes with HMA4, a Gene Encoding a Heavy Metal ATPase. Plant Physiol. 2007, 144, 1052–1065. [Google Scholar] [CrossRef] [Green Version]
- Sasaki, A.; Yamaji, N.; Yokosho, K.; Ma, J.F. Nramp5 Is a Major Transporter Responsible for Manganese and Cadmium Uptake in Rice. Plant Cell 2012, 24, 2155–2167. [Google Scholar] [CrossRef] [Green Version]
- Takahashi, R.; Ishimaru, Y.; Shimo, H.; Ogo, Y.; Senoura, T.; Nishizawa, N.K.; Nakanishi, H. The OsHMA2 Transporter Is Involved in Root-to-Shoot Translocation of Zn and Cd in Rice. Plant Cell Environ. 2012, 35, 1948–1957. [Google Scholar] [CrossRef]
- Clemens, S. Toxic Metal Accumulation, Responses to Exposure and Mechanisms of Tolerance in Plants. Biochimie 2006, 88, 1707–1719. [Google Scholar] [CrossRef]
- Mendoza-Cózatl, D.G.; Butko, E.; Springer, F.; Torpey, J.W.; Komives, E.A.; Kehr, J.; Schroeder, J.I. Identification of High Levels of Phytochelatins, Glutathione and Cadmium in the Phloem Sap of Brassica Napus. A Role for Thiol-Peptides in the Long-Distance Transport of Cadmium and the Effect of Cadmium on Iron Translocation. Plant J. 2008, 54, 249–259. [Google Scholar] [CrossRef] [Green Version]
- Rizwan, M.; Meunier, J.-D.; Miche, H.; Keller, C. Effect of Silicon on Reducing Cadmium Toxicity in Durum Wheat (Triticum Turgidum L. Cv. Claudio W.) Grown in a Soil with Aged Contamination. J. Hazard. Mater. 2012, 209, 326–334. [Google Scholar] [CrossRef] [PubMed]
- Nwugo, C.C.; Huerta, A.J. Effects of Silicon Nutrition on Cadmium Uptake, Growth and Photosynthesis of Rice Plants Exposed to Low-Level Cadmium. Plant Soil 2008, 311, 73–86. [Google Scholar] [CrossRef]
- Qi, X.; Tam, N.F.; Li, W.C.; Ye, Z. The Role of Root Apoplastic Barriers in Cadmium Translocation and Accumulation in Cultivars of Rice (Oryza Sativa L.) with Different Cd-Accumulating Characteristics. Environ. Pollut. 2020, 264, 114736. [Google Scholar] [CrossRef]
- Ma, J.; Sheng, H.; Li, X.; Wang, L. ITRAQ-Based Proteomic Analysis Reveals the Mechanisms of Silicon-Mediated Cadmium Tolerance in Rice (Oryza Sativa) Cells. Plant Physiol. Biochem. 2016, 104, 71–80. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.-H.; Khan, A.L.; Kim, D.-H.; Lee, S.-Y.; Kim, K.-M.; Waqas, M.; Jung, H.-Y.; Shin, J.-H.; Kim, J.-G.; Lee, I.-J. Silicon Mitigates Heavy Metal Stress by Regulating P-Type Heavy Metal ATPases, Oryza Sativa Low Silicon Genes, and Endogenous Phytohormones. BMC Plant Biol. 2014, 14, 13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feng Shao, J.; Che, J.; Yamaji, N.; Fang Shen, R.; Feng Ma, J. Silicon Reduces Cadmium Accumulation by Suppressing Expression of Transporter Genes Involved in Cadmium Uptake and Translocation in Rice. J. Exp. Bot. 2017, 68, 5641–5651. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nwugo, C.C.; Huerta, A.J. The Effect of Silicon on the Leaf Proteome of Rice (Oryza Sativa L.) Plants under Cadmium-Stress. J. Proteome Res. 2011, 10, 518–528. [Google Scholar] [CrossRef] [PubMed]
- Ishimaru, Y.; Takahashi, R.; Bashir, K.; Shimo, H.; Senoura, T.; Sugimoto, K.; Ono, K.; Yano, M.; Ishikawa, S.; Arao, T. Characterizing the Role of Rice NRAMP5 in Manganese, Iron and Cadmium Transport. Sci. Rep. 2012, 2, 286. [Google Scholar] [CrossRef]
- Mendoza-Cózatl, D.G.; Jobe, T.O.; Hauser, F.; Schroeder, J.I. Long-Distance Transport, Vacuolar Sequestration, Tolerance, and Transcriptional Responses Induced by Cadmium and Arsenic. Curr. Opin. Plant Biol. 2011, 14, 554–562. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Treatment | Root Length | Root Surface Area | Root Volume | Root Average Diameter |
|---|---|---|---|---|
| m plant−1 | cm2 plant−1 | cm3 plant−1 | mm | |
| No Cd applied | ||||
| Cultivar | ||||
| UI Platinum | 24.70 B * | 234.3 B | 1.778 B | 0.301 |
| LCS Star | 28.39 A | 262.6 A | 1.953 A | 0.293 |
| Si level | ||||
| No Si applied | 26.6 | 253.3 | 1.938 | 0.302 |
| Si applied | 26.5 | 243.6 | 1.797 | 0.291 |
| Source of variance | ||||
| Cultivar | <0.001 | 0.001 | 0.037 | 0.243 |
| Si | 0.927 | 0.252 | 0.097 | 0.077 |
| Cultivar × Si | 0.592 | 0.538 | 0.525 | 0.833 |
| Cd applied | ||||
| Cultivar | ||||
| UI Platinum | 6.699 | 78.46 | 0.747 | 0.389 |
| LCS Star | 6.837 | 79.47 | 0.741 | 0.384 |
| Si level | ||||
| No Si applied | 4.188 B | 54.25 B | 0.56 | 0.415 A |
| Si applied | 9.348 A | 103.7 A | 0.925 | 0.359 B |
| Source of variance | ||||
| Cultivar | 0.770 | 0.830 | 0.881 | 0.521 |
| Si | <0.001 | <0.001 | <0.001 | <0.001 |
| Cultivar × Si | 0.130 | 0.060 | 0.028 | 0.780 |
| Treatment | Shoot | Root | TF | Cd Uptake per Unit Root Length | Apoplastic Bypass Flow | ||
|---|---|---|---|---|---|---|---|
| GSH | GSSG | GSH | GSSG | ||||
| nmol g−1 fw | nmol g−1 fw | % | µg m−1 | nmol PTS g−1 fw | |||
| No Cd applied | |||||||
| Cultivar | |||||||
| UI Platinum | 1046 | 527.2 | 8.391 | 1.282 | 7.174 | 0.078 | 13.34 A |
| LCS Star | 891.7 | 572.6 | 6.031 | 1.882 | 6.352 | 0.080 | 8.538 B |
| Si level | |||||||
| No Si applied | 905.6 | 562.5 | 10.63 A | 1.406 | 8.296A | 0.092 | 12.74 A |
| Si applied | 1033 | 603.1 | 3.79 B | 1.757 | 5.494 B | 0.068 | 9.137 B |
| Source of variance | |||||||
| Cultivar | 0.443 | 0.308 | 0.477 | 0.273 | 0.493 | 0.894 | 0.002 |
| Si | 0.546 | 0.953 | 0.050 | 0.516 | 0.026 | 0.120 | 0.015 |
| Si × Cultivar | 0.294 | 0.864 | 0.289 | 0.882 | 0.892 | 0.118 | 0.151 |
| Cd applied | |||||||
| Cultivar | |||||||
| UI Platinum | 916.6 B * | 583.4 B | 13.36 | 1.598 | 1.754 B | 27.85 | 2.710 |
| LCS Star | 1598 A | 756.0 A | 15.47 | 1.908 | 2.200 A | 26.59 | 3.075 |
| Si level | |||||||
| No Si applied | 1280 | 684.4 | 18.86 A | 3.050 | 1.677 B | 44.39 A | 4.214 A |
| Si applied | 1235 | 655.0 | 9.963 B | 2.846 | 2.302 A | 16.68 B | 1.571 B |
| Source of variance | |||||||
| Cultivar | 0.008 | 0.050 | 0.164 | 0.323 | 0.050 | 0.702 | 0.582 |
| Si | 0.850 | 0.730 | <0.001 | 0.153 | 0.010 | <0.001 | <0.001 |
| Si × Cultivar | 0.863 | 0.608 | 0.645 | 0.331 | 0.748 | 0.504 | 0.080 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, R.; Liang, X.; Strawn, D.G. Variability in Cadmium Uptake in Common Wheat under Cadmium Stress: Impact of Genetic Variation and Silicon Supplementation. Agriculture 2022, 12, 848. https://doi.org/10.3390/agriculture12060848
Yang R, Liang X, Strawn DG. Variability in Cadmium Uptake in Common Wheat under Cadmium Stress: Impact of Genetic Variation and Silicon Supplementation. Agriculture. 2022; 12(6):848. https://doi.org/10.3390/agriculture12060848
Chicago/Turabian StyleYang, Rui, Xi Liang, and Daniel G. Strawn. 2022. "Variability in Cadmium Uptake in Common Wheat under Cadmium Stress: Impact of Genetic Variation and Silicon Supplementation" Agriculture 12, no. 6: 848. https://doi.org/10.3390/agriculture12060848
APA StyleYang, R., Liang, X., & Strawn, D. G. (2022). Variability in Cadmium Uptake in Common Wheat under Cadmium Stress: Impact of Genetic Variation and Silicon Supplementation. Agriculture, 12(6), 848. https://doi.org/10.3390/agriculture12060848