
Impacts of Nitrogen and Phosphorus Fertilization on Biomass, Polyphenol Contents, and Essential Oil Yield and Composition of Vitex negundo Linn

Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Materials
2.2. Time and Location of Experiments
2.3. Experimental Design
2.4. Soil Sampling and Chemical Analysis
2.5. Harvesting and Processing of Plant Materials
2.6. Essential Oil Extraction
2.7. Essential Oil Composition Analysis
2.8. Statistical Analysis
3. Results
3.1. Soil Chemical Properties
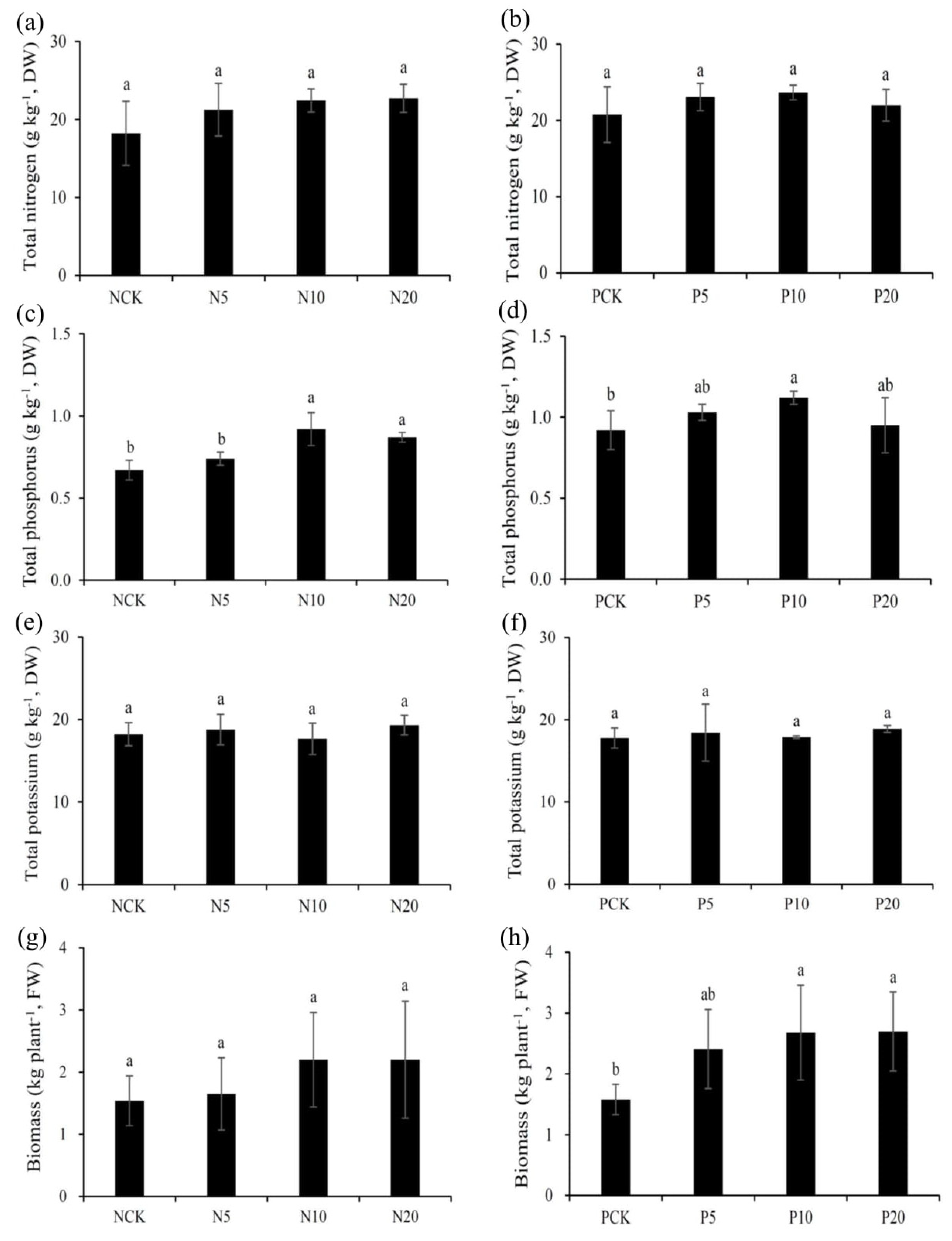
3.2. Plant Biomass Production
3.3. Nutrient (N, P, and K) Contents in Plant Parts
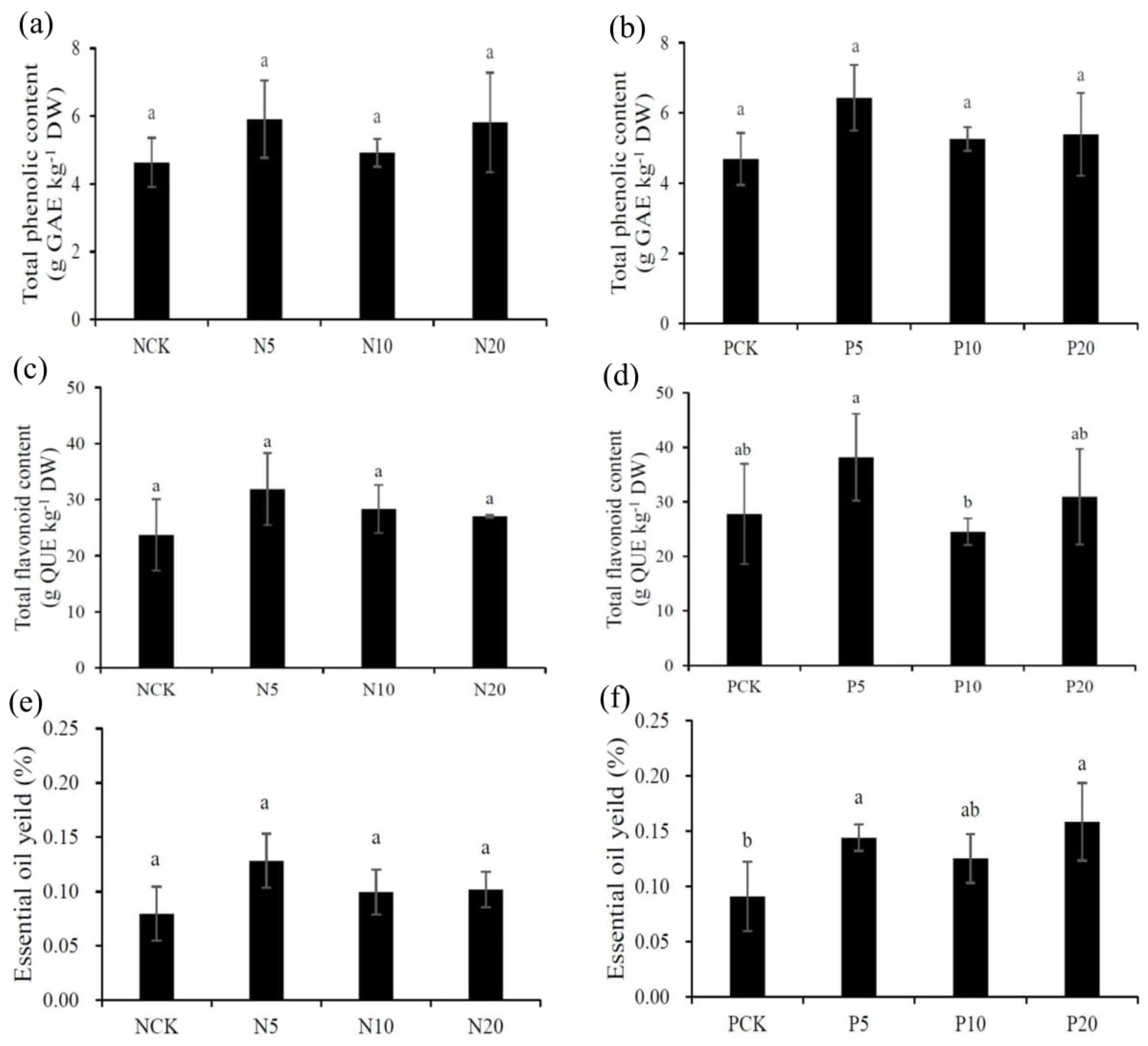
3.4. Total Phenolic and Total Flavonoid Contents
3.5. Essential Oil Yields
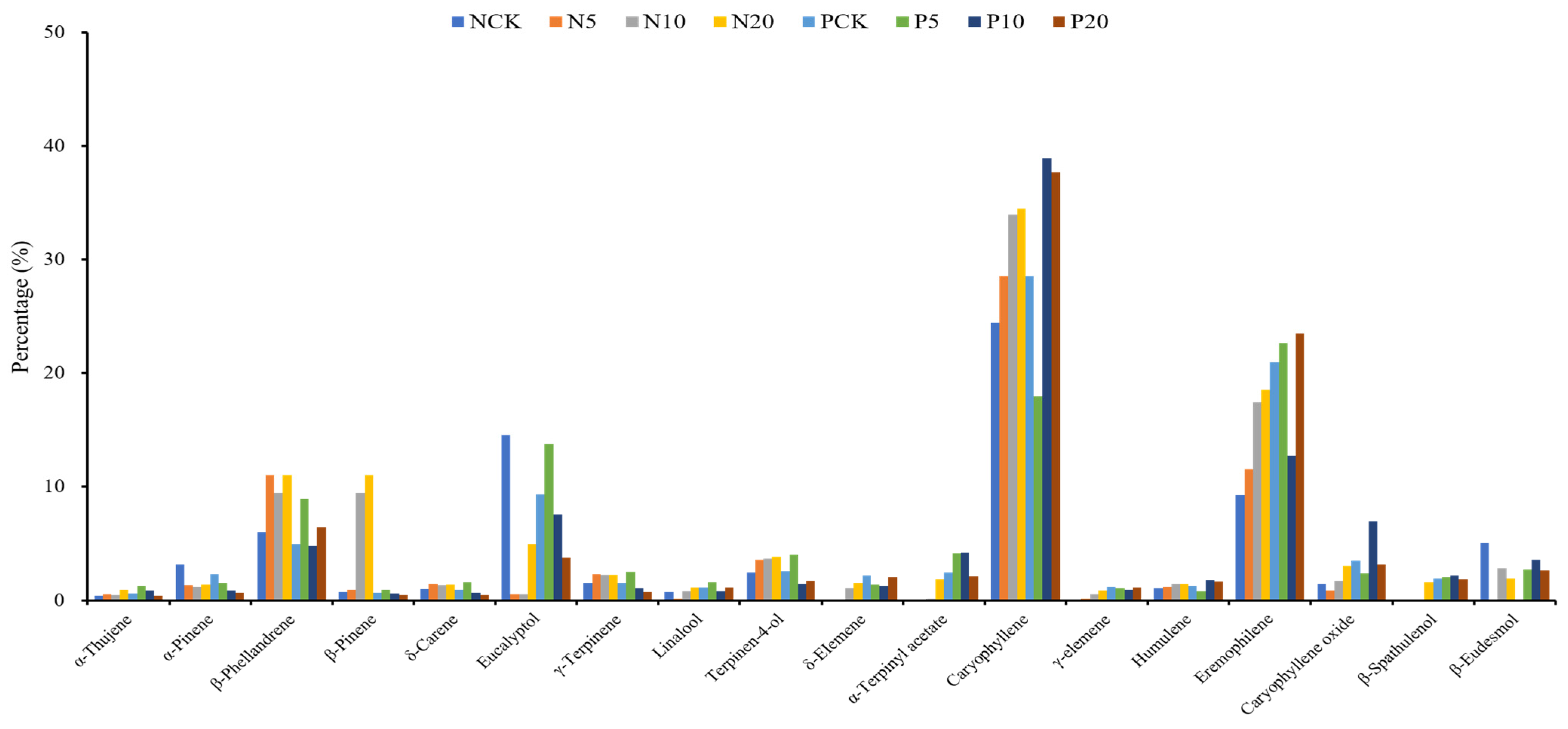
3.6. Essential Oil Composition
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Chen, S.L.; Yu, H.; Luo, H.M.; Wu, Q.; Li, C.F.; Steinmetz, A. Conservation and sustainable use of medicinal plants: Problems, progress, and prospects. Chin. Med. 2016, 11, 37. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arsenijević, J.; Drobac, M.; Šoštarić, I.; Jevđović, R.; Živković, J.; Ražić, S.; Moravčević, D.; Maksimović, Z. Comparison of essential oils and hydromethanol extracts of cultivated and wild growing Thymus pannonicus All. Ind. Crops Prod. 2019, 130, 162–169. [Google Scholar] [CrossRef]
- Pandey, V.; Patel, A.; Patra, D.D. Integrated nutrient regimes ameliorate crop productivity, nutritive value, antioxidant activity and volatiles in basil (Ocimum basilicum L.). Ind. Crops Prod. 2016, 87, 124–131. [Google Scholar] [CrossRef]
- Bistgani, Z.E.; Ataollah Siadat, S.; Bakhshandeh, A.; Ghasemi Pirbalouti, A.; Hashemi, M.; Maggi, F.; Reza Morshedloo, M. Application of combined fertilizers improves biomass, essential oil yield, aroma profile, and antioxidant properties of Thymus daenensis Celak. Ind. Crops Prod. 2018, 121, 434–440. [Google Scholar] [CrossRef]
- Chiu, S.M.; Lu, H.Y.; Liu, H.I. The growth and development of turmeric (Curcuma aromatica Salisb.) plants. II. Effects of N and K fertilization on plant growth and rhizome yield and quality. J. Agric. Res. China 1993, 42, 370–379. [Google Scholar]
- Chung, R.S.; Chen, C.C.; Ng, L.T. Nitrogen fertilization affects the growth performance, betaine and polysaccharide concentrations of Lycium barbarum. Ind. Crops Prod. 2010, 32, 650–655. [Google Scholar] [CrossRef]
- Chrysargyris, A.; Petropoulos, S.A.; Fernandes, Â.; Barros, L.; Tzortzakis, N.; Ferreira, I.C.F.R. Effect of phosphorus application rate on Mentha spicata L. grown in deep flow technique (DFT). Food Chem. 2019, 276, 84–92. [Google Scholar] [CrossRef] [Green Version]
- Ramezani, S.; Reza Rezaei, M.; Sotoudehnia, P. Improved growth, yield and essential oil content of basil grown under different levels of phosphorus sprays in the field. J. Appl. Biol. Sci. 2009, 3, 96–101. [Google Scholar]
- Erbaş, S.; Kucukyumuk, Z.; Baydar, H.; Erdal, I.; Şanli, A. Effects of different phosphorus doses on nutrient concentrations as well as yield and quality characteristics of lavandin (Lavandula × intermedia Emeric ex Loisel. var. Super). Turk. J. Field Crops 2017, 22, 32–38. [Google Scholar] [CrossRef] [Green Version]
- Trivino, M.G.; Johnson, C.B. Season has a major effect on the essential oil yield response to nutrient supply in Origanum majorana. J. Hortic. Sci. Biotechnol. 2000, 75, 520–527. [Google Scholar] [CrossRef]
- Nell, N.; Vötsch, M.; Vierheilig, H.; Steinkellner, S.; Zitterl-Eglseer, K.; Franz, C.; Novak, J. Effect of phosphorus uptake on growth and secondary metabolites of garden sage (Salvia officinalis L.). J. Sci. Food Agric. 2009, 89, 1090–1096. [Google Scholar] [CrossRef]
- Bansod, M.; Harle, U.N. Vitex negundo L: Phytochemical constituents, traditional uses and pharmacological properties: Comprehensive review. Pharmacologyonline 2009, 1, 286–302. [Google Scholar]
- Gill, B.S.; Mehra, R.; Navgeet; Kumar, S. Vitex negundo and its medicinal value. Mol. Biol. Rep. 2018, 45, 2925–2934. [Google Scholar] [CrossRef] [PubMed]
- Vishwanathan, A.S.; Basavaraju, R. A Review on Vitex negundo L.—A medicinally important plant. Eur. J. Biol. Sci. 2010, 3, 30–42. [Google Scholar]
- Khan, A.; Naz, S.; Farooq, U.; Shahid, M.; Ullah, I.; Ali, I.; Rauf, A.; Mabkhot, Y.N. Bioactive chromone constituents from Vitex negundo alleviate pain and inflammation. J. Pain Res. 2018, 11, 95–102. [Google Scholar] [CrossRef] [Green Version]
- Tiwari, O.P.; Tripathi, Y.B. Antioxidant properties of different fractions of Vitex negundo Linn. Food Chem. 2007, 100, 1170–1176. [Google Scholar] [CrossRef]
- Kamruzzaman, M.; Bari, S.M.N.; Faruque, S.M. In vitro and in vivo bactericidal activity of Vitex negundo leaf extract against diverse multidrug resistant enteric bacterial pathogens. Asian Pac. J. Trop. Med. 2013, 6, 352–359. [Google Scholar] [CrossRef] [Green Version]
- Khokra, S.L.; Prakash, O.; Jain, S.; Aneja, K.R.; Dhingra, Y. Essential oil composition and antibacterial studies of Vitex negundo Linn. extracts. Indian J. Pharm. Sci. 2008, 70, 522–526. [Google Scholar] [CrossRef] [Green Version]
- Prasad, E.M.; Mopuri, R.; Islam, M.S.; Kodidhela, L.D. Cardioprotective effect of Vitex negundo on isoproterenol-induced myocardial necrosis in wistar rats: A dual approach study. Biomed. Pharmacother. 2017, 85, 601–610. [Google Scholar] [CrossRef]
- Kadir, F.A.; Kassim, N.M.; Abdulla, M.A.; Yehye, W.A. Hepatoprotective role of ethanolic extract of Vitex negundo in thioacetamide-induced liver fibrosis in male rats. Evid. Based Complement. Alternat. Med. 2013, 2013, 739850. [Google Scholar] [CrossRef] [Green Version]
- Tandon, V.R.; Khajuria, V.; Kapoor, B.; Kour, D.; Gupta, S. Hepatoprotective activity of Vitex negundo leaf extract against anti-tubercular drugs induced hepatotoxicity. Fitoterapia 2008, 79, 533–538. [Google Scholar] [CrossRef] [PubMed]
- Villaseñor, I.M.; Lamadrid, M.R.A. Comparative anti-hyperglycemic potentials of medicinal plants. J. Ethnopharmacol. 2006, 104, 129–131. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.C.; Chang, T.Y.; Chang, L.Z.; Wang, H.F.; Yih, K.H.; Hsieh, W.Y.; Chang, T.M. Inhibition of melanogenesis versus antioxidant properties of essential oil extracted from leaves of Vitex negundo Linn and chemical composition analysis by GC-MS. Molecules 2012, 17, 3902–3916. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chattopadhyay, P.; Banerjee, S.; Pathak, M.P.; Agnihotri, A.; Karmakar, S.; Goyary, D.; Dhiman, S.; Veer, V. Acute and subchronic dermal toxicity of Vitex negundo essential oil. Cutan. Ocul. Toxicol. 2014, 33, 16–21. [Google Scholar] [CrossRef] [PubMed]
- Singh, V.; Dayal, R.; Bartley, J.P. Volatile constituents of Vitex negundo leaves. Planta Med. 1999, 65, 580–582. [Google Scholar] [CrossRef] [PubMed]
- Fidyt, K.; Fiedorowicz, A.; Strzadala, L.; Szumny, A. Beta-caryophyllene and beta-caryophyllene oxide-natural compounds of anticancer and analgesic properties. Cancer Med. 2016, 5, 3007–3017. [Google Scholar] [CrossRef]
- Gertsch, J.; Leonti, M.; Raduner, S.; Racz, I.; Chen, J.Z.; Xie, X.Q.; Altmann, K.H.; Karsak, M.; Zimmer, A. Beta-caryophyllene is a dietary cannabinoid. Proc. Natl. Acad. Sci. USA 2008, 105, 9099–9104. [Google Scholar] [CrossRef] [Green Version]
- Yuyama, K.T.; Fortkamp, D.; Abraham, W.R. Eremophilane-type sesquiterpenes from fungi and their medicinal potential. Biol. Chem. 2017, 399, 13–28. [Google Scholar] [CrossRef]
- Chou, C.H.; Yao, C. Phytochemical adaptation of coastal vegetation in Taiwan I. isolation, identification, and biological activities of compounds in Vitex negundo L. Bot. Bull. Acad. Sin. 1983, 24, 155–168. [Google Scholar]
- Huang, S.H.; Ng, L.T. Quantification of polyphenolic content and bioactive constituents of some commercial rice varieties in Taiwan. J. Food Compos. Anal. 2012, 26, 122–127. [Google Scholar] [CrossRef]
- Singh, A.; Sharma, P.; Garg, V.; Sharad, V. Extraction and analysis of essential oil of Nirgundi (Vitex negundo L.). Der. Pharm. Sin. 2011, 2, 262–266. [Google Scholar]
- Li, Z.G.; Zhang, R.H.; Xia, S.J.; Wang, L.; Liu, C.; Zhang, R.Q.; Fan, Z.H.; Chen, F.; Liu, Y. Interactions between N, P and K fertilizers affect the environment and the yield and quality of satsumas. Glob. Ecol. Conserv. 2019, 19, e00663. [Google Scholar] [CrossRef]
- Chrysargyris, A.; Panayiotou, C.; Tzortzakis, N. Nitrogen and phosphorus levels affected plant growth, essential oil composition and antioxidant status of lavender plant (Lavandula angustifolia Mill.). Ind. Crops Prod. 2016, 83, 577–586. [Google Scholar] [CrossRef]
- Ahmad, I.; Jabeen, N.; Ziaf, K.; Dole, J.M.; Khan, M.A.S.; Bakhtavar, M.A. Macronutrient application affects morphological, physiological, and seed yield attributes of Calendula officinalis L. Can. J. Plant Sci. 2017, 97, 906–916. [Google Scholar] [CrossRef] [Green Version]
- Lawlor, D.W.; Mengel, K.; Kirkby, E.A. Principles of plant nutrition. Ann. Bot. 2004, 93, 479–480. [Google Scholar] [CrossRef] [Green Version]
- Thomson, C.; Marschner, H.; Römheld, V. Effect of nitrogen fertilizer form on pH of the bulk soil and rhizosphere, and on the growth, phosphorus, and micronutrient uptake of bean. J. Plant Nutr. 1993, 16, 493–506. [Google Scholar] [CrossRef]
- Ruan, J.; Zhang, F.; Wong, M.H. Effect of nitrogen form and phosphorus source on the growth, nutrient uptake and rhizosphere soil property of Camellia sinensis L. Plant Soil 2000, 223, 65–73. [Google Scholar] [CrossRef]
- Kougan, G.B.; Tabopda, T.; Kuete, V.; Verpoorte, V. Simple phenols, phenolic acids, and related esters from the medicinal plants of africa. In Medicinal Plant Research in Africa; Kuete, V., Ed.; Elsevier: Oxford, UK, 2013; pp. 225–249. [Google Scholar]
- Olesińska, K.; Sugier, D.; Kaczmarski, Z. Yield and chemical composition of raw material from meadow Arnica (Arnica chamissonis Less.) depending on soil conditions and nitrogen fertilization. Agriculture 2021, 11, 810. [Google Scholar] [CrossRef]
- Heimler, D.; Romani, A.; Ieri, F. Plant polyphenol content, soil fertilization and agricultural management: A review. Eur. Food Res. Technol. 2017, 243, 1107–1115. [Google Scholar] [CrossRef]
- Radušienė, J.; Marksa, M.; Ivanauskas, L.; Jakštas, V.; Çalişkan, Ö.; Kurt, D.; Odabaş, M.S.; Çirak, C. Effect of nitrogen on herb production, secondary metabolites and antioxidant activities of Hypericum pruinatum under nitrogen application. Ind. Crop. Prod. 2019, 139, 111519. [Google Scholar] [CrossRef]
- Deng, B.; Li, Y.; Xu, D.; Ye, Q.; Liu, G. Nitrogen availability alters flavonoid accumulation in Cyclocarya paliurus via the effects on the internal carbon/nitrogen balance. Sci. Rep. 2019, 9, 2370. [Google Scholar] [CrossRef] [PubMed]
- Król, B.; Sęczyk, Ł.; Kołodziej, B.; Paszko, T. Biomass production, active substance content, and bioaccessibility of Greek oregano (Origanum vulgare ssp. hirtum (Link) Ietswaart) following the application of nitrogen. Ind. Crop. Prod. 2020, 148, 112271. [Google Scholar] [CrossRef]
- Sun, J.; Luo, H.; Jiang, Y.; Wang, L.; Xiao, C.; Weng, L. Influence of nutrient (NPK) factors on growth, and pharmacodynamic component biosynthesis of Atractylodes chinensis: An insight on acetyl-CoA carboxylase (ACC), 3-hydroxy-3-methylglutaryl-CoA reductase (HMGR), and farnesyl pyrophosphate synthase (FPPS) signaling responses. Front. Plant Sci. 2022, 13, 799201. [Google Scholar] [PubMed]
- Gershenzon, J. Metabolic costs of terpenoid accumulation in higher plants. J. Chem. Ecol. 1994, 20, 1281–1328. [Google Scholar] [CrossRef] [PubMed]
- Yang, D.; Du, X.; Liang, X.; Han, R.; Liang, Z.; Liu, Y.; Liu, F.; Zhao, J. Different roles of the mevalonate and methylerythritol phosphate pathways in cell growth and tanshinone production of Salvia miltiorrhiza hairy roots. PLoS ONE 2012, 7, e46797. [Google Scholar] [CrossRef]
- Roberts, S.C. Production and engineering of terpenoids in plant cell culture. Nat. Chem. Biol. 2007, 3, 387–395. [Google Scholar] [CrossRef]
- Padalia, R.C.; Verma, R.S.; Chauhan, A.; Chanotiya, C.S.; Thul, S. Phytochemical diversity in essential oil of Vitex negundo L. populations from India. Rec. Nat. Prod. 2016, 10, 452–464. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| Treatments | pH | EC | OM | TN | P | K |
|---|---|---|---|---|---|---|
| Before | 4.6 ± 0.3 | 0.3 ± 0.1 | 31.9 ± 1.3 | 1.1 ± 0.8 | 19.4 ± 3.1 | 130.1 ± 24.3 |
| After nitrogen fertilization | ||||||
| NCK (Control) | 4.4 ± 0.2 b | 0.3 ± 0.1 b | 45.6 ± 2.1 a * | 1.2 ± 0.2 c | 45.0 ± 1.2 c * | 149.1 ± 21.9 b |
| N5 | 4.7 ± 0.2 b | 0.7 ± 0.2 a | 49.3 ± 9.8 a | 1.6 ± 0.1 b | 91.9 ± 3.2 b | 378.4 ± 39.7 a |
| N10 | 5.2 ± 0.2 a | 0.7 ± 0.3 a | 44.0 ± 4.4 a | 1.7 ± 0.2 b | 102.6 ± 7.8 a | 319.2 ± 41.3 a |
| N20 | 5.4 ± 0.5 a | 0.8 ± 0.4 a | 49.6 ± 7.4 a | 2.6 ± 0.6 a | 111.5 ± 5.7 a | 397.9 ± 72.2 a |
| After phosphorus fertilization | ||||||
| PCK (Control) | 4.3 ± 0.1 b | 0.5 ± 0.2 b | 43.1 ± 4.4 a * | 1.4 ± 0.2 c | 33.5 ± 1.4 d * | 202.5 ± 46.2 b |
| P5 | 4.7 ± 0.3 b | 0.6 ± 0.2 b | 52.4 ± 5.7 a | 2.1 ± 0.3 a | 51.8 ± 5.7 c | 410.9 ± 73.8 a |
| P10 | 5.2 ± 0.2 a | 0.6 ± 0.1 b | 53.8 ± 12.1 a | 1.9 ± 0.2 a | 73.8 ± 9.8 b | 338.2 ± 66.0 a |
| P20 | 5.1 ± 0.3 a | 1.8 ± 0.2 a | 55.2 ± 6.8 a | 1.8 ± 0.2 a | 143.5 ± 2.7 a | 335.1 ± 35.3 a |
| No | Components | RT | NCK | N5 | N10 | N20 | PCK | P5 | P10 | P20 |
|---|---|---|---|---|---|---|---|---|---|---|
| 1 | α-Thujene | 7.24 | 0.37 | 0.50 | 0.49 | 0.92 | 0.62 | 1.23 | 0.86 | 0.39 |
| 2 | α-Pinene | 7.47 | 3.12 | 1.30 | 1.18 | 1.36 | 2.32 | 1.49 | 0.88 | 0.67 |
| 3 | β-Phellandrene | 8.81 | 5.97 | 11.01 | 9.43 | 10.96 | 4.90 | 8.91 | 4.78 | 6.44 |
| 4 | β-Pinene | 8.97 | 0.75 | 0.92 | 9.43 | 0.93 | 0.64 | 0.92 | 0.56 | 0.48 |
| 5 | 1-Octen-3-ol | 9.05 | - | - | - | 0.06 | - | - | - | - |
| 6 | β-Myrcene | 9.38 | 0.67 | 0.37 | 0.25 | 0.30 | 0.49 | 0.48 | 0.23 | - |
| 7 | β-Thujene | 9.39 | - | - | - | - | - | - | - | 0.27 |
| 8 | α-Phellandrene | 9.98 | 0.12 | - | - | 0.06 | 0.13 | 0.09 | - | - |
| 9 | δ-Carene | 10.37 | 0.96 | 1.42 | 1.31 | 1.40 | 0.92 | 1.58 | 0.65 | 0.45 |
| 10 | p-Cymene | 10.64 | 0.22 | 0.16 | 0.20 | 0.22 | 0.20 | 0.30 | 0.22 | 0.08 |
| 11 | D-Limonene | 10.83 | - | 0.23 | 0.21 | 0.48 | - | 0.99 | 0.60 | 0.25 |
| 12 | Sabinene | 10.88 | - | - | - | - | - | - | - | 0.18 |
| 13 | Eucalyptol | 10.95 | 14.52 | 0.54 | 0.52 | 4.92 | 9.30 | 13.76 | 7.53 | 3.73 |
| 14 | β-Ocimene | 11.48 | 0.07 | 0.08 | 0.08 | 0.25 | - | 0.06 | - | - |
| 15 | γ-Terpinene | 11.91 | 1.52 | 2.30 | 2.21 | 2.24 | 1.52 | 2.48 | 1.04 | 0.74 |
| 16 | (Z)-β-Terpineol | 11.92 | - | - | - | 0.06 | - | - | - | - |
| 17 | 2-Carene | 12.92 | 3.23 | 0.07 | 0.09 | - | 0.37 | - | 0.10 | 0.17 |
| 18 | Methyl benzoate | 13.21 | - | - | - | - | - | - | 0.14 | - |
| 19 | Linalool | 13.48 | 0.73 | 0.16 | 0.76 | 1.10 | 1.14 | 1.55 | 0.76 | 1.13 |
| 20 | Fenchene | 14.39 | 0.09 | - | - | - | - | - | 0.06 | - |
| 21 | p-Menth-2-en-1-ol | 14.40 | - | 0.11 | 0.11 | 0.13 | 0.09 | 0.09 | - | - |
| 22 | Lavandulol | 15.90 | - | 1.29 | 0.35 | 0.19 | 0.24 | 0.12 | 0.30 | 0.60 |
| 23 | Myrcenol | 16.12 | - | - | - | 0.07 | - | - | - | 0.11 |
| 24 | Terpinen-4-ol | 16.53 | 2.41 | 3.52 | 3.69 | 3.79 | 2.54 | 4.02 | 1.43 | 1.70 |
| 25 | α-Terpineol | 17.05 | 0.85 | 0.17 | 0.19 | 0.40 | 0.97 | 1.01 | 0.47 | 0.39 |
| 26 | (R)-Lavandulyl acetate | 20.31 | 0.15 | - | 0.33 | - | 1.18 | 0.33 | 0.81 | 1.83 |
| 27 | Lavandulyl acetate | 20.32 | - | 1.42 | - | 0.63 | - | - | - | - |
| 28 | Pseudolimonen | 21.36 | - | - | - | - | - | 0.06 | 0.06 | - |
| 29 | (E)-β-Terpinolene | 22.04 | 0.16 | - | - | - | - | - | - | - |
| 30 | α-Terpinene | 22.05 | - | - | - | 0.12 | - | - | - | 0.14 |
| 31 | δ-EIemene | 22.17 | - | - | 1.07 | 1.51 | 2.14 | 1.37 | 1.24 | 2.02 |
| 32 | α-Terpinyl acetate | 22.54 | - | - | 0.13 | 1.80 | 2.45 | 4.14 | 4.21 | 2.11 |
| 33 | β-Bourbonene | 23.85 | - | - | - | - | - | 0.09 | - | - |
| 34 | β-Elemene | 24.05 | 0.18 | 0.10 | 0.39 | 0.34 | - | 0.34 | 0.27 | 0.44 |
| 35 | β-Caryophyllene | 25.15 | 24.41 | 28.52 | 33.91 | 34.48 | 28.48 | 17.91 | 38.92 | 37.66 |
| 36 | Artemisia triene | 25.38 | 0.14 | - | - | - | - | - | - | - |
| 37 | γ-Elemene | 25.39 | - | 0.15 | 0.50 | 0.86 | 1.18 | 1.03 | 0.91 | 1.14 |
| 38 | α-Funebrene | 25.77 | - | - | 0.08 | - | - | - | - | - |
| 39 | α-Muurolene | 25.78 | - | - | - | - | 0.26 | - | - | - |
| 40 | α-Ylangene | 25.79 | - | - | - | 0.17 | 0.23 | 0.29 | 0.17 | - |
| 41 | β-Patchoulene | 25.80 | - | - | - | - | - | - | - | 0.24 |
| 42 | α-Cubebene | 26.04 | - | - | - | - | - | 0.08 | - | - |
| 43 | Humulene | 26.26 | 1.05 | 1.15 | 1.44 | 1.47 | 1.27 | 0.81 | 1.78 | 1.61 |
| 44 | 4,5-Di-epi-aristolochene | 26.73 | - | - | - | - | 0.06 | 0.08 | - | 0.06 |
| 45 | cis-Muurola-3,5-diene | 26.95 | - | - | - | - | - | - | 0.24 | - |
| 46 | Isoledene | 26.96 | - | - | - | - | - | - | - | 0.27 |
| 47 | Cyclosativene | 26.98 | - | - | - | 0.16 | - | - | - | - |
| 48 | β-Copaene | 27.09 | 0.14 | 0.59 | 0.22 | 0.40 | 0.53 | 0.50 | - | - |
| 49 | D-Germacrene | 27.10 | - | - | - | - | - | - | 0.39 | 0.54 |
| 50 | Eremophilene | 27.35 | 9.20 | 11.50 | 17.41 | 18.51 | 20.93 | 22.61 | 12.68 | 23.44 |
| 51 | Selinene | 27.59 | - | - | 0.13 | - | 0.34 | 0.31 | 0.20 | - |
| 52 | Guaia-9,11-diene | 27.60 | - | - | - | 0.18 | - | - | - | 0.33 |
| 53 | α-Longipinene | 27.84 | - | - | - | - | - | 0.26 | - | - |
| 54 | δ-Cadinene | 28.28 | - | - | - | - | 0.12 | 0.07 | 0.14 | 0.09 |
| 55 | Elixene | 29.57 | - | - | - | - | 0.35 | - | - | 0.36 |
| 56 | cis-3-Hexenyl benzoate | 29.90 | 0.13 | 0.13 | 0.17 | 0.11 | 0.11 | - | 0.29 | 0.15 |
| 57 | Caryophyllene oxide | 30.30 | 1.41 | 0.86 | 1.68 | 3.01 | 3.49 | 2.35 | 6.90 | 3.15 |
| 58 | Isocamphane | 31.13 | - | - | - | - | - | - | - | 0.10 |
| 59 | Humulene-1,2-epoxide | 31.14 | - | - | - | - | - | - | 0.24 | - |
| 60 | α-Patchoulene | 31.36 | - | - | - | - | - | - | - | 0.18 |
| 61 | Bulnesol | 31.37 | - | - | - | - | - | 0.19 | - | - |
| 62 | Longifolene | 31.47 | - | - | - | - | - | - | 0.06 | - |
| 63 | Valencene | 31.53 | 0.19 | - | - | - | 0.13 | - | 0.14 | - |
| 64 | β-Nootkatol | 31.67 | - | - | 0.41 | - | - | - | - | - |
| 65 | β-Spathulenol | 31.68 | - | - | - | 1.54 | 1.93 | 2.05 | 2.17 | 1.81 |
| 66 | Aromandendrene | 31.85 | - | - | 0.10 | 0.07 | 0.08 | 2.05 | - | 0.11 |
| 67 | Caryophylla-4(12),8(13)-diene-5β-ol | 31.95 | 0.51 | - | - | - | - | - | - | - |
| 68 | Isoaromadendrene epoxide | 31.96 | - | - | - | - | 0.09 | - | 0.42 | 0.12 |
| 69 | β-Gurjurene | 32.03 | - | - | - | - | - | - | 0.17 | |
| 70 | α-Guaiene | 32.04 | 0.46 | - | - | - | - | 0.28 | - | - |
| 71 | α-Cadinol | 32.48 | - | 1.14 | - | - | 3.47 | - | - | - |
| 72 | β-Eudesmol | 32.49 | 5.05 | - | 2.83 | 1.89 | - | 2.66 | 3.53 | 2.61 |
| 73 | cis-Meta-mentha-2,8-diene | 32.92 | - | - | - | 0.23 | - | - | - | - |
| 74 | α-Selinene | 38.84 | 0.95 | 1.33 | 1.23 | - | - | - | - | - |
| 75 | 4-Quinolinol, 2,7,8-trimethyl | 40.08 | 1.94 | - | - | - | - | - | - | - |
| 76 | 2-Methyl-1-tripropylsilyloxypropan | 40.09 | - | - | 0.79 | - | - | - | - | - |
| 77 | 1-Indanone, 3,3,4,5,7-pentamethyl | 40.10 | - | 1.54 | - | - | - | - | - | - |
| 78 | 2,3-Dihydrothieno(2,3-b)quinoline | 40.18 | 0.93 | - | - | - | - | - | - | - |
| 79 | (E)-Farnesene epoxide | 41.03 | - | - | - | - | - | - | - | 0.08 |
| 80 | Pyridine,2-methyl-3-(2-propenyloxy) | 41.05 | - | 1.83 | 0.46 | - | - | - | 0.10 | - |
| 81 | Epimanool | 41.52 | - | 2.19 | - | - | - | - | - | - |
| 82 | Epimanoyl oxide | 41.92 | - | - | 0.13 | - | - | - | - | - |
| 83 | Isopimaradiene | 42.02 | - | - | - | 0.27 | - | - | - | - |
| 84 | 7-Methyl-1,2,3,5,8,8a-hexahydro-naphthalene | 42.65 | 2.04 | 12.19 | 0.99 | - | - | - | 1.02 | 0.23 |
| 85 | Abitatriene | 43.41 | 0.97 | 1.90 | 0.40 | - | - | - | 0.46 | - |
| 86 | p-Toluic acid,2-adamantyl ester | 44.08 | - | - | - | - | - | - | - | 0.29 |
| 87 | 1,8-Nonadiene-2,7-dimethyl-5-(methylethenyl) | 44.77 | 0.42 | - | - | - | - | - | - | - |
| Total | 86.03 | 90.69 | 95.30 | 97.59 | 95.21 | 98.94 | 97.96 | 99.06 | ||
| No. of compounds | 36 | 33 | 39 | 40 | 37 | 40 | 42 | 45 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Peng, L.-C.; Ng, L.-T. Impacts of Nitrogen and Phosphorus Fertilization on Biomass, Polyphenol Contents, and Essential Oil Yield and Composition of Vitex negundo Linn. Agriculture 2022, 12, 859. https://doi.org/10.3390/agriculture12060859
Peng L-C, Ng L-T. Impacts of Nitrogen and Phosphorus Fertilization on Biomass, Polyphenol Contents, and Essential Oil Yield and Composition of Vitex negundo Linn. Agriculture. 2022; 12(6):859. https://doi.org/10.3390/agriculture12060859
Chicago/Turabian StylePeng, Li-Chen, and Lean-Teik Ng. 2022. "Impacts of Nitrogen and Phosphorus Fertilization on Biomass, Polyphenol Contents, and Essential Oil Yield and Composition of Vitex negundo Linn" Agriculture 12, no. 6: 859. https://doi.org/10.3390/agriculture12060859
APA StylePeng, L. -C., & Ng, L. -T. (2022). Impacts of Nitrogen and Phosphorus Fertilization on Biomass, Polyphenol Contents, and Essential Oil Yield and Composition of Vitex negundo Linn. Agriculture, 12(6), 859. https://doi.org/10.3390/agriculture12060859