
From Ground to Grain: Tracing Phosphorus and Potassium in Flooded Rice Cultivar Grown on Histosols

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
3. Statistical Analysis
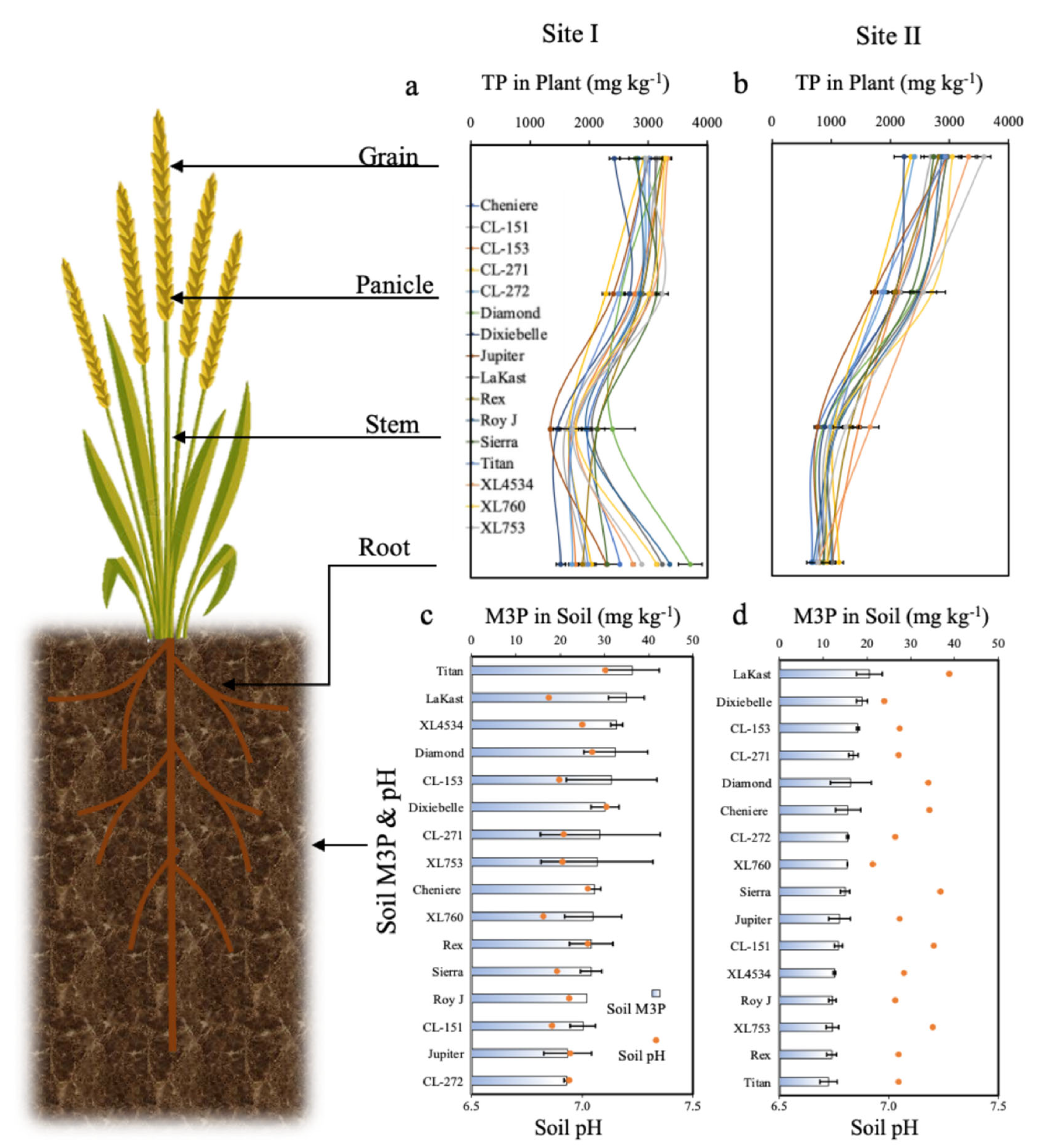
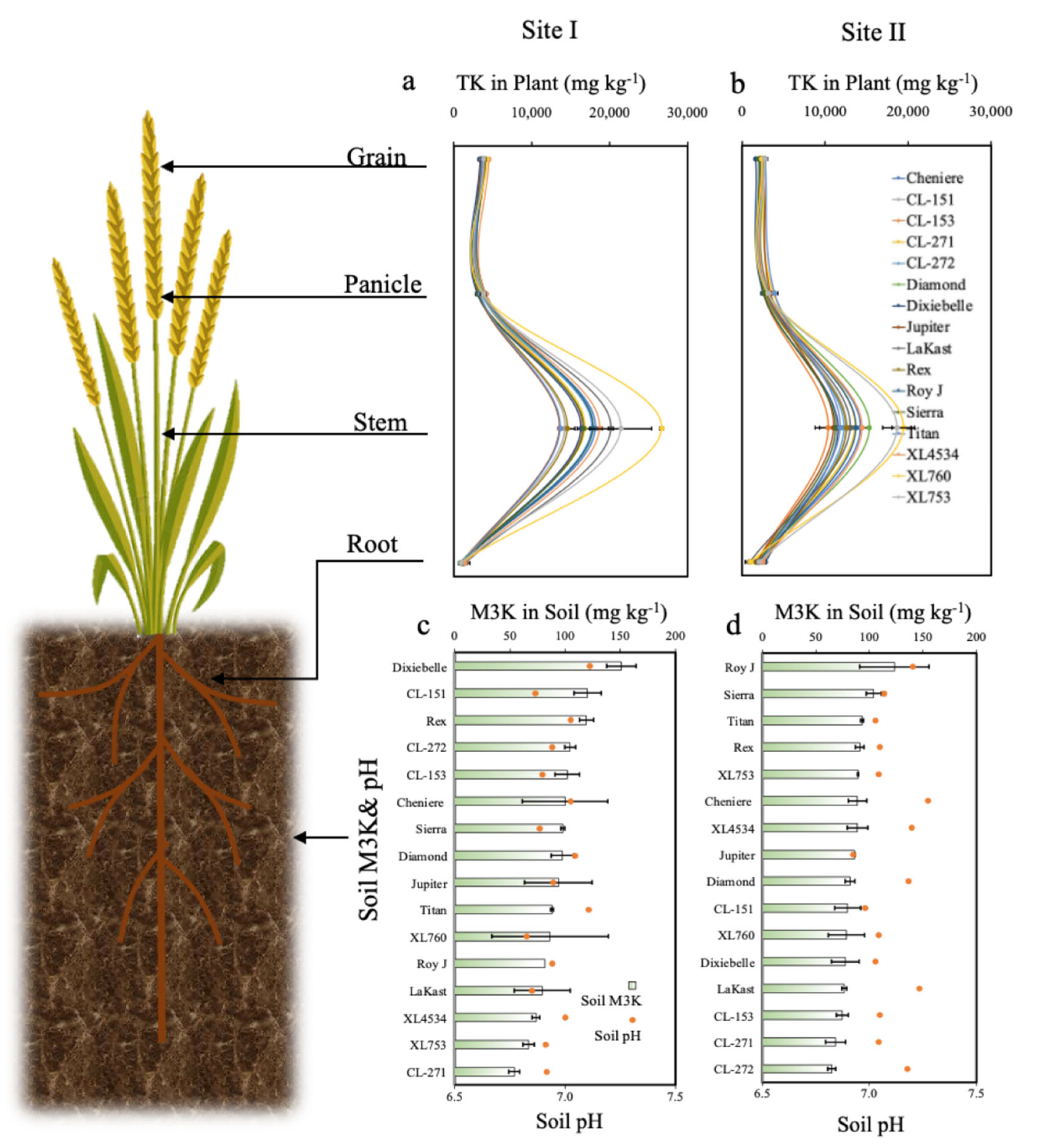
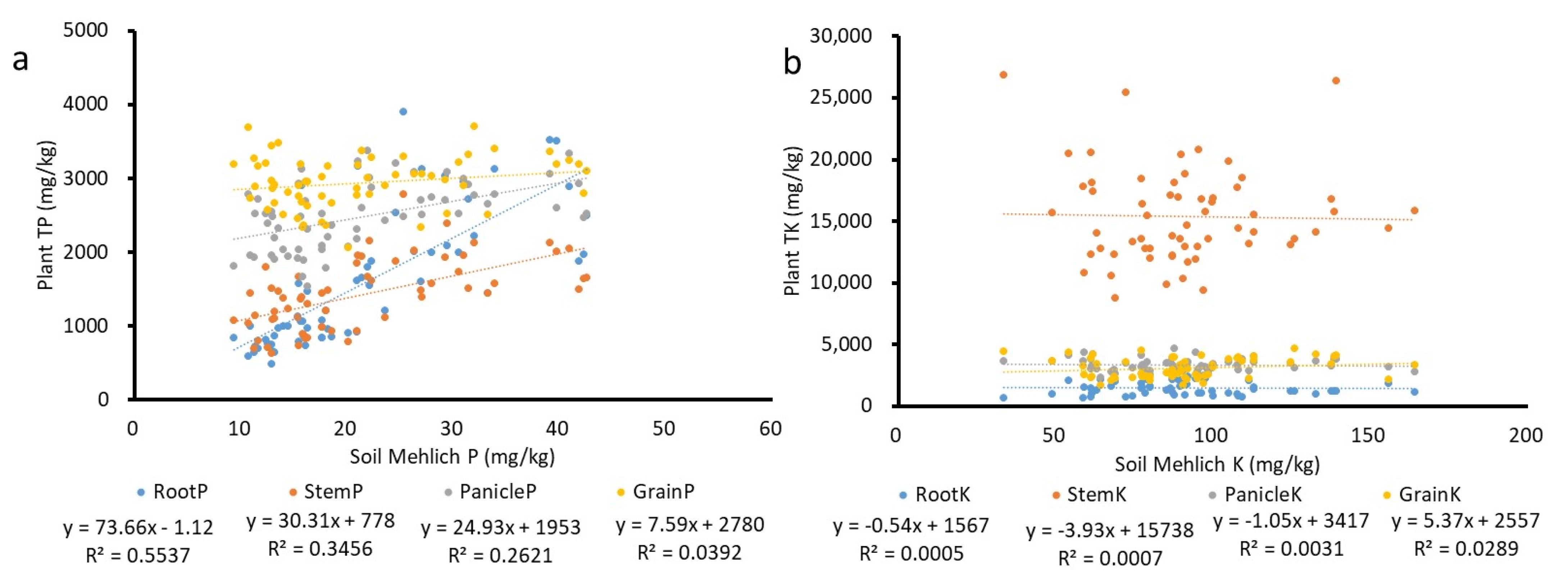
4. Results
4.1. Soil pH and Mehlich-3 Potassium and Phosphorus
4.2. Plant Total TP and TK
4.3. Yield
5. Discussion
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Fageria, N.K.; Baligar, V.C. Upland rice genotypes evaluation for phosphorus use efficiency. J. Plant Nutr. 1997, 20, 499–509. [Google Scholar] [CrossRef]
- Taiz, L.; Zeiger, E. Plant Physiology, 3rd ed.; Sinauer Associates: Sunderland, UK, 2003. [Google Scholar]
- Nath, M.; Tuteja, N. NPKS uptake, sensing, and signaling and miRNAs in plant nutrient stress. Protoplasma 2016, 253, 767–786. [Google Scholar] [CrossRef] [PubMed]
- Malvi, U.R. Interaction of micronutrients with major nutrients with special reference to potassium. Karnataka J. Agric. Sci. 2011, 24, 106–109. [Google Scholar]
- Maruyama, H.; Wasaki, J. Transgenic approaches for improving phosphorus use efficiency in plants. In Plant Macronutrient Use Efficiency; Elsevier: Amsterdam, The Netherlands, 2018; pp. 323–338. [Google Scholar]
- Maathuis, F.J. Plant Mineral Nutrients Methods and Protocols. In Methods in Molecular Biology; Walker, J.M., Ed.; Human Press: New Delhi, India, 2013; p. 290. [Google Scholar]
- Maathuis, F.J.; Podar, D. Uptake, distribution, and physiological functions of potassium, calcium, and magnesium. In The Molecular and Physiological Basis of Nutrient Use Efficiency in Crops; Hawkesford, M., Barraclough, P., Eds.; Wiley-Blackwell: New York, NY, USA, 2011; pp. 265–293. Available online: https://onlinelibrary.wiley.com/doi/book/10.1002/9780470960707 (accessed on 20 June 2022).
- Baffes, J.; Koh, W.C. Fertilizer Prices Expected to Remain Higher for Longer. World Bank Blogs. 2022. Available online: https://blogs.worldbank.org/opendata/fertilizer-prices-expected-remain-higher-longer (accessed on 20 June 2022).
- Lynch, J.P. Root phenes for enhanced soil exploration and phosphorus acquisition: Tools for future crops. Plant Physiol. 2011, 156, 1041–1049. [Google Scholar] [CrossRef] [PubMed]
- Haefele, S.M.; Nelson, A.; Hijmans, R.J. Soil quality and constraints in global rice production. Geoderma 2014, 235, 250–259. [Google Scholar] [CrossRef]
- Bhadha, J.; Khatiwada, R.; Galindo, S.; Xu, N.; Capasso, J. Evidence of soil health benefits of flooded rice compared to fallow practice. Sustain. Agric. Res. 2018, 7, 31–41. [Google Scholar] [CrossRef]
- Bhadha, J.H.; Trotta, L.; VanWeelden, M.T. Trends in Rice Production and Varieties in the Everglades Agricultural Area: SL439/SS653, 5/2016. EDIS 2016, 2016, 4. [Google Scholar]
- Hardke, J.T. Trends in Arkansas Rice Production, 2018; Arkansas Agricultural Experiment Station, University of Arkansas System: Fayetteville, AR, USA, 2019; pp. 15–24. [Google Scholar]
- De Datta, S.K.; Broadbent, F.E. Nitrogen-use efficiency of 24 rice genotypes on an N-deficient soil. Field Crops Res. 1990, 23, 81–92. [Google Scholar] [CrossRef]
- Somaweera, K.A.T.N.; Suriyagoda, L.D.B.; Sirisena, D.N.; De Costa, W.A.J.M. Growth, root adaptations, phosphorus and potassium nutrition of rice when grown under the co-limitations of phosphorus, potassium and moisture. J. Plant Nutr. 2017, 40, 795–812. [Google Scholar] [CrossRef]
- Suriyagoda, L.; Sirisena, D.; Kekulandara, D.; Bandaranayake, P.; Samarasinghe, G.; Wissuwa, M. Biomass and nutrient accumulation rates of rice cultivars differing in their growth duration when grown in fertile and low-fertile soils. J. Plant Nutr. 2020, 43, 251–269. [Google Scholar] [CrossRef]
- Julia, C.; Wissuwa, M.; Kretzschmar, T.; Jeong, K.; Rose, T. Phosphorus uptake, partitioning and redistribution during grain filling in rice. Ann. Bot. 2016, 118, 1151–1162. [Google Scholar] [CrossRef] [PubMed]
- White, P.J.; Veneklaas, E.J. Nature and nurture: The importance of seed phosphorus content. Plant Soil 2012, 357, 1–8. [Google Scholar] [CrossRef]
- Inthapanya, P.; Sihavong, P.; Sihathep, V.; Chanphengsay, M.; Fukai, S.; Basnayake, J. Genotypic performance under fertilised and non-fertilised conditions in rainfed lowland rice. Field Crops Res. 2000, 65, 1–14. [Google Scholar] [CrossRef]
- Kekulandara, D.S.; Sirisena, D.N.; Bandaranayake, P.C.G.; Samarasinghe, G.; Wissuwa, M.; Suriyagoda, L.D.B. Variation in grain yield, and nitrogen, phosphorus and potassium nutrition of irrigated rice cultivars grown at fertile and low-fertile soils. Plant Soil 2019, 434, 107–123. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genotype | Year Released/State | Description |
|---|---|---|
| Cheniere | 2003/Louisiana | A short season, semi-dwarf long-grain variety with good yield potential and milling quality comparable to Cypress. Susceptible to sheath blight and blast. |
| CL-151 | 2007/Louisiana | A mid-season, semi-dwarf long-grain Clearfield variety similar to Cocodrie with good yield potential. It is very susceptible to blast and straighthead, and susceptible to lodging and sheath blight. |
| CL-153 | 2016/Louisiana | A mid-season, semi-dwarf long-grain Clearfield variety similar to CL151 with good yield potential. Susceptible to sheath blight, kernel smut, and false smut. Moderately susceptible to blast. |
| CL-271 | 2011/Louisiana | Clearfield, medium grain variety with excellent grain quality. It is moderately resistant to blast and susceptible to sheath blight. |
| CL-272 | 2016/Louisiana | A mid-season, medium-grain Clearfield variety. High tolerance to Newpath herbicide. Very susceptible to bacterial panicle blight. Susceptible to sheath blight and blast. |
| Diamond | 2016/Arkansas | A mid-season, long-grain variety with excellent yield potential and good milling quality. Very good straw strength. Susceptible to blast and sheath blight, moderately susceptible to bacterial panicle blight. Very susceptible to false smut. |
| Dixiebelle | 1996/Texas | Short-season long-grain with ‘Newrex’ quality; specialty rice used for canning and steam tables. |
| Jupiter | 2006/Louisiana | A mid-season, semi-dwarf, medium-grain variety with excellent yield potential and milling quality. It has a small grain size but has moderate resistance to bacterial panicle blight. |
| LaKast | 2014/Arkansas | A mid-season, long-grain variety with excellent yield potential and good milling quality. Susceptible to blast and sheath blight. |
| Rex | 2010/Mississippi | A short season, semi-dwarf long-grain variety with excellent yield potential and good milling quality. Very good straw strength but is susceptible to most diseases. |
| Sierra | 2005/Texas | An aromatic long-grain with the fragrance and cooking qualities of a basmati style rice. |
| Titan | 2016/Arkansas | A short season, medium-grain variety with excellent yield potential. Moderately susceptible to blast and bacterial panicle blight. It has a preferred large grain size |
| XL753 | 2011/RiceTec, Inc | A short season, long-grain hybrid with excellent yield potential. Resistant to blast, moderately susceptible to sheath blight and straighthead. |
| XL760 | 2014/RiceTec, Inc. | A short season, long-grain hybrid with good yield potential. |
| Month | Air Temperature | Humidity (%) | Rainfall (mm) | Solar Radiation (MJm−2) | |
|---|---|---|---|---|---|
| Maximum (°C) | Minimum (°C) | ||||
| March | 26.0 ± 3.0 | 13.2 ± 2.8 | 76.8 ± 5.6 | 17.5 | 18.8 ± 3.9 |
| April | 28.1 ± 2.4 | 16.5 ± 2.8 | 78.6 ± 8.8 | 35.7 | 22.0 ± 7.3 |
| May | 30.8 ± 1.9 | 19.2 ± 2.5 | 81.6 ± 6.2 | 134.6 | 23.9 ± 4.9 |
| June | 30.5 ± 2.2 | 22.5 ± 1.0 | 90.8 ± 4.4 | 326.9 | 17.4 ± 6.2 |
| July | 32.0 ± 1.5 | 22.9 ± 0.6 | 89.3 ± 3.6 | 167.5 | 18.4 ± 5.2 |
| August | 32.3 ± 2.0 | 22.7 ± 0.8 | 88.1 ± 4.6 | 213.9 | 20.3 ± 5.6 |
| September | 31.5 ± 1.6 | 19.3 ± 10.2 | 88.6 ± 3.7 | 384.6 | 17.9 ± 4.8 |
| Effect | Df | pH | SM3K | SM3P | RK | RP | SK | SP | PK | PP | GK | GP | Yield |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Location | 1 | * | ns | * | * | * | * | * | * | * | * | * | * |
| Variety | 13 | ns | ns | ns | * | * | * | * | * | * | * | * | * |
| Location × variety | 13 | ns | ns | ns | * | * | * | * | * | * | * | * | ns |
| Parameter | Site I | Site II |
|---|---|---|
| Soil pH | ns | ns |
| SM3K | ns | ns |
| SM3P | ns | ns |
| RK | ns | * |
| RP | * | ns |
| SK | * | * |
| SP | * | ns |
| PK | ns | * |
| PP | ns | * |
| GK | ns | * |
| GP | ns | * |
| Yield | ns | * |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Amgain, N.R.; Fan, Y.; VanWeelden, M.T.; Rabbany, A.; Bhadha, J.H. From Ground to Grain: Tracing Phosphorus and Potassium in Flooded Rice Cultivar Grown on Histosols. Agriculture 2022, 12, 1250. https://doi.org/10.3390/agriculture12081250
Amgain NR, Fan Y, VanWeelden MT, Rabbany A, Bhadha JH. From Ground to Grain: Tracing Phosphorus and Potassium in Flooded Rice Cultivar Grown on Histosols. Agriculture. 2022; 12(8):1250. https://doi.org/10.3390/agriculture12081250
Chicago/Turabian StyleAmgain, Naba R., Yuchuan Fan, Matthew T. VanWeelden, Abul Rabbany, and Jehangir H. Bhadha. 2022. "From Ground to Grain: Tracing Phosphorus and Potassium in Flooded Rice Cultivar Grown on Histosols" Agriculture 12, no. 8: 1250. https://doi.org/10.3390/agriculture12081250
APA StyleAmgain, N. R., Fan, Y., VanWeelden, M. T., Rabbany, A., & Bhadha, J. H. (2022). From Ground to Grain: Tracing Phosphorus and Potassium in Flooded Rice Cultivar Grown on Histosols. Agriculture, 12(8), 1250. https://doi.org/10.3390/agriculture12081250