

Screening and Identification of the Rhizosphere Fungal Communities Associated with Land Reclamation in Egypt


 , ,
, ,  , and
, and 
Abstract
:1. Introduction
2. Materials and Methods
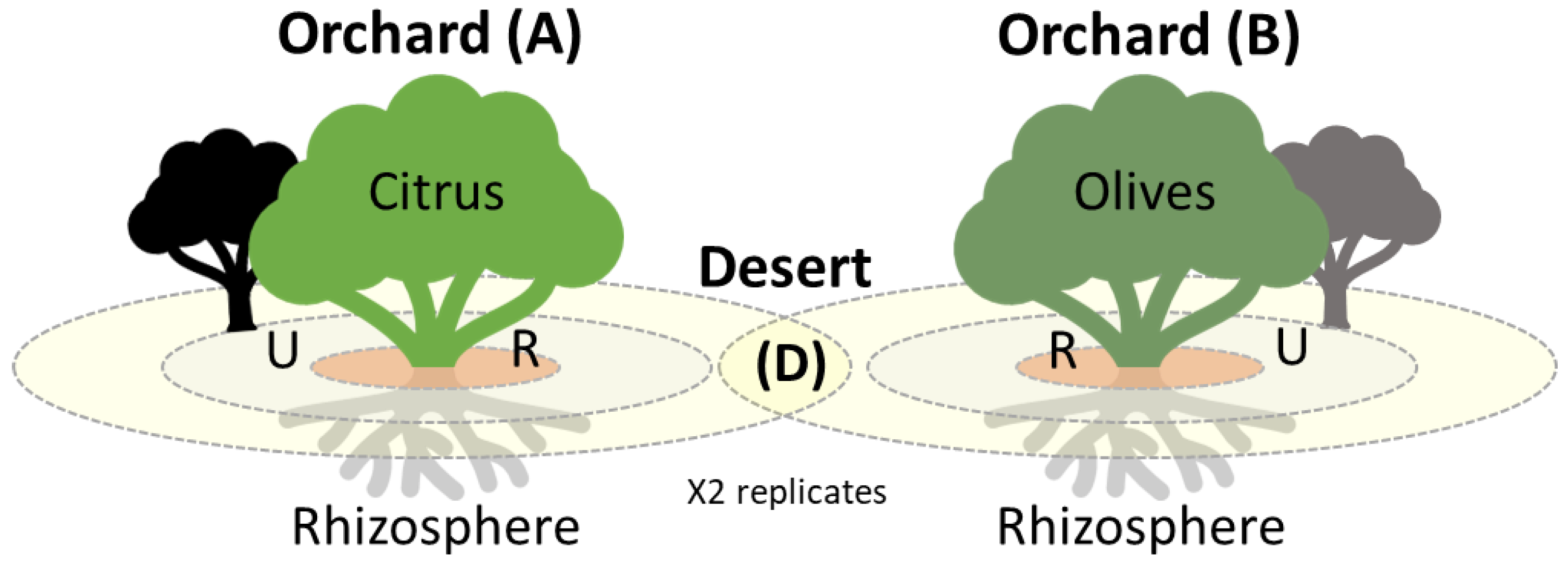
2.1. Sampling Locations
2.2. Soil Physio-Chemical Characterization
2.3. Fungal Isolation
2.4. DNA Extraction, PCR Amplification, and Sequencing
2.5. Sequence Alignment and Data Analysis
2.6. Phosphate-Solubilizing Test
2.7. Indole Acetic Acid (IAA) Production
3. Results
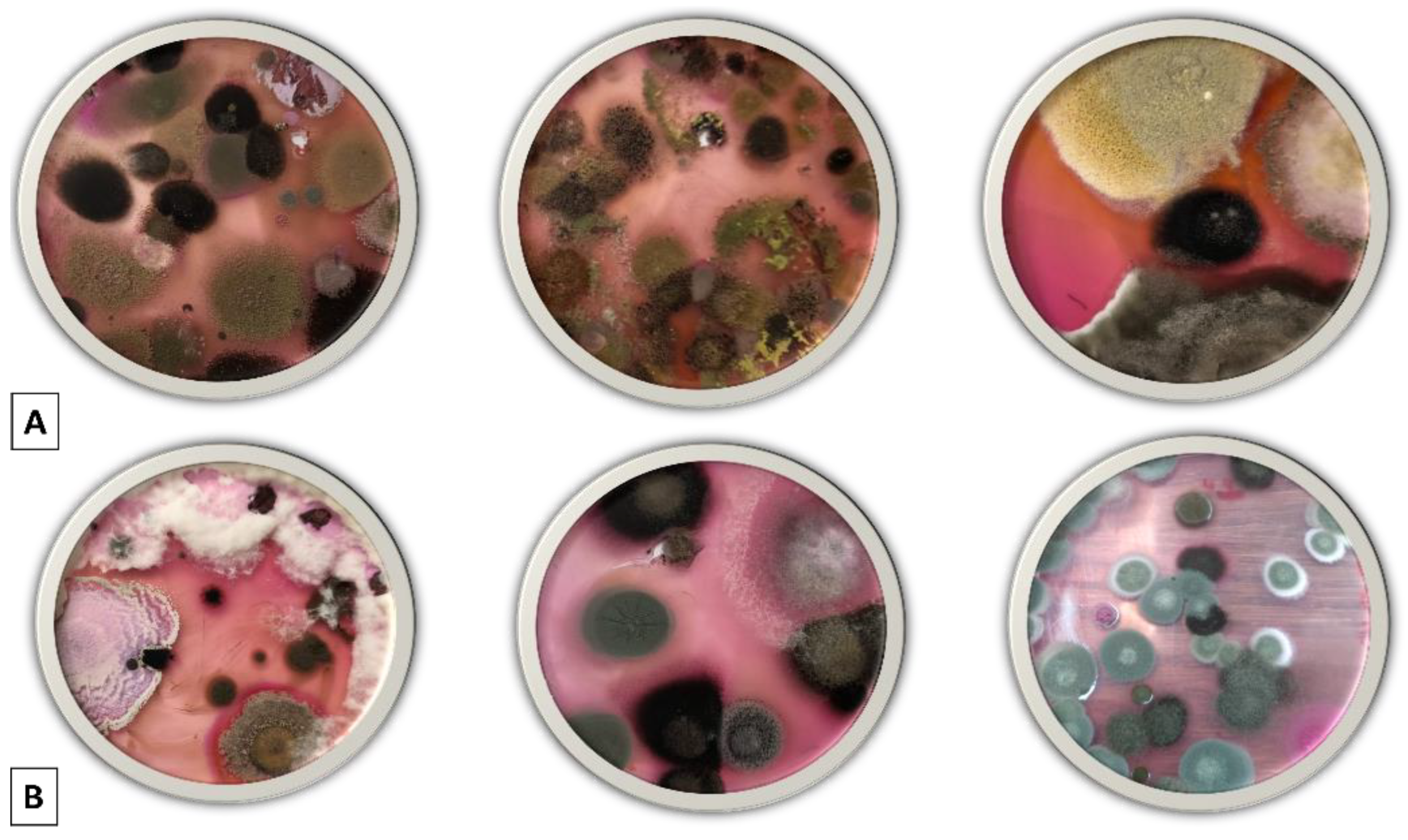
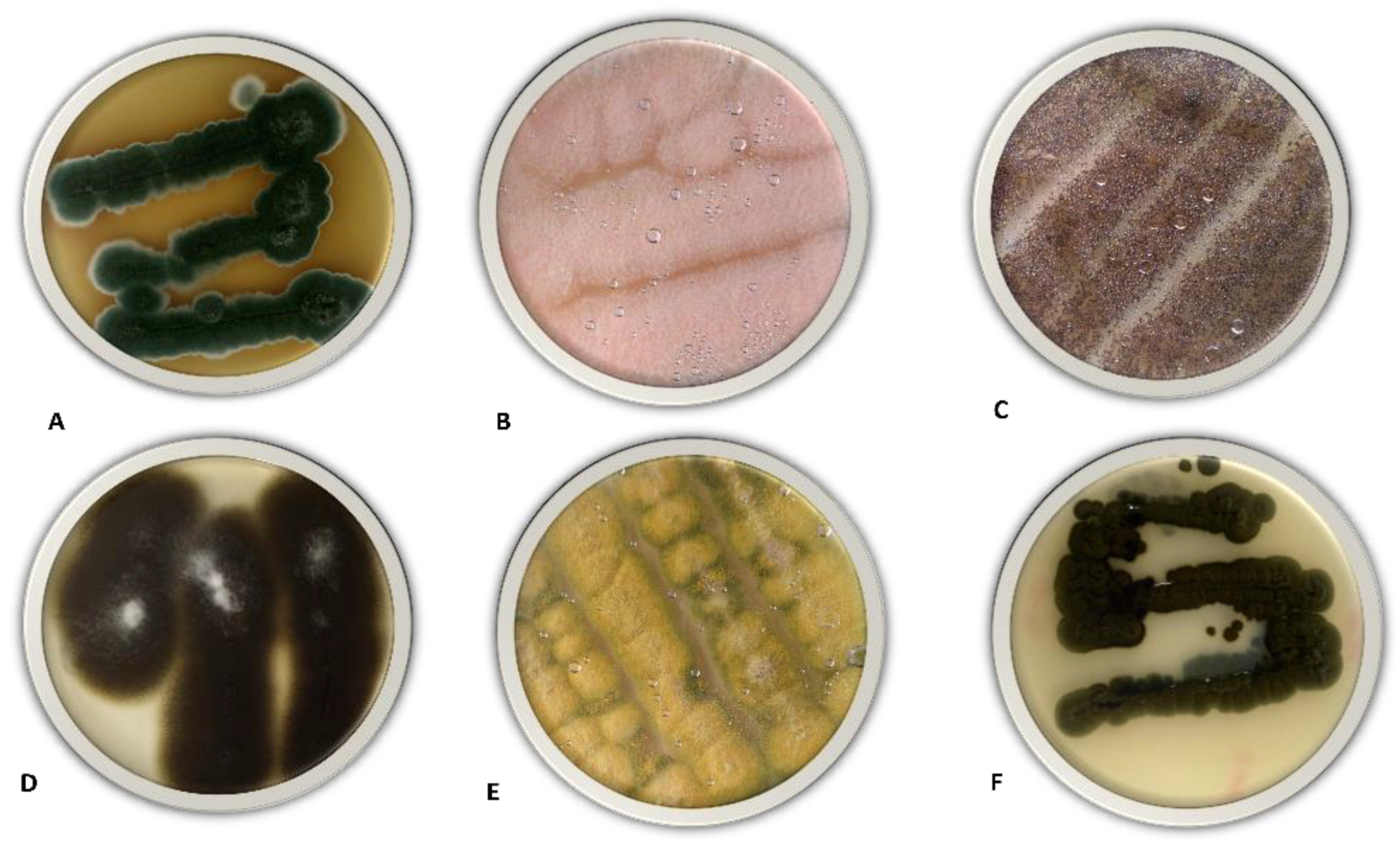
3.1. Fungal Screening and Isolation
3.2. Species Abundance
3.3. Soil Physio-Chemical Characterization
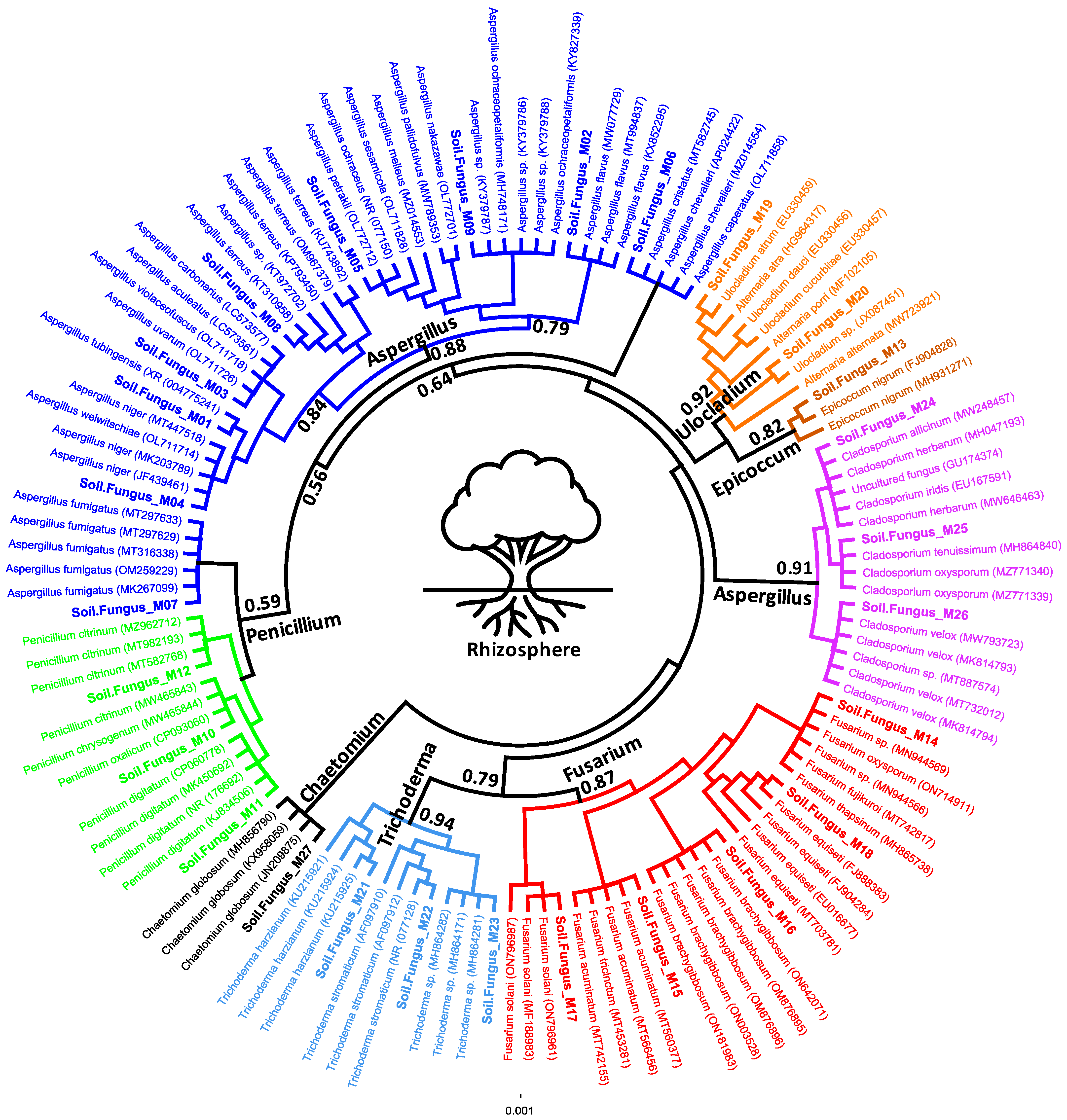
3.4. Molecular Characterization and Phylogeny
3.5. Phosphate-Solubilizing Test
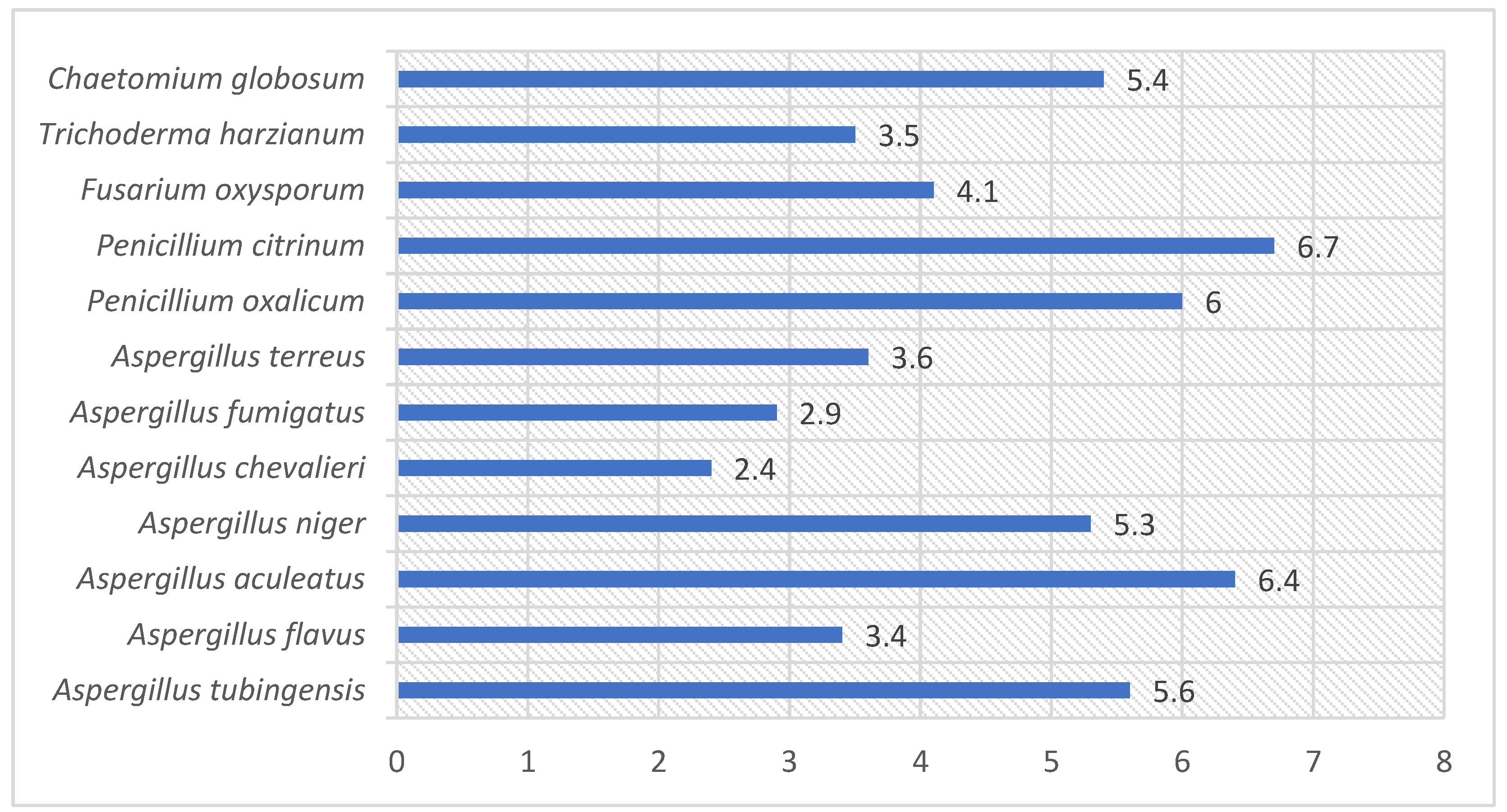
3.6. Indole Acetic Acid (IAA) Production
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Jeger, M.; Beresford, R.; Bock, C.; Brown, N.; Fox, A.; Newton, A.; Vicent, A.; Xu, X.; Yuen, J. Global Challenges Facing Plant Pathology: Multidisciplinary Approaches to Meet the Food Security and Environmental Challenges in the Mid-Twenty-First Century. CABI Agric. Biosci. 2021, 2, 20. [Google Scholar] [CrossRef]
- Nadarajah, K.; Abdul Rahman, N.S.N. Plant–Microbe Interaction: Aboveground to Belowground, from the Good to the Bad. IJMS 2021, 22, 10388. [Google Scholar] [CrossRef] [PubMed]
- Rashid, M.I.; Mujawar, L.H.; Shahzad, T.; Almeelbi, T.; Ismail, I.M.I.; Oves, M. Bacteria and Fungi Can Contribute to Nutrients Bioavailability and Aggregate Formation in Degraded Soils. Microbiol. Res. 2016, 183, 26–41. [Google Scholar] [CrossRef]
- Larney, F.J.; Angers, D.A. The Role of Organic Amendments in Soil Reclamation: A Review. Can. J. Soil. Sci. 2012, 92, 19–38. [Google Scholar] [CrossRef]
- Yang, H.-X.; Guo, S.-X.; Liu, R.-J. Characteristics of Arbuscular Mycorrhizal Fungal Diversity and Functions in Saline-Alkali Land. Yingyong Shengtai Xuebao 2015, 26, 311–320. [Google Scholar]
- Vannette, R.L.; Rasmann, S. Arbuscular Mycorrhizal Fungi Mediate Below-Ground Plant-Herbivore Interactions: A Phylogenetic Study. Funct. Ecol. 2012, 26, 1033–1042. [Google Scholar] [CrossRef]
- Hou, M.P.; Oluranti, B.O. Evaluation of Plant Growth Promoting Potential of Four Rhizobacterial Species for Indigenous System. J. Cent. South Univ. 2013, 20, 164–171. [Google Scholar] [CrossRef]
- Igiehon, N.O.; Babalola, O.O. Biofertilizers and Sustainable Agriculture: Exploring Arbuscular Mycorrhizal Fungi. Appl. Microbiol. Biotechnol. 2017, 101, 4871–4881. [Google Scholar] [CrossRef]
- Dias, A.C.F.; Hoogwout, E.F.; Pereira e Silva, M.d.C.; Salles, J.F.; van Overbeek, L.S.; van Elsas, J.D. Potato Cultivar Type Affects the Structure of Ammonia Oxidizer Communities in Field Soil under Potato beyond the Rhizosphere. Soil Biol. Biochem. 2012, 50, 85–95. [Google Scholar] [CrossRef]
- Philippot, L.; Raaijmakers, J.M.; Lemanceau, P.; van der Putten, W.H. Going Back to the Roots: The Microbial Ecology of the Rhizosphere. Nat. Rev. Microbiol. 2013, 11, 789–799. [Google Scholar] [CrossRef]
- Chen, Y.-L.; Xu, T.-L.; Veresoglou, S.D.; Hu, H.-W.; Hao, Z.-P.; Hu, Y.-J.; Liu, L.; Deng, Y.; Rillig, M.C.; Chen, B.-D. Plant Diversity Represents the Prevalent Determinant of Soil Fungal Community Structure across Temperate Grasslands in Northern China. Soil Biol. Biochem. 2017, 110, 12–21. [Google Scholar] [CrossRef]
- Lenc, L.; Kwaśna, H.; Sadowski, C.; Grabowski, A. Microbiota in Wheat Roots, Rhizosphere and Soil in Crops Grown in Organic and Other Production Systems. J. Phytopathol. 2015, 163, 245–263. [Google Scholar] [CrossRef]
- Alexopoulos, C.J.; Mims, C.W.; Blackwell, M. Introductory Mycology; John Wiley and Sons: Hoboken, NJ, USA, 1996; ISBN 0-471-52229-5. [Google Scholar]
- Ainsworth, G.C. Ainsworth & Bisby’s Dictionary of the Fungi; CABI: Wallingford, UK, 2008; ISBN 0-85199-826-7. [Google Scholar]
- Cookson, W.R.; Murphy, D.V.; Roper, M.M. Characterizing the Relationships between Soil Organic Matter Components and Microbial Function and Composition along a Tillage Disturbance Gradient. Soil Biol. Biochem. 2008, 40, 763–777. [Google Scholar] [CrossRef]
- Huang, Y.; Long, X.-E. Contribution of Fungi to Soil Nitrous Oxide Emission and Their Research Methods: A Review. Yingyong Shengtai Xuebao 2014, 25, 1213–1220. [Google Scholar] [PubMed]
- Haas, H. Fungal Siderophore Metabolism with a Focus on Aspergillus fumigatus. Nat. Prod. Rep. 2014, 31, 1266–1276. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Z.; Bai, T.; Dai, L.; Wang, F.; Tao, J.; Meng, S.; Hu, Y.; Wang, S.; Hu, S. A Study of Organic Acid Production in Contrasts between Two Phosphate Solubilizing Fungi: Penicillium oxalicum and Aspergillus niger. Sci. Rep. 2016, 6, 25313. [Google Scholar] [CrossRef]
- Narsian, V.; Patel, H.H. Aspergillus Aculeatus as a Rock Phosphate Solubilizer. Soil Biol. Biochem. 2000, 32, 559–565. [Google Scholar] [CrossRef]
- Kumar, N.V.; Rajam, K.S.; Rani, M.E. Plant Growth Promotion Efficacy of Indole Acetic Acid (IAA) Produced by a Mangrove Associated Fungi-Trichoderma Viride VKF3. Int. J. Curr. Microbiol. Appl. Sci. 2017, 6, 2692–2701. [Google Scholar] [CrossRef]
- Korkar, M.H.; Magdy, M.; Rizk, S.M.; Fiteha, Y.G.; Atta, A.H.; Rashed, M.A.-S. Rhizosphere-Associated Microbiome Profile of Agriculture Reclaimed Lands in Egypt. Life Sci. 2022. preprint. [Google Scholar]
- Wang, Q.; Ren, Y.; Meng, L.; Li, H.; Fu, H.; Wang, H. Simultaneous Determination of Total Nitrogen and Organic Carbon in Soil with an Elemental Analyzer. Chin. J. Anal. Lab. 2013, 32, 41–45. [Google Scholar]
- Xu, W.; Yuan, W. Responses of Microbial Biomass Carbon and Nitrogen to Experimental Warming: A Meta-Analysis. Soil Biol. Biochem. 2017, 115, 265–274. [Google Scholar] [CrossRef]
- Muhammad, N.; Dai, Z.; Xiao, K.; Meng, J.; Brookes, P.C.; Liu, X.; Wang, H.; Wu, J.; Xu, J. Changes in Microbial Community Structure Due to Biochars Generated from Different Feedstocks and Their Relationships with Soil Chemical Properties. Geoderma 2014, 226–227, 270–278. [Google Scholar] [CrossRef]
- Jones, D.; Willett, V. Experimental Evaluation of Methods to Quantify Dissolved Organic Nitrogen (DON) and Dissolved Organic Carbon (DOC) in Soil. Soil Biol. Biochem. 2006, 38, 991–999. [Google Scholar] [CrossRef]
- Reddy, P.L.N.; Babu, B.S.; Radhaiah, A.; Sreeramulu, A. Screening, Identification and Isolation of Cellulolytic Fungi from Soils of Chittoor District, India. Int. J. Curr. Microbiol. Appl. Sci. 2014, 3, 761–771. [Google Scholar]
- Gaddeyya, G.; Niharika, P.S.; Bharathi, P.; Kumar, P.R. Isolation and Identification of Soil Mycoflora in Different Crop Fields at Salur Mandal. Adv. Appl. Sci. Res. 2012, 3, 2020–2026. [Google Scholar]
- Li, X.; Li, D.; Yan, J.; Zhang, Y.; Wang, H.; Zhang, J.; Ahmed, T.; Li, B. Effect of Plant-Growth-Promoting Fungi on Eggplant (Solanum melongena L.) in New Reclamation Land. Agriculture 2021, 11, 1036. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and Direct Sequencing of Fungal Ribosomal RNA Genes for Phylogenetics. In PCR Protocols; Elsevier: Amsterdam, The Netherlands, 1990; pp. 315–322. ISBN 978-0-12-372180-8. [Google Scholar]
- Kearse, M.; Moir, R.; Wilson, A.; Stones-Havas, S.; Cheung, M.; Sturrock, S.; Buxton, S.; Cooper, A.; Markowitz, S.; Duran, C.; et al. Geneious Basic: An Integrated and Extendable Desktop Software Platform for the Organization and Analysis of Sequence Data. Bioinformatics 2012, 28, 1647–1649. [Google Scholar] [CrossRef] [Green Version]
- Katoh, K.; Standley, D.M. MAFFT Multiple Sequence Alignment Software Version 7: Improvements in Performance and Usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [Green Version]
- Price, M.N.; Dehal, P.S.; Arkin, A.P. FastTree 2—Approximately Maximum-Likelihood Trees for Large Alignments. PLoS ONE 2010, 5, e9490. [Google Scholar] [CrossRef]
- Ronquist, F.; Teslenko, M.; van der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient Bayesian Phylogenetic Inference and Model Choice Across a Large Model Space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef] [Green Version]
- Wakelin, S.A.; Gupta, V.V.S.R.; Harvey, P.R.; Ryder, M.H. The Effect of Penicillium Fungi on Plant Growth and Phosphorus Mobilization in Neutral to Alkaline Soils from Southern Australia. Can. J. Microbiol. 2007, 53, 106–115. [Google Scholar] [CrossRef] [PubMed]
- Bano, N.; Musarrat, J. Characterization of a New Pseudomonas Aeruginosa Strain NJ-15 as a Potential Biocontrol Agent. Curr. Microbiol. 2003, 46, 324–328. [Google Scholar] [CrossRef] [PubMed]
- Khamna, S.; Yokota, A.; Lumyong, S. Actinomycetes Isolated from Medicinal Plant Rhizosphere Soils: Diversity and Screening of Antifungal Compounds, Indole-3-Acetic Acid and Siderophore Production. World J. Microbiol. Biotechnol. 2009, 25, 649–655. [Google Scholar] [CrossRef]
- Kacaniova, M.; Fikselová, M. Mycological Flora on Tree Fruits, Crust, Leaves and Pollen Sorbus domestica L. Ann. Agric. Environ. Med. 2007, 14, 229–232. [Google Scholar] [PubMed]
- Anderson, I.C.; Parkin, P.I. Detection of Active Soil Fungi by RT-PCR Amplification of Precursor RRNA Molecules. J. Microbiol. Methods 2007, 68, 248–253. [Google Scholar] [CrossRef] [PubMed]
- Houbraken, J.; Kocsubé, S.; Visagie, C.M.; Yilmaz, N.; Wang, X.-C.; Meijer, M.; Kraak, B.; Hubka, V.; Bensch, K.; Samson, R.A.; et al. Classification of Aspergillus, Penicillium, Talaromyces and Related Genera (Eurotiales): An Overview of Families, Genera, Subgenera, Sections, Series and Species. Stud. Mycol. 2020, 95, 5–169. [Google Scholar] [CrossRef]
- Plassart, P.; Akpa Vinceslas, M.; Gangneux, C.; Mercier, A.; Barray, S.; Laval, K. Molecular and Functional Responses of Soil Microbial Communities under Grassland Restoration. Agric. Ecosyst. Environ. 2008, 127, 286–293. [Google Scholar] [CrossRef]
- Harman, G.E.; Lorito, M.; Lynch, J.M. Uses of Trichoderma Spp. to Alleviate or Remediate Soil and Water Pollution. In Advances in Applied Microbiology; Elsevier: Amsterdam, The Netherlands, 2004; Volume 56, pp. 313–330. ISBN 978-0-12-002658-6. [Google Scholar]
- Contreras-Cornejo, H.A.; Macías-Rodríguez, L.; Cortés-Penagos, C.; López-Bucio, J. Trichoderma Virens, a Plant Beneficial Fungus, Enhances Biomass Production and Promotes Lateral Root Growth through an Auxin-Dependent Mechanism in Arabidopsis. Plant Physiol. 2009, 149, 1579–1592. [Google Scholar] [CrossRef] [Green Version]
- Yadav, L.S.; Kushwaha, V.; Jain, A. Isolation and Screening of Phosphate Solubilizing Fungi from Okra Rhizosphere Soil and Their Effect on the Growth of Okra Plant (Abelmoschous esculentus L.). Trop. Plant Res. 2020, 7, 277–284. [Google Scholar] [CrossRef]
- Udoh, I.P.; Eleazar, C.I.; Ogeneh, B.O.; Ohanu, M.E. Studies on Fungi Responsible for the Spoilage/Deterioration of Some Edible Fruits and Vegetables. AiM 2015, 05, 285–290. [Google Scholar] [CrossRef] [Green Version]
- Alwakeel, S.S. Molecular Identification of Isolated Fungi from Stored Apples in Riyadh, Saudi Arabia. Saudi J. Biol. Sci. 2013, 20, 311–317. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, F.; Cai, T.; Yang, X.; Sui, W. Soil Fungal Community Variation by Large-Scale Reclamation in Sanjiang Plain, China. Ann. Microbiol. 2017, 67, 679–689. [Google Scholar] [CrossRef]
- Li, H.; Liu, W.; Wan, F.; Cao, Y. Invasive Impacts of Ageratina adenophora (Asteraceae) on the Changes of Microbial Community Structure, Enzyme Activity and Fertility in Soil Ecosystem. Sci. Agric. Sin. 2009, 42, 3964–3971. [Google Scholar]
- Birkhofer, K.; Schöning, I.; Alt, F.; Herold, N.; Klarner, B.; Maraun, M.; Marhan, S.; Oelmann, Y.; Wubet, T.; Yurkov, A.; et al. General Relationships between Abiotic Soil Properties and Soil Biota across Spatial Scales and Different Land-Use Types. PLoS ONE 2012, 7, e43292. [Google Scholar] [CrossRef]
- Ye, X.; Huang, Z.-G.; Wu, H.-T. Carbon and Nitrogen Distributions and Microbial Characteristics in the Soils of Four Types of Wetlands in Sanjiang Plain, Northeast China. Yingyong Shengtai Xuebao 2014, 25, 2847–2854. [Google Scholar]
- Gyaneshwar, P.; Naresh Kumar, G.; Parekh, L.J.; Poole, P.S. Role of soil microorganisms in improving P nutrition of plants. Plant Soil 2002, 245, 83–93. [Google Scholar] [CrossRef]
- Zapata, F.; Zaharah, A.R. Phosphorus Availability from Phosphate Rock and Sewage Sludge as Influenced by the Addition of Water Soluble Phosphate Fertilizer. Nutr. Cycl. Agroecosyst. 2002, 63, 43–48. [Google Scholar] [CrossRef]
- Ali, S.; Khan, S.A.; Hamayun, M.; Iqbal, A.; Khan, A.L.; Hussain, A.; Shah, M. Endophytic Fungi from Caralluma acutangula Can Secrete Plant Growth Promoting Enzymes. Fresenius Environ. Bull. 2019, 28, 2688–2696. [Google Scholar]
- Khan, A.L.; Al-Harrasi, A.; Al-Rawahi, A.; Al-Farsi, Z.; Al-Mamari, A.; Waqas, M.; Asaf, S.; Elyassi, A.; Mabood, F.; Shin, J.-H.; et al. Endophytic Fungi from Frankincense Tree Improves Host Growth and Produces Extracellular Enzymes and Indole Acetic Acid. PLoS ONE 2016, 11, e0158207. [Google Scholar] [CrossRef] [Green Version]
- Nath, R.; Sharma, G.; Barooah, M. Plant Growth Promoting Endophytic Fungi Isolated from Tea (Camellia sinensis) Shrubs of Assam, India. Appl. Ecol. Environ. Res. 2015, 13, 877–891. [Google Scholar]
- Ripa, F.A.; Cao, W.; Tong, S.; Sun, J. Assessment of Plant Growth Promoting and Abiotic Stress Tolerance Properties of Wheat Endophytic Fungi. BioMed Res. Int. 2019, 2019, 6105865. [Google Scholar] [CrossRef] [PubMed]
- Mehmood, A.; Irshad, M.; Husna, A.A.; Hussain, A. In Vitro Maize Growth Promotion by Endophytic Fusarium Oxysporum WLW. J. Appl. Environ. Biol. Sci. 2018, 8, 30–35. [Google Scholar]
- Mehmood, A.; Khan, N.; Irshad, M.; Hamayun, M.; Husna, I.; Javed, A.; Hussain, A. IAA Producing Endopytic Fungus Fusariun Oxysporum Wlw Colonize Maize Roots and Promoted Maize Growth Under Hydroponic Condition. Eur. J. Exp. Biol. 2018, 8, 24. [Google Scholar] [CrossRef]
- Yoo, S.-J.; Shin, D.J.; Won, H.Y.; Song, J.; Sang, M.K. Aspergillus Terreus JF27 Promotes the Growth of Tomato Plants and Induces Resistance against Pseudomonas Syringae Pv. Tomato. Mycobiology 2018, 46, 147–153. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wardeh, M.; Risley, C.; McIntyre, M.K.; Setzkorn, C.; Baylis, M. Database of Host-Pathogen and Related Species Interactions, and Their Global Distribution. Sci. Data 2015, 2, 150049. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Reclaimed Lands | Plantation | Sample Area | Code | Composite Samples | Replicates |
|---|---|---|---|---|---|
| Citrus Orchard (A) | Citrus trees 1 | Rhizosphere (R) | AR1 | 5 | 2x |
| Uncultivated (U) | AU1 | 5 | 2x | ||
| Citrus trees 2 | Rhizosphere (R) | AR2 | 5 | 2x | |
| Uncultivated (U) | AU2 | 5 | 2x | ||
| Olive Orchard (B) | Olives trees 1 | Rhizosphere (R) | BR1 | 5 | 2x |
| Uncultivated (U) | BU1 | 5 | 2x | ||
| Olives trees 2 | Rhizosphere (R) | BR2 | 5 | 2x | |
| Uncultivated (U) | BU2 | 5 | 2x | ||
| Desert Lands (D) | No plantations | Rhizosphere (R) | DU1 | 5 | 2x |
| DU2 | 5 | 2x | |||
| DU3 | 5 | 2x |
| Soil Samples Code | Vegetation | Sample Area | Total CFU/g |
|---|---|---|---|
| AR1 | Citrus trees | Rhizosphere | 3.8 × 103 |
| AU1 | Uncultivated | 1.6 × 103 | |
| AR2 | Citrus trees | Rhizosphere | 4.4 × 103 |
| AU2 | Uncultivated | 1.8 × 103 | |
| BR1 | Olives trees | Rhizosphere | 4.8 × 103 |
| BU1 | Uncultivated | 2.7 × 103 | |
| BR2 | Olives trees | Rhizosphere | 3.0 × 103 |
| BU2 | Uncultivated | 2.2 × 103 | |
| DU1 | Desert | Uncultivated | 1.2 × 102 |
| DU2 | 1.1 × 102 | ||
| DU3 | 0.7 × 102 |
| Species Code | Species Name | Reclaimed Land | Desert Land | |||
|---|---|---|---|---|---|---|
| Olives Trees | Citrus Trees | |||||
| R% | U% | R% | U% | |||
| SFSA1 | Aspergillus tubingensis | 7.69 | 4.08 | - | - | - |
| SFSA2 | Aspergillus flavus | 8.97 | - | - | - | 6.89 |
| SFSA3 | Aspergillus aculeatus | 6.41 | 6.12 | 8.53 | - | 6.89 |
| SFSA4 | Aspergillus niger | 2.56 | 6.12 | 6.09 | 8.82 | - |
| SFSA5 | Aspergillus ochraceus | - | - | - | - | 13.79 |
| SFSA6 | Aspergillus chevalieri | - | - | - | - | 10.34 |
| SFSA7 | Aspergillus fumigatus | - | - | 8.53 | - | 6.89 |
| SFSA8 | Aspergillus terreus | 7.69 | - | - | - | - |
| SFSA9 | Aspergillus sp. | 3.84 | 4.08 | 4.87 | 23.52 | - |
| SFSA10 | Penicillium oxalicum | 10.25 | 8.16 | 7.31 | 5.88 | - |
| SFSA11 | Penicillium digitatum | - | - | 6.09 | 8.82 | 6.89 |
| SFSA12 | Penicillium citrinum | 3.84 | 10.20 | 7.31 | 11.76 | - |
| SFSA13 | Epicoccum nigrum | 5.12 | 6.12 | - | - | - |
| SFSA14 | Fusarium oxysporum | 6.41 | 14.28 | 4.87 | 8.82 | 6.89 |
| SFSA15 | Fusarium acuminatum | - | - | 6.09 | 8.82 | - |
| SFSA16 | Fusarium brachygibbosum | - | - | 4.87 | - | - |
| SFSA17 | Fusarium solani | - | - | 7.31 | 2.94 | - |
| SFSA18 | Fusarium equiseti | 2.56 | 8.16 | - | - | - |
| SFSA19 | Ulocladium atrum | 2.56 | - | 8.82 | - | |
| SFSA20 | Ulocladium sp. | - | - | - | - | 13.79 |
| SFSA21 | Trichoderma harzianum | 11.53 | 14.28 | 6.09 | - | - |
| SFSA22 | Trichoderma stromaticum | 8.97 | 6.12 | 4.87 | 5.88 | - |
| SFSA23 | Trichoderma sp. | 3.84 | 4.08 | 4.87 | - | - |
| SFSA24 | Cladosporium herbarum | - | - | - | - | 10.34 |
| SFSA25 | Cladosporium oxysporum | - | - | 7.31 | 5.88 | - |
| SFSA26 | Cladosporium velox | - | - | - | - | 6.89 |
| SFSA27 | Chaetomium globosum | 7.69 | 6.12 | 4.87 | - | 10.34 |
| Fungal Species * | IAA Concentrations (µg/mL) * | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| pH5 NS | pH7 NS | pH9 NS | ||||||||||
| 0.1% b | 0.5% b | 1.0% a | 1.5% a | 0.1% b | 0.5% b | 1.0% a | 1.5% a | 0.1% b | 0.5% b | 1.0% a | 1.5% a | |
| A. tubingensis a | 27 | 75 | 240 | 176 | 10 | 126 | 215 | 260 | 63 | 25 | 73 | 84 |
| A. flavus a | 45 | 31 | 33 | 85 | 52 | 92 | 198 | 210 | 98 | 84 | 92 | 72 |
| A. niger ab | 76 | 75 | 72 | 89 | 95 | 87 | 124 | 85 | 59 | 79 | 75 | 73 |
| A. fumigatus a | 41 | 78 | 210 | 97 | 37 | 85 | 197 | 212 | 88 | 43 | 76 | 79 |
| A. terreus c | 16 | 22 | 35 | 83 | 43 | 7 | 36 | 39 | 35 | 56 | 79 | 85 |
| P. oxalicum c | 13 | 22 | 34 | 62 | 11 | 19 | 23 | 40 | 35 | 45 | 76 | 82 |
| P. citrinum c | 7 | 21 | 32 | 79 | 6 | 26 | 45 | 66 | 35 | 67 | 77 | 80 |
| F. oxysporum c | 22 | 31 | 34 | 75 | 23 | 19 | 40 | 33 | 35 | 53 | 57 | 61 |
| T. harzianum bc | 45 | 54 | 43 | 89 | 18 | 30 | 48 | 55 | 39 | 73 | 78 | 92 |
| C. globosum c | 32 | 31 | 34 | 22 | 19 | 43 | 79 | 50 | 62 | 76 | 63 | 78 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nafaa, M.; Rizk, S.M.; Aly, T.A.-G.A.; Rashed, M.A.-S.; Abd El-Moneim, D.; Ben Bacha, A.; Alonazi, M.; Magdy, M. Screening and Identification of the Rhizosphere Fungal Communities Associated with Land Reclamation in Egypt. Agriculture 2023, 13, 215. https://doi.org/10.3390/agriculture13010215
Nafaa M, Rizk SM, Aly TA-GA, Rashed MA-S, Abd El-Moneim D, Ben Bacha A, Alonazi M, Magdy M. Screening and Identification of the Rhizosphere Fungal Communities Associated with Land Reclamation in Egypt. Agriculture. 2023; 13(1):215. https://doi.org/10.3390/agriculture13010215
Chicago/Turabian StyleNafaa, Mostafa, Samah Mohamed Rizk, Tahany Abdel-Ghafar Ahmed Aly, Mohamed Abdel-Salam Rashed, Diaa Abd El-Moneim, Abir Ben Bacha, Mona Alonazi, and Mahmoud Magdy. 2023. "Screening and Identification of the Rhizosphere Fungal Communities Associated with Land Reclamation in Egypt" Agriculture 13, no. 1: 215. https://doi.org/10.3390/agriculture13010215
APA StyleNafaa, M., Rizk, S. M., Aly, T. A.-G. A., Rashed, M. A.-S., Abd El-Moneim, D., Ben Bacha, A., Alonazi, M., & Magdy, M. (2023). Screening and Identification of the Rhizosphere Fungal Communities Associated with Land Reclamation in Egypt. Agriculture, 13(1), 215. https://doi.org/10.3390/agriculture13010215