
Spent Coffee Grounds Applied as a Top-Dressing or Incorporated into the Soil Can Improve Plant Growth While Reducing Slug Herbivory



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Methods
2.1. Survival and Growth of BSF on SCG
2.1.1. Soldier Fly Colony
2.1.2. Effects of BSF Size on Survival and Growth on SCG
2.1.3. Effects of SCG on Larval Growth and Development
2.2. Effects of SCG and BSF Frass of Plant Growth
2.2.1. Soil Amendments
2.2.2. Effects of Soil Amendments on Growth of Radish
2.2.3. Effects of Soil Amendments on Growth of Tomato
2.3. Effects of SCG on Slug Herbivory
2.3.1. Slug Species
2.3.2. Repellent Effects of SCG on Slug Herbivory
2.3.3. Effects of Plant Quality of Slug Herbivory
2.4. Statistical Analyses
3. Results
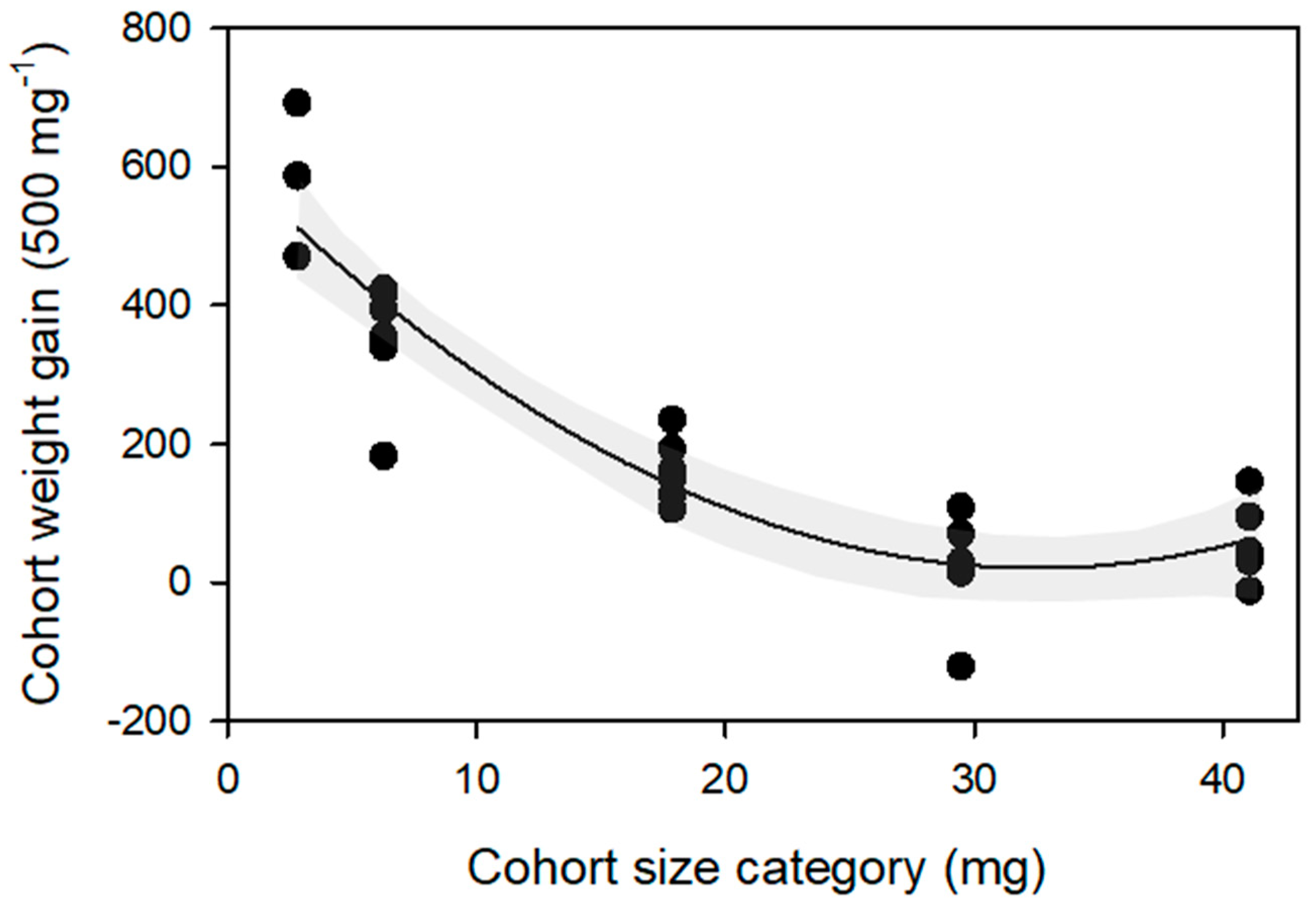
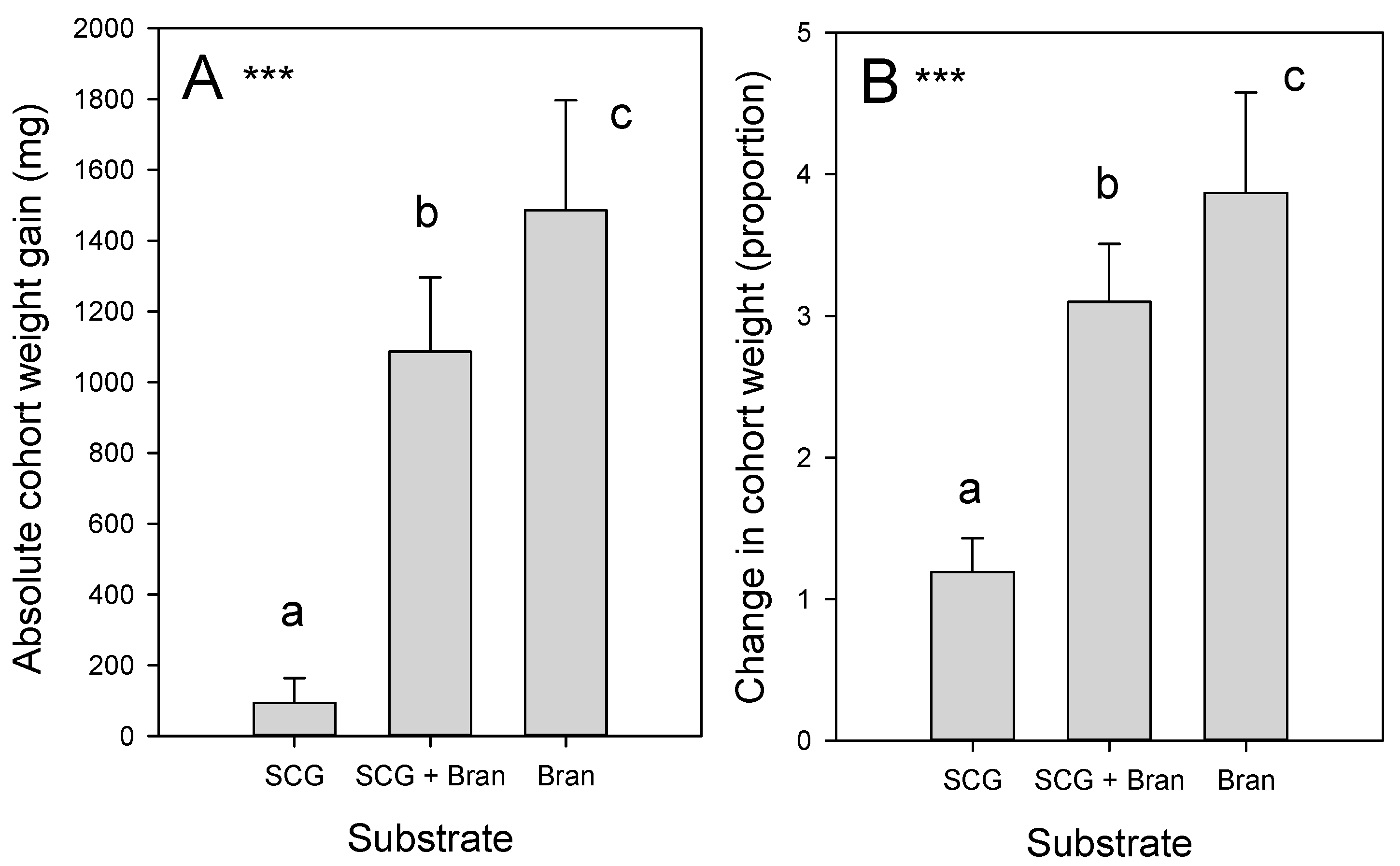
3.1. BSF Larval Survival and Growth on SCG
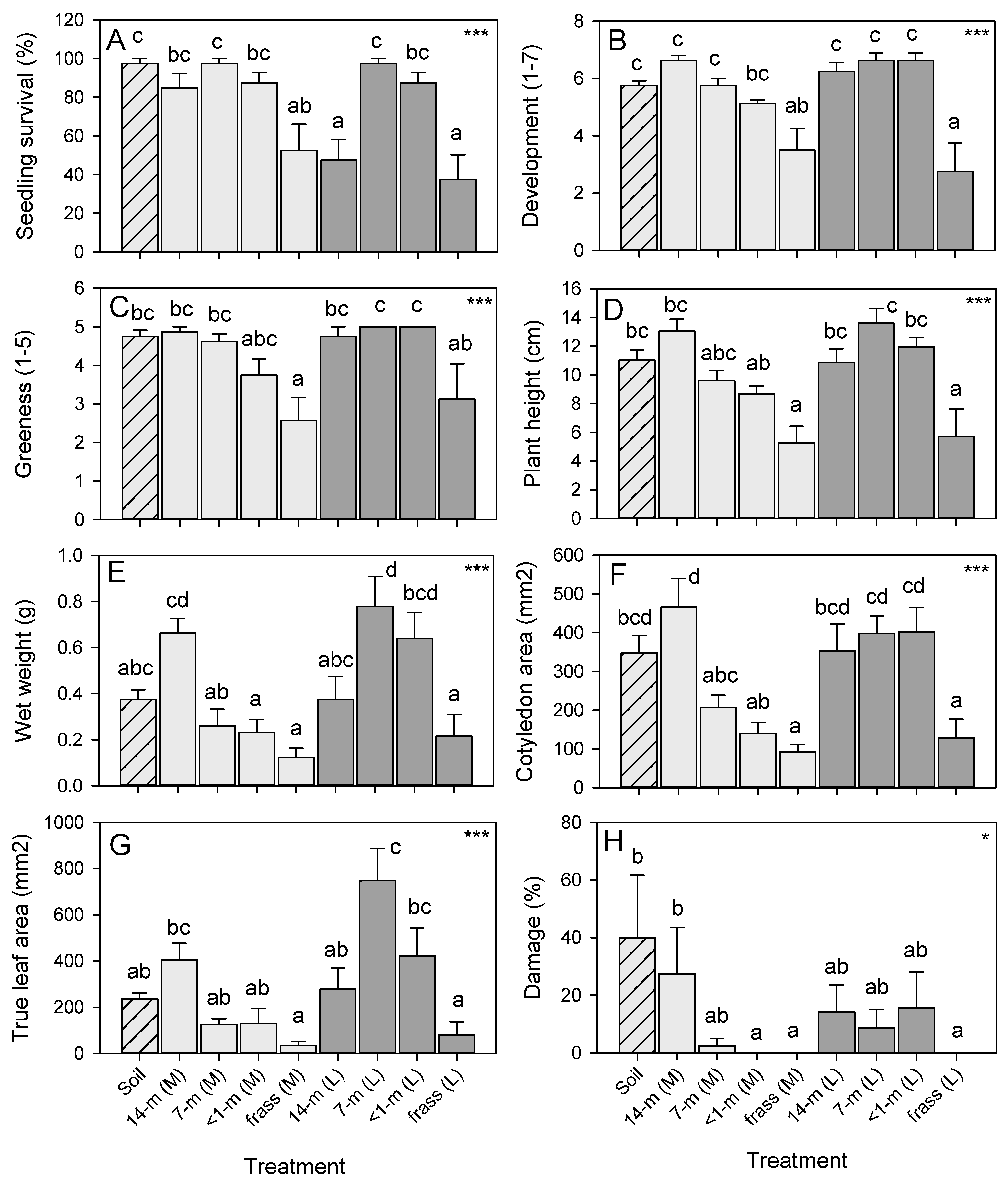
3.2. Effects of Amendments on Radish Growth and Development
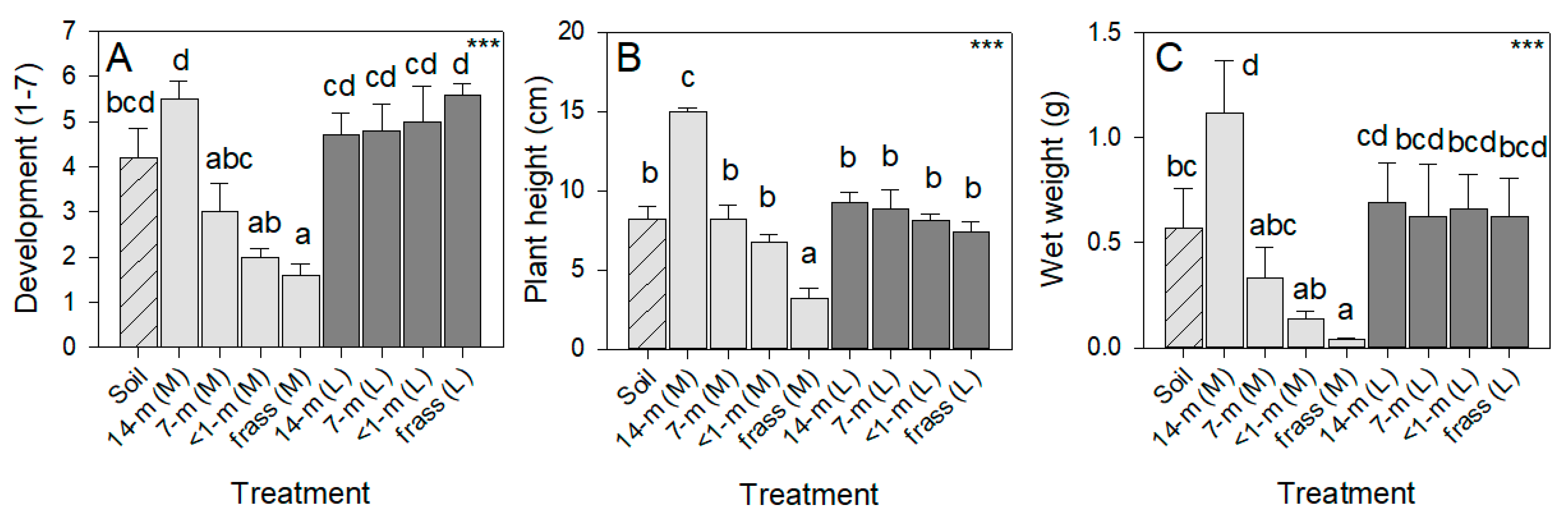
3.3. Effects of Amendments on Tomato Growth and Development
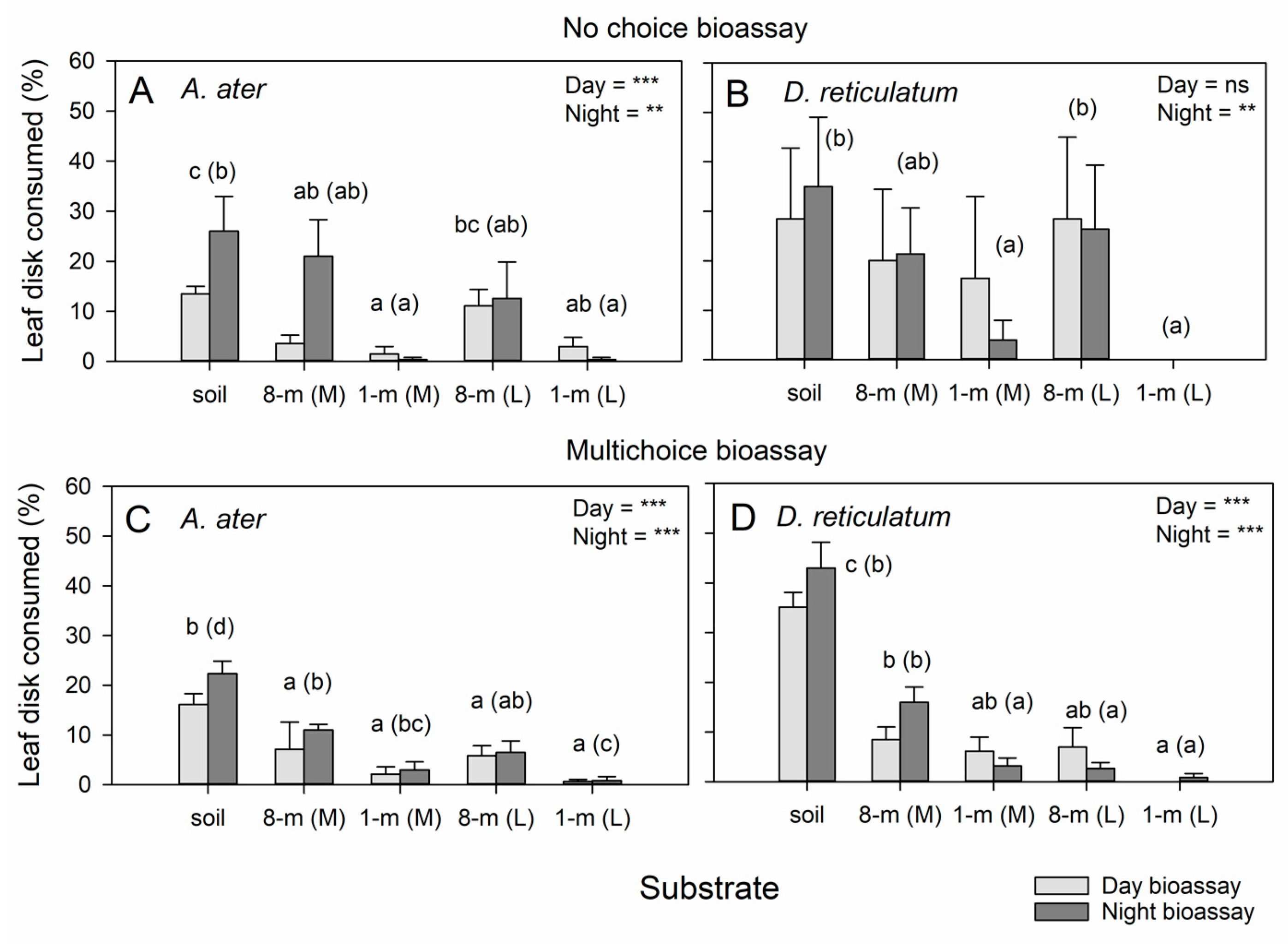
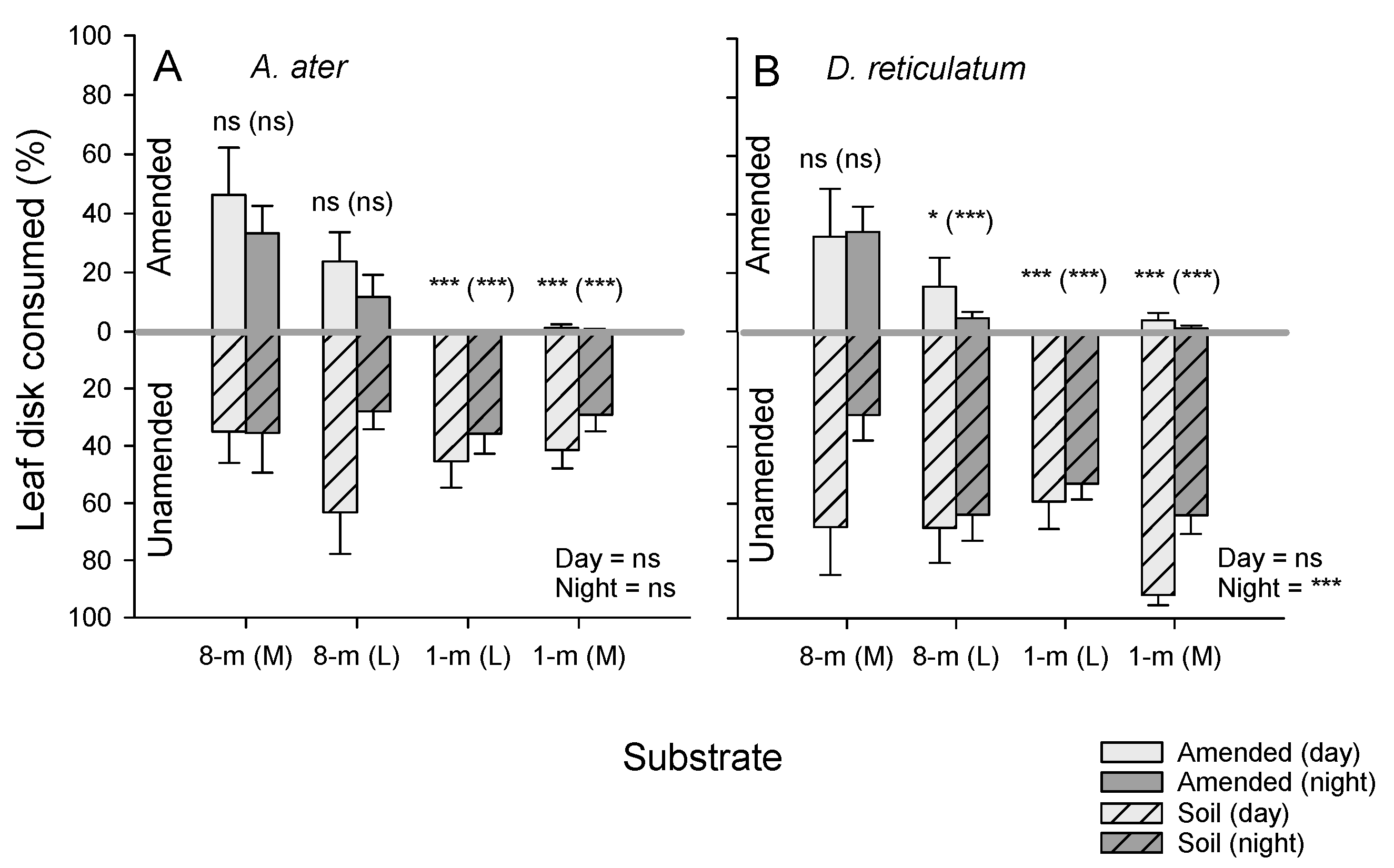
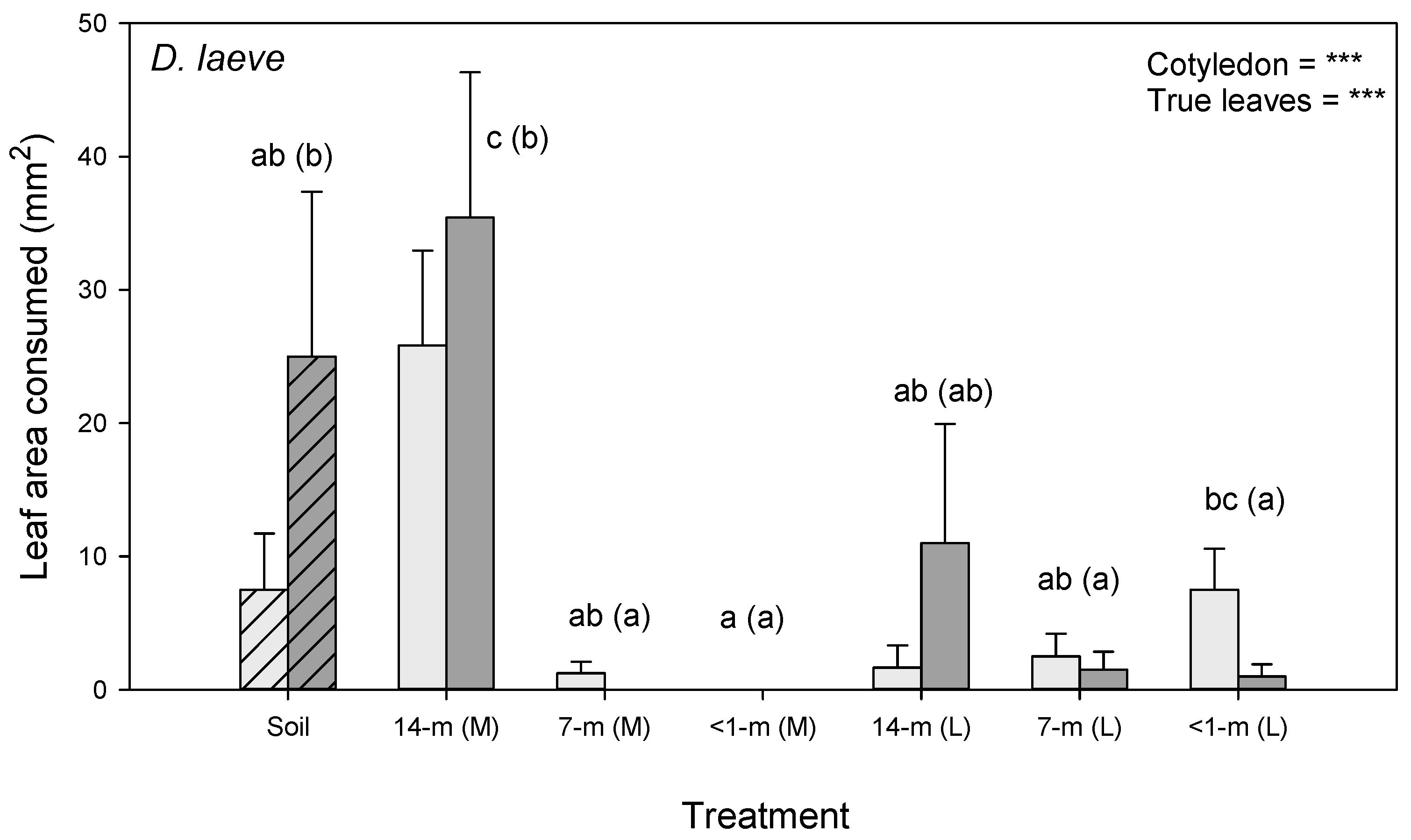
3.4. Repellent Effects of SCG on Slug Herbivory
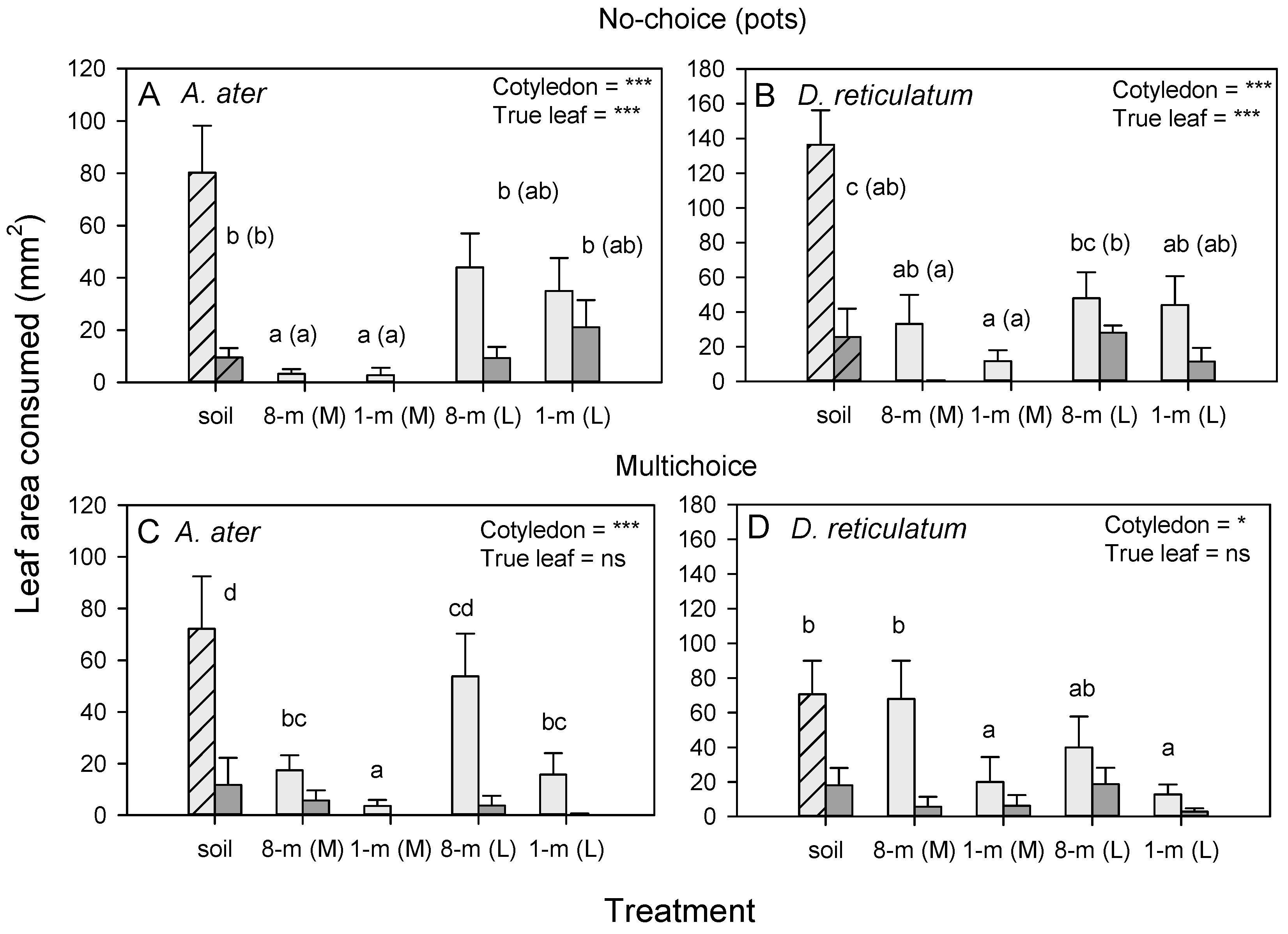
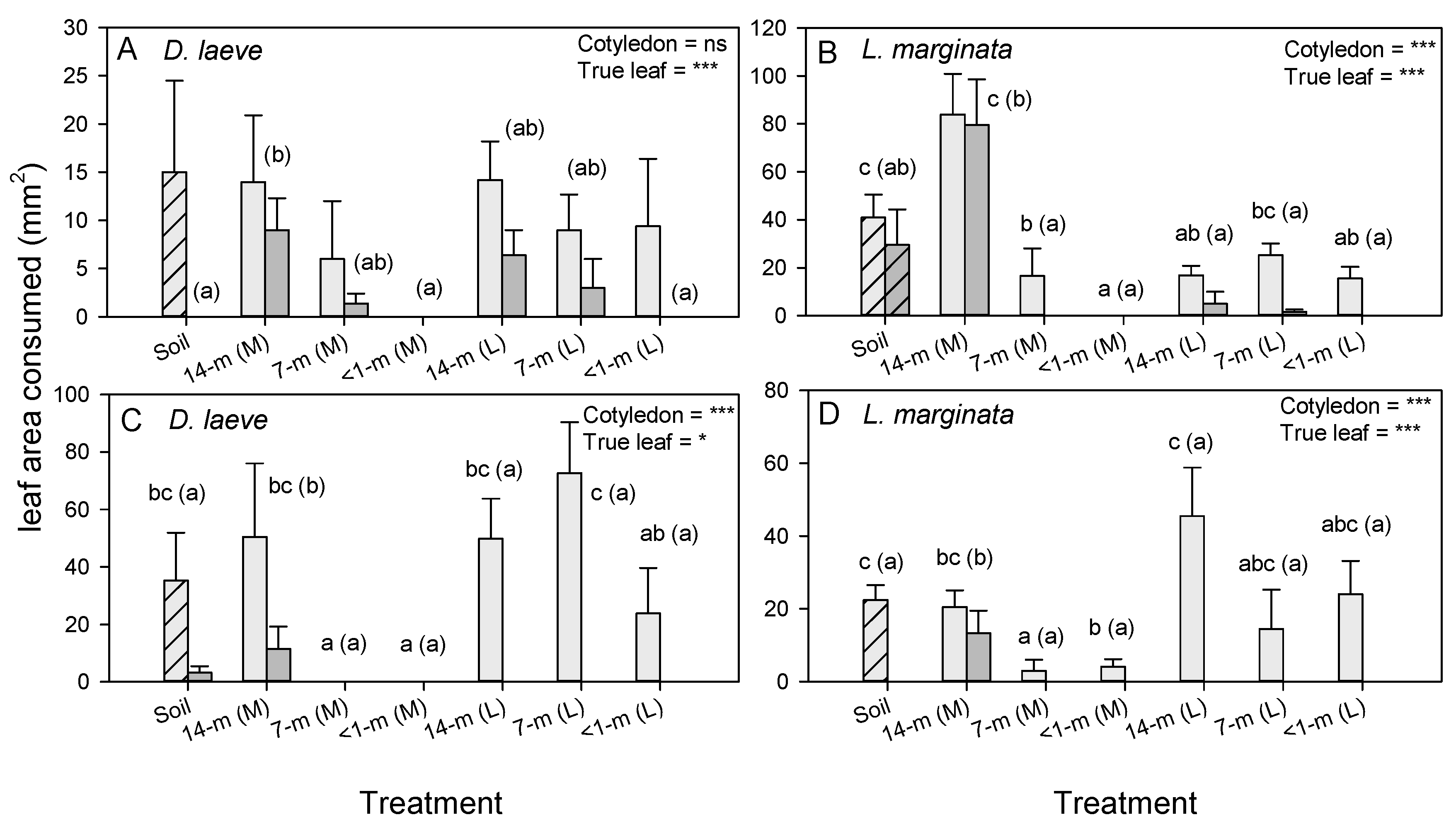
3.5. Leaf-Quality Effects on Slug Herbivory on Radish
3.6. Leaf-Quality Effects on Slug Herbivory on Tomato
4. Discussion
4.1. Effects of SCG on BSF Larval Growth and Development
4.2. Effects of SCG on Plant Growth
4.3. Effects of SCG on Slug Herbivory
4.4. Recommendations
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Schroeder, P.; Anggraeni, K.; Weber, U. The relevance of circular economy practices to the Sustainable Development Goals. J. Ind. Ecol. 2019, 23, 77–95. [Google Scholar] [CrossRef] [Green Version]
- Rosemarin, A.; Macura, B.; Carolus, J.; Barquet, K.; Ek, F.; Järnberg, L.; Lorick, D.; Johannesdottir, S.; Pedersen, S.M.; Koskiaho, J.; et al. Circular nutrient solutions for agriculture and wastewater – a review of technologies and practices. Curr. Opin. Environ. Sustain. 2020, 45, 78–91. [Google Scholar] [CrossRef]
- Diacono, M.; Persiani, A.; Testani, E.; Montemurro, F.; Ciaccia, C. Recycling agricultural wastes and by-products in organic farming: Biofertilizer production, yield performance and carbon footprint analysis. Sustainability 2019, 11, 3824. [Google Scholar] [CrossRef] [Green Version]
- Duque-Acevedo, M.; Belmonte-Ureña, L.J.; Plaza-Úbeda, J.A.; Camacho-Ferre, F. The management of agricultural waste biomass in the framework of circular economy and bioeconomy: An opportunity for greenhouse agriculture in southeast Spain. Agronomy 2020, 10, 489. [Google Scholar] [CrossRef] [Green Version]
- Dahlan, I.; Yusoff, M.S.; Akinbile, C.O.; Wang, L.K.; Wang, M.-H.S. Restaurant waste recycle and disposal. In Solid Waste Engineering and Management; Wang, L.K., Wang, M.-H.S., Hung, Y.-T., Eds.; Springer International Publishing: Cham, Switzerland, 2022; Volume 2, pp. 439–542. [Google Scholar]
- Voora, V.; Bermúdez, S.; Larrea, C.; Baliño, S. Global Market Report: Coffee; International Institute for Sustainable Development: Winnipeg, MB, Canada, 2019; p. 6. [Google Scholar]
- Rajesh Banu, J.; Kavitha, S.; Yukesh Kannah, R.; Dinesh Kumar, M.; Preethi; Atabani, A.E.; Kumar, G. Biorefinery of spent coffee grounds waste: Viable pathway towards circular bioeconomy. Bioresour. Technol. 2020, 302, 122821. [Google Scholar] [CrossRef] [PubMed]
- Murthy, P.S.; Madhava Naidu, M. Sustainable management of coffee industry by-products and value addition—A review. Resour. Conserv. Recycl. 2012, 66, 45–58. [Google Scholar] [CrossRef]
- Kamil, M.; Ramadan, K.M.; Awad, O.I.; Ibrahim, T.K.; Inayat, A.; Ma, X. Environmental impacts of biodiesel production from waste spent coffee grounds and its implementation in a compression ignition engine. Sci. Total Environ. 2019, 675, 13–30. [Google Scholar] [CrossRef]
- Afriliana, A.; Hidayat, E.; Mitoma, Y.; Masuda, T.; Harada, H. Studies on composting spent coffee grounds by Aspergillus sp and Aspergillus sp in aerobic static batch temperature control. J. Agric. Chem. Environ. 2020, 10, 91–112. [Google Scholar]
- Mustard, J.A. The buzz on caffeine in invertebrates: Effects on behavior and molecular mechanisms. Cell. Mol. Life Sci. 2014, 71, 1375–1382. [Google Scholar] [CrossRef] [Green Version]
- Campos-Vega, R.; Loarca-Piña, G.; Vergara-Castañeda, H.A.; Oomah, B.D. Spent coffee grounds: A review on current research and future prospects. Trends Food Sci. Technol. 2015, 45, 24–36. [Google Scholar] [CrossRef]
- Kourmentza, C.; Economou, C.N.; Tsafrakidou, P.; Kornaros, M. Spent coffee grounds make much more than waste: Exploring recent advances and future exploitation strategies for the valorization of an emerging food waste stream. J. Clean. Prod. 2018, 172, 980–992. [Google Scholar] [CrossRef]
- Cervera-Mata, A.; Pastoriza, S.; Rufián-Henares, J.Á.; Párraga, J.; Martín-García, J.M.; Delgado, G. Impact of spent coffee grounds as organic amendment on soil fertility and lettuce growth in two Mediterranean agricultural soils. Arch. Agron. Soil Sci. 2018, 64, 790–804. [Google Scholar] [CrossRef]
- Chen, X.-E.; Mangindaan, D.; Chien, H.-W. Green sustainable photothermal materials by spent coffee grounds. J. Taiwan Inst. Chem. Eng. 2022, 137, 104259. [Google Scholar] [CrossRef]
- Chien, H.-W.; Chen, X.-E. Spent coffee grounds as potential green photothermal materials for biofilm elimination. J. Environ. Eng. 2022, 10, 107131. [Google Scholar] [CrossRef]
- Cruz, R.; Baptista, P.; Cunha, S.; Pereira, J.A.; Casal, S. Carotenoids of lettuce (Lactuca sativa L.) grown on soil enriched with spent coffee grounds. Molecules 2012, 17, 1535–1547. [Google Scholar] [CrossRef] [Green Version]
- Chilosi, G.; Aleandri, M.P.; Luccioli, E.; Stazi, S.R.; Marabottini, R.; Morales-Rodríguez, C.; Vettraino, A.M.; Vannini, A. Suppression of soil-borne plant pathogens in growing media amended with espresso spent coffee grounds as a carrier of Trichoderma spp. Sci. Hortic. 2020, 259, 108666. [Google Scholar] [CrossRef]
- Ciesielczuk, T.; Rosik-Dulewska, C.; Poluszyńska, J.; Miłek, D.; Szewczyk, A.; Sławińska, I. Acute toxicity of experimental fertilizers made of spent coffee grounds. Waste Biomass Valorization 2018, 9, 2157–2164. [Google Scholar] [CrossRef] [Green Version]
- Hardgrove, S.J.; Livesley, S.J. Applying spent coffee grounds directly to urban agriculture soils greatly reduces plant growth. Urban For. Urban Green. 2016, 18, 1–8. [Google Scholar] [CrossRef]
- Yamane, K.; Kono, M.; Fukunaga, T.; Iwai, K.; Sekine, R.; Watanabe, Y.; Iijima, M. Field evaluation of coffee grounds application for crop growth enhancement, weed control, and soil improvement. Plant Prod. Sci. 2014, 17, 93–102. [Google Scholar] [CrossRef]
- Hirooka, Y.; Kurashige, S.; Yamane, K.; Kakiuchi, M.; Miyagawa, T.; Iwai, K.; Iijima, M. Effects of different application methods of spent coffee grounds on weed growth. Weed Technol. 2022, 36, 692–699. [Google Scholar] [CrossRef]
- Hirooka, Y.; Kurashige, S.; Yamane, K.; Watanabe, Y.; Kakiuchi, M.; Ishikawa, D.; Miyagawa, T.; Iwai, K.; Iijima, M. Effectiveness of direct application of top dressing with spent coffee grounds for soil improvement and weed control in wheat-soybean double cropping system. Plant Prod. Sci. 2022, 25, 148–156. [Google Scholar] [CrossRef]
- Hollingsworth, R.G.; Armstrong, J.W.; Campbell, E. Caffeine as a repellent for slugs and snails. Nature 2002, 417, 915–916. [Google Scholar] [CrossRef]
- Jeong, K.J.; Lee, S.W.; Hong, J.K.; Shin, C.Y.; Yun, J.G. Effective control of slug damage through tobacco extract and caffeine solution in combination with alcohol. Hortic. Environ. Biotechnol. 2012, 53, 123–128. [Google Scholar] [CrossRef]
- Chalker-Scott, L. Impact of mulches on landscape plants and the environment—A review. J. Environ. Hortic. 2007, 25, 239–249. [Google Scholar] [CrossRef]
- Chalker-Scott, L. Coffee Grounds–Will They Perk up Plants? Puyallup Research and Extension Center, Washington State University: Pullman, WA, USA, 2009; pp. 3–4. [Google Scholar]
- Cervera-Mata, A.; Navarro-Alarcón, M.; Rufián-Henares, J.Á.; Pastoriza, S.; Montilla-Gómez, J.; Delgado, G. Phytotoxicity and chelating capacity of spent coffee grounds: Two contrasting faces in its use as soil organic amendment. Sci. Total Environ. 2020, 717, 137247. [Google Scholar] [CrossRef]
- El-Saied, R.M.; Rashwan, B.R.A. Combined application of various sources of organic fertilizers with biofertilizers for improvement potato productivity and soil fertility status. Environ. Biodivers. Soil Secur. 2021, 5, 155–170. [Google Scholar] [CrossRef]
- Ronga, D.; Pane, C.; Zaccardelli, M.; Pecchioni, N. Use of spent coffee ground compost in peat-based growing media for the production of basil and tomato potting plants. Commun. Soil Sci. Plant Anal. 2016, 47, 356–368. [Google Scholar] [CrossRef]
- Liu, K.; Price, G.W. Evaluation of three composting systems for the management of spent coffee grounds. Bioresour. Technol. 2011, 102, 7966–7974. [Google Scholar] [CrossRef]
- Santos, C.; Fonseca, J.; Aires, A.; Coutinho, J.; Trindade, H. Effect of different rates of spent coffee grounds (SCG) on composting process, gaseous emissions and quality of end-product. Waste Manag. 2017, 59, 37–47. [Google Scholar] [CrossRef]
- Shin, J.-H.; Park, S.-H.; Kim, A.; Son, Y.-H.; Joo, S.-H. Changes in physical, chemical, and biological traits during composting of spent coffee grounds. Korean J. Environ. Agric. 2020, 39, 178–187. [Google Scholar] [CrossRef]
- Fischer, H.; Romano, N.; Sinha, A.K. Conversion of spent coffee and donuts by Black Soldier Fly (Hermetia illucens) larvae into potential resources for animal and plant farming. Insects 2021, 12, 332. [Google Scholar] [CrossRef] [PubMed]
- Permana, A.D.; Putra, J.E.N.R.E. Growth of black soldier fly (Hermetia illucens) larvae fed on spent coffee ground. IOP Conf. Ser. Earth Environ. Sci. 2018, 187, 012070. [Google Scholar] [CrossRef]
- Barragan-Fonseca, K.B.; Dicke, M.; van Loon, J.J. Nutritional value of the black soldier fly (Hermetia illucens L.) and its suitability as animal feed–a review. J. Insects Food Feed 2017, 3, 105–120. [Google Scholar] [CrossRef]
- Broeckx, L.; Frooninckx, L.; Slegers, L.; Berrens, S.; Noyens, I.; Goossens, S.; Verheyen, G.; Wuyts, A.; Van Miert, S. Growth of black soldier fly larvae reared on organic side-streams. Sustainability 2021, 13, 12953. [Google Scholar] [CrossRef]
- Barragán-Fonseca, K.Y.; Nurfikari, A.; van de Zande, E.M.; Wantulla, M.; van Loon, J.J.A.; de Boer, W.; Dicke, M. Insect frass and exuviae to promote plant growth and health. Trends Plant Sci. 2022, 27, 646–654. [Google Scholar] [CrossRef]
- Borkent, S.; Hodge, S. Glasshouse evaluation of the black soldier fly waste product HexaFrass™ as an organic fertilizer. Insects 2021, 12, 977. [Google Scholar] [CrossRef]
- Lopes, I.G.; Yong, J.W.H.; Lalander, C. Frass derived from black soldier fly larvae treatment of biodegradable wastes. A critical review and future perspectives. Waste Manag. 2022, 142, 65–76. [Google Scholar] [CrossRef]
- Cervera-Mata, A.; Delgado, G.; Fernández-Arteaga, A.; Fornasier, F.; Mondini, C. Spent coffee grounds by-products and their influence on soil C–N dynamics. J. Environ. Manag. 2022, 302, 114075. [Google Scholar] [CrossRef]
- Cervera-Mata, A.; Navarro-Alarcón, M.; Delgado, G.; Pastoriza, S.; Montilla-Gómez, J.; Llopis, J.; Sánchez-González, C.; Rufián-Henares, J.Á. Spent coffee grounds improve the nutritional value in elements of lettuce (Lactuca sativa L.) and are an ecological alternative to inorganic fertilizers. Food Chem. 2019, 282, 1–8. [Google Scholar] [CrossRef]
- Surendra, K.C.; Tomberlin, J.K.; van Huis, A.; Cammack, J.A.; Heckmann, L.-H.L.; Khanal, S.K. Rethinking organic wastes bioconversion: Evaluating the potential of the black soldier fly (Hermetia illucens (L.)) (Diptera: Stratiomyidae) (BSF). Waste Manag. 2020, 117, 58–80. [Google Scholar] [CrossRef]
- Barragan-Fonseca, K.B.; Dicke, M.; van Loon, J.J.A. Influence of larval density and dietary nutrient concentration on performance, body protein, and fat contents of black soldier fly larvae (Hermetia illucens). Entomol. Exp. Appl. 2018, 166, 761–770. [Google Scholar] [CrossRef]
- Cohn, Z.; Latty, T.; Abbas, A. Understanding dietary carbohydrates in black soldier fly larvae treatment of organic waste in the circular economy. Waste Manag. 2022, 137, 9–19. [Google Scholar] [CrossRef]
- Horgan, F.G.; Launders, M.; Mundaca, E.A.; Crisol-Martínez, E. Effects of interspecific competition and larval size on bioconversion of apple pomace inoculated with black soldier fly. 2023, accepted.
- Hadj Saadoun, J.; Montevecchi, G.; Zanasi, L.; Bortolini, S.; Macavei, L.I.; Masino, F.; Maistrello, L.; Antonelli, A. Lipid profile and growth of black soldier flies (Hermetia illucens, Stratiomyidae) reared on by-products from different food chains. J. Sci. Food Agric. 2020, 100, 3648–3657. [Google Scholar] [CrossRef]
- Tegtmeier, D.; Hurka, S.; Klüber, P.; Brinkrolf, K.; Heise, P.; Vilcinskas, A. Cottonseed press cake as a potential diet for industrially farmed black soldier fly larvae triggers adaptations of their bacterial and fungal gut microbiota. Front. Microbiol. 2021, 12, 634503. [Google Scholar] [CrossRef]
- Fenoll, J.; Ruiz, E.; Flores, P.; Vela, N.; Hellín, P.; Navarro, S. Use of farming and agro-industrial wastes as versatile barriers in reducing pesticide leaching through soil columns. J. Hazard. Mater. 2011, 187, 206–212. [Google Scholar] [CrossRef]
- Chrysargyris, A.; Antoniou, O.; Xylia, P.; Petropoulos, S.; Tzortzakis, N. The use of spent coffee grounds in growing media for the production of Brassica seedlings in nurseries. Environ. Sci. Pollut. Res. 2021, 28, 24279–24290. [Google Scholar] [CrossRef]
- Girotto, F.; Pivato, A.; Cossu, R.; Nkeng, G.E.; Lavagnolo, M.C. The broad spectrum of possibilities for spent coffee grounds valorisation. J. Mater. Cycles Waste Manag. 2018, 20, 695–701. [Google Scholar] [CrossRef]
- Turek, M.E.; Freitas, K.S.; Armindo, R.A. Spent coffee grounds as organic amendment modify hydraulic properties in a sandy loam Brazilian soil. Agric. Water Manag. 2019, 222, 313–321. [Google Scholar] [CrossRef]
- Gomes, T.; Pereira, J.A.; Ramalhosa, E.; Casal, S.; Baptista, P. Effect of fresh and composted spent coffee grounds on lettuce growth, photosynthetic pigments and mineral composition. In Proceedings of the VII Congreso Ibérico de Agroingeniería y Ciencias Horticolas, Madrid, Spain, 26-29 August 2013; pp. 1–5. [Google Scholar]
- Cruz, R.; Gomes, T.; Ferreira, A.; Mendes, E.; Baptista, P.; Cunha, S.; Pereira, J.A.; Ramalhosa, E.; Casal, S. Antioxidant activity and bioactive compounds of lettuce improved by espresso coffee residues. Food Chem. 2014, 145, 95–101. [Google Scholar] [CrossRef]
- Cruz, R.; Mendes, E.; Torrinha, Á.; Morais, S.; Pereira, J.A.; Baptista, P.; Casal, S. Revalorization of spent coffee residues by a direct agronomic approach. Food Res. Int. 2015, 73, 190–196. [Google Scholar] [CrossRef]
- Hanc, A.; Hrebeckova, T.; Grasserova, A.; Cajthaml, T. Conversion of spent coffee grounds into vermicompost. Bioresour. Technol. 2021, 341, 125925. [Google Scholar] [CrossRef]
- Chuang, Y.-H.; Liu, C.-H.; Hammerschmidt, R.; Zhang, W.; Boyd, S.A.; Li, H. Metabolic demethylation and oxidation of caffeine during uptake by lettuce. J. Agric. Food Chem. 2018, 66, 7907–7915. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.Q.; Hua, T.; Gersberg, R.M.; Zhu, J.; Ng, W.J.; Tan, S.K. Fate of caffeine in mesocosms wetland planted with Scirpus validus. Chemosphere 2013, 90, 1568–1572. [Google Scholar] [CrossRef] [PubMed]
- Kang, W.H.; Joo, J.H.; Yoon, B.S.; Kim, I.S. Caffeine as a killing and repellent agent for snails (Acusta despecta) and slugs (Deroceras varians) on horticultural plants. J. Bio-Environ. Control 2002, 11, 149–150. [Google Scholar]
- Das, B.K. Efficacy of some plant derived molluscicides for controlling pestiferous slug, Laevicaulis alte in Odisha. J. Emerg. Technol. Innov. Res. 2022, 9, 468–478. [Google Scholar]
- de Brito, F.C.; Arenzon, A.; Reichert, N.; Lazzarotto, M.; Ávila, S.; Pacheco, S.D.G.; Kruger, I.; Rozino, L.; Vargas, V.M.F. Ecotoxicological evaluation of fruit extracts from yerba mate progenies (Ilex paraguariensis a St-Hil.): A natural biopesticide. J. Environ. Sci. Health Part B 2021, 56, 782–791. [Google Scholar] [CrossRef]
- Castle, G.; Mills, G.; Gravell, A.; Jones, L.; Townsend, I.; Cameron, D.; Fones, G. Review of the molluscicide metaldehyde in the environment. Environ. Sci. Water Res. Technol. 2017, 3, 415–428. [Google Scholar] [CrossRef] [Green Version]
- Horgan, F.G.; Bernal, C.C.; Letana, S.; Naredo, A.I.; Ramp, D.; Almazan, M.L.P. Reduced efficiency of tropical flies (Diptera) in the decomposition of snail cadavers following molluscicide poisoning. Appl. Soil Ecol. 2018, 129, 61–71. [Google Scholar] [CrossRef]
- Langan, A.; Pilkington, G.; Wheater, C. Feeding preferences of a predatory beetle (Pterostichus madidus) for slugs exposed to lethal and sub-lethal dosages of metaldehyde. Entomol. Exp. Appl. 2001, 98, 245–248. [Google Scholar] [CrossRef]
- Langan, A.M.; Taylor, A.; Wheater, C.P. Effects of metaldehyde and methiocarb on feeding preferences and survival of a slug predator (Pterostichus melanarius (F.): Carabidae, Pterostichini). J. Appl. Entomol. 2004, 128, 51–55. [Google Scholar] [CrossRef]









Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Horgan, F.G.; Floyd, D.; Mundaca, E.A.; Crisol-Martínez, E. Spent Coffee Grounds Applied as a Top-Dressing or Incorporated into the Soil Can Improve Plant Growth While Reducing Slug Herbivory. Agriculture 2023, 13, 257. https://doi.org/10.3390/agriculture13020257
Horgan FG, Floyd D, Mundaca EA, Crisol-Martínez E. Spent Coffee Grounds Applied as a Top-Dressing or Incorporated into the Soil Can Improve Plant Growth While Reducing Slug Herbivory. Agriculture. 2023; 13(2):257. https://doi.org/10.3390/agriculture13020257
Chicago/Turabian StyleHorgan, Finbarr G., Dylan Floyd, Enrique A. Mundaca, and Eduardo Crisol-Martínez. 2023. "Spent Coffee Grounds Applied as a Top-Dressing or Incorporated into the Soil Can Improve Plant Growth While Reducing Slug Herbivory" Agriculture 13, no. 2: 257. https://doi.org/10.3390/agriculture13020257
APA StyleHorgan, F. G., Floyd, D., Mundaca, E. A., & Crisol-Martínez, E. (2023). Spent Coffee Grounds Applied as a Top-Dressing or Incorporated into the Soil Can Improve Plant Growth While Reducing Slug Herbivory. Agriculture, 13(2), 257. https://doi.org/10.3390/agriculture13020257