

Surrounding Semi-Natural Vegetation as a Source of Aphidophagous Syrphids (Diptera, Syrphidae) for Aphid Control in Apple Orchards


Abstract
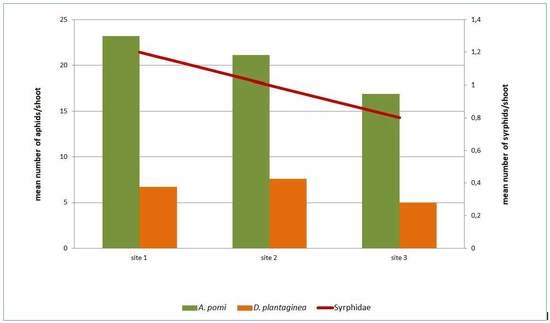
:
1. Introduction
2. Materials and Methods
2.1. Research Sites
2.2. Method of Sampling
2.2.1. Sampling of Aphids
2.2.2. Sampling of Syrphidae
Yellow Traps
Hand Picking
2.3. Statistical Analysis
3. Results
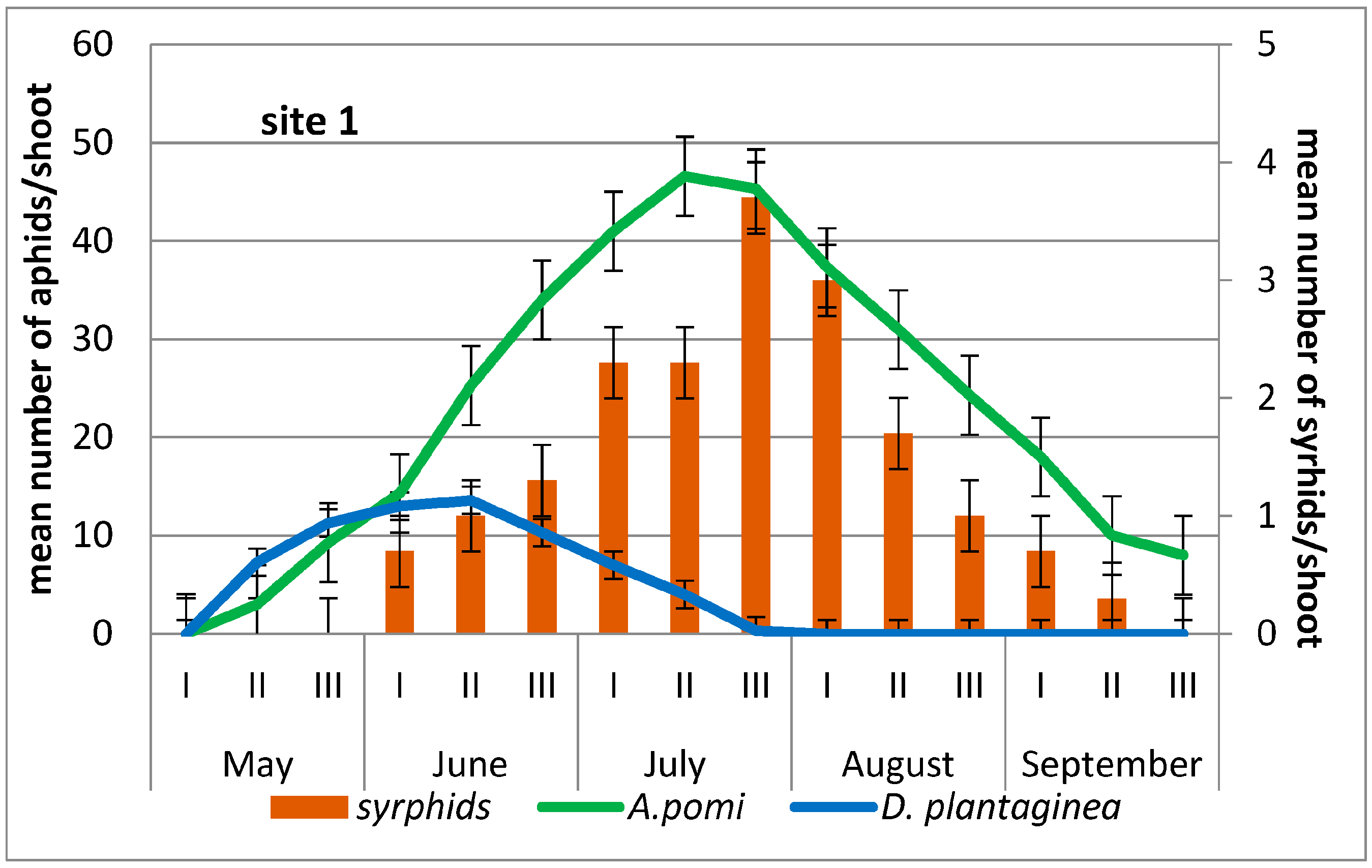
3.1. Occurrence of D. plantaginea and A. pomi on Apple Trees
3.2. Syrphids Collected from the Aphid Colonies
3.3. Syrphid Collected in Yellow Traps
3.4. Significant Difference and Correlations between the Syrphidae from Different Habitats
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Foley, J.A.; Ramankutty, N.; Brauman, K.A.; Cassidy, E.S.; Gerber, J.S.; Johnston, M.; Mueller, N.D.; O’Connell, C.; Ray, D.K.; West, O.C.; et al. Solutions for a cultivated planet. Nature 2011, 478, 337–342. [Google Scholar] [CrossRef] [PubMed]
- Wratten, S.D.; Gillespie, M.; Decourtye, A.; Mader, E.; Desneux, N. Pollinator habitat enhancement: Benefits to other ecosystem services. Agric. Ecosyst. Environ. 2012, 159, 112–122. [Google Scholar] [CrossRef]
- Altieri, M.A.; Nicholls, A.H. Biodiversity and Pest Management in Agroecosystems, 2nd ed.; Haworth Food & Agricultural Products Press, Inc.: Binghamton, NY, USA, 2004. [Google Scholar]
- Simon, S.; Bouvier, J.C.; Debras, J.F.; Sauphanor, B. Biodiversity and pest management in orchard systems: A review. Agron. Sustain. Dev. 2010, 30, 139–152. [Google Scholar] [CrossRef]
- Gurr, G.M.; Wratten, S.D.; Landis, D.A.; You, M.S. Habitat Management to Suppress Pest Populations: Progress and Prospects. Annu. Rev. Entomol. 2017, 62, 91–109. [Google Scholar] [CrossRef]
- Rodríguez-Gasol, N.; Avilla, J.; Aparicio, Y.; Arnó, J.; Gabarra, R.; Riudavets, J.; Alegre, S.; Lordan, J.; Alins, G. The Contribution of Surrounding Margins in the Promotion of Natural Enemies in Mediterranean Apple Orchards. Insects 2019, 10, 148. [Google Scholar] [CrossRef] [PubMed]
- Fiedler, Z.; Sosnowska, D. Side effects of fungicides and insecticides on predatory mites, in laboratory conditions. J. Plant Prot. Res. 2014, 54, 349–353. [Google Scholar] [CrossRef]
- De França, S.M.; Breda, M.O.; Barbosa, D.R.; Araujo, A.M.; Guedes, C.A. The sublethal effects of insecticides in insects. In Biological Control of Pest and Vector Insects; Shields, V.D.C., Ed.; Intech Open: London, UK, 2017; Chapter 2; pp. 23–39. [Google Scholar] [CrossRef]
- Guzmán, G.; Ruiz, R.G. Side effects of insecticides on beneficial insects: A practical tool to identify organic agroecosystems. WJASS 2019, 4, 1–5. [Google Scholar] [CrossRef]
- Wojciechowicz-Żytko, E.; Wilk, E. The quantity and quality structure of coccinellids (Coleoptera, Coccinellidae) in apple orchards with integrated and ecological management and in their surroundings. Pol. J. Environ. Stud. 2022, 31, 2895–2903. [Google Scholar] [CrossRef]
- Belyakov, N.V.; Nikolina, N. Plant protection technologies: From advanced to innovative. JPCS 2021, 1942, 012072. [Google Scholar] [CrossRef]
- Pobożniak, M.; Gaborska, M.; Wójtowicz, T. Resistance and tolerance of ten carrot cultivars to the hawthorn-carrot aphid, Dysaphis crataegi Kalt., in Poland. PLoS ONE 2021, 16, e0247978. [Google Scholar] [CrossRef]
- Rousselin, A.; Bevacqua, D.; Sauge, M.H.; Lescourret, F.; Mody, K.; Jordan, M.O. Harnessing the aphid life cycle to reduce insecticide reliance in apple and peach orchards. A review. Agron. Sustain. Dev. 2017, 37, 38. [Google Scholar] [CrossRef]
- Lefebvre, V.; Fontaine, C.; Villemant, C.; Daugeron, C. Are empidine dance flies major flower visitors in alpine environments? A case study in the Alps, France. Biol. Lett. 2014, 10, 20140742. [Google Scholar] [CrossRef]
- A lbert, L.; Franck, P.; Gilles, Y.; Plantegenest, M. Impact of groecological infrastructures on the dynamics of Dysaphis plantaginea (Hemiptera: Aphididae) and its natural enemies in apple orchards in northwestern France. Environ. Entomol. 2017, 46, 528–537. [Google Scholar] [CrossRef] [PubMed]
- Wojciechowicz-Żytko, E. Attractiveness of some Apiaceae flowers for Syrphidae (Diptera)—Pollinators and biotic agents. Acta Hortic. 2019, 1264, 275–282. [Google Scholar] [CrossRef]
- Rodriguez-Gasol, N.; Alins, G.; Veronesi, E.; Wratten, S. The ecology of hoverflies as ecosystem-service providers in agricultural systems. Biol. Control 2020, 151, 104405. [Google Scholar] [CrossRef]
- Miller, N.; Al-Dobaib, S.; Legaspia, J.; Sivinski, J. Estimating attraction of Syrphidae (Diptera) to flowering plants with interception traps. BioControl Sci. Technol. 2013, 23, 1040–1052. [Google Scholar] [CrossRef]
- Miñarro, M.; Prida, E. Hedgerows surrounding organic apple orchards in north-west Spain:potential to conserve beneficial insects. Agric. For. Entomol. 2013, 15, 382–390. [Google Scholar] [CrossRef]
- Zhou, H.X.; Yu, Y.; Tan, X.M.; Chen, A.D.; Feng, J.G. Biological control of insect pests in apple orchards in China. Biol. Control 2014, 68, 47–56. [Google Scholar] [CrossRef]
- Gontijo, L.M.; Beers, E.H.; Snyder, W.E. Complementary suppression of aphids by predators and parasitoids. Biol. Control 2015, 90, 83–91. [Google Scholar] [CrossRef]
- Campbell, A.J.; Wilby, A.; Sutton, P.; Wackers, F. Getting more power from your flowers: Multi-functional flower strips enhance pollinators and pest control agents in apple orchards. Insects 2017, 8, 101. [Google Scholar] [CrossRef]
- Cahenzli, F.; Pfiffner, L.; Daniel, C. Reduced crop damage by self-regulation of aphids in an ecologically enriched, insecticide-free apple orchard. Agron. Sustain. Dev. 2017, 37, 65. [Google Scholar] [CrossRef]
- Woltz, J.M.; Landis, D.A. Coccinellid response to landscape composition and configuration. Agric. For. Entomol. 2014, 16, 341–349. [Google Scholar] [CrossRef]
- Holland, J.M.; Bianchi, F.J.J.A.; Entling, M.H.; Moonen, A.C.; Smith, B.M.; Jeanneret, P. Structure, function and management of semi-natural habitats for conservation biological control: A review of European studies. Pest Manag. Sci. 2016, 72, 1638–1651. [Google Scholar] [CrossRef] [PubMed]
- Wojciechowicz-Żytko, E.; Jankowska, B. Sambucus nigra L. as a reservoir of beneficial insects (Diptera, Syrphidae). Folia Hortic. 2016, 28, 209–216. [Google Scholar] [CrossRef]
- Landis, D.A.; Wratten, S.D.; Gurr, G.M. Habitat management to conserve natural enemies of arthropod pests in agriculture. Annu. Rev. Entomol. 2000, 45, 175–201. [Google Scholar] [CrossRef]
- Odorizzi Santos, L.A.; Costa, M.B.; Lavigne, C.; Fernandes, O.A.; Bischoff, A.; Franck, P. Influence of the margin vegetation on the conservation of aphid biological control in apple orchards. J. Insect Conserv. 2018, 22, 465–475. [Google Scholar] [CrossRef]
- A lbrecht, M.; Kleijn, D.; Williams, N.M.; Tschumi, M.; Blaauw, B.R.; Bommarco, R.; Campbell, A.J.; Dainese, M.; Drummond, F.A.; Entling, M.H.; et al. The effectiveness of flower strips and hedgerows on pest control, pollination services and crop yield: A quantitative synthesis. Ecol. Lett. 2020, 23, 1488–1498. [Google Scholar] [CrossRef]
- Bianchi, F.; Booij, C.; Tscharntke, T. Sustainable pest regulation in agricultural landscapes: A review on landscape composition, biodiversity and natural pest control. Proc. R. Soc. B 2006, 273, 1715–1727. [Google Scholar] [CrossRef]
- Chaplin-Kramer, R.; O’Rourke, M.; Blitzer, E.; Kremen, C. A meta-analizys of crop pest and natural enemy response to landscape complexity. Ecol. Lett. 2011, 14, 922–932. [Google Scholar] [CrossRef]
- Shackelford, G.; Steward, P.; Benton, T.; Kunin, E.; Potts, S.; Biesmeijer, J.; Sait, S. Comparison of pollinators and natural enemies: A meta analysis of landscape and local effects on abundance and richness in crops. Biol. Rev. 2013, 88, 1002–1021. [Google Scholar] [CrossRef]
- Blitzer, E.; Dormann, C.; Holzschuh, A.; Tscharntke, T.; Klein, A.; Rand, T. Spillover of functionally important organisms between managed and natural habitats. Agric. Ecosyst. Environ. 2012, 146, 34–43. [Google Scholar] [CrossRef]
- Macfadyen, S.; Muller, W. Edges in Agricultural Landscapes: Species Interactions and Movement of Natural Enemies. PLoS ONE 2013, 8, e59659. [Google Scholar] [CrossRef]
- Garratt, M.; Senapathi, D.; Coston, D.; Mortimer, S.; Potts, S. The benefits of hedgerows for pollinators and natural enemies depends on hedge quality and landscape context. Agric. Ecosyst. Environ. 2017, 247, 363–370. [Google Scholar] [CrossRef]
- Happea, A.-K.; Alinsc, G.; Blüthgena, N.; Boreuxd, V.; Bosche, J.; Garcíaf, D.; Hambäckg, P.; Kleind, A.-M.; Martínez-Sastreh, R.; Miñarroh, M.; et al. Predatory arthropods in apple orchards across Europe: Responses to agricultural management, adjacent habitat, landscape composition and country. Agric. Ecosyst. Environ. 2019, 273, 141–150. [Google Scholar] [CrossRef]
- Daelemans, R.; Hulsmans, E.; Laenen, E.; Remy, E.; Beliën, T.; Honnay, O. Direct and indirect effects of management and landscape on biological pest control and crop pest infestation in apple orchards. J. Appl. Ecol. 2023, 60, 181–192. [Google Scholar] [CrossRef]
- Moericke, V. Wie finden geflügelte Blattläuse ihre Wirtspflanze? In Angewandte Chemie; Mitteilungen aus der Biologischen Reichsanstalt: Berlin, Germany, 1953; Volume 75, p. 90. [Google Scholar]
- Van Veen, M. Hoverflies of Northwest Europe: Identification Keys to the Syrphidae; KNNV. Publishing: Utrecht, The Netherlands, 2004. [Google Scholar]
- Rotheray, G.E. Colour Guide to Hoverfly Larvae (Diptera, Syrphidae) in Britain and Europe. Diperists Dig. 1993, 9, 1–156. [Google Scholar]
- Soszyński, B. Syrphidae. In Checklist of Animals of Poland; Razowski, J., Ed.; Institute of Systematics and Evolution of Animals, Polish Academy of Sciences: Krakow, Poland, 1991. [Google Scholar]
- Kasprzak, K.; Niedbała, W. Biocenotic indicators used in ordering and analyzing data in quantitative research. In Methods Used in Soil Zoology; Górny, M., Grum, L., Eds.; Scientific PWN: Warsaw, Poland, 1981. [Google Scholar]
- Chao, A.; Chazdon, R.L.; Colwell, R.K.; Shen, T.-S. A new statistical approach for assessing 357 similarity of species composition with incidence and abundance data. Ecol. Lett. 2005, 8, 148–149. [Google Scholar] [CrossRef]
- Debras, J.F.; Torre, F.; Rieux, R.; Kreiter, S.; Garcin, M.S.; Van Helden, M.; Buisson, E.; Dutoit, T. Discrimination between agricultural management and the hedge effect in pear orchards (south-eastern France). Ann. Appl. Biol. 2006, 149, 347–355. [Google Scholar] [CrossRef]
- Horton, D.R.; Broers, D.A.; Lewis, R.R.; Granatstein, D.; Zack, R.S.; Unruh, T.R.; Moldenke, A.R.; Brown, J.J. Effects of mowing frequency on densities of natural enemies in three Pacific Northwest pear orchards. Entomol. Exp. Appl. 2003, 106, 135–145. [Google Scholar] [CrossRef]
- Boller, E.F.; Häni, F.; Poehling, H.M. Ecological Infrastructures: Ideabook on Functional Diversity at the Farm Level; IOBC WPRS: Winterthur, Switzerland, 2004. [Google Scholar]
- Lindgren, J.; Lindborg, R.; Cousins, S.A.O. Local conditions in small habitats and surrounding landscape are important for pollination services, biological pest control and seed predation. Agric. Ecosyst. Environ. 2018, 251, 107–113. [Google Scholar] [CrossRef]
- Piekarska-Boniecka, H.; Siatkowski, M.; Trzciński, P.; Siatkowski, I. The impact of the vegetation of apple orchard edges on quantity and quality structure of predatory hoverflies (Diptera: Syrphidae) communities. Turk. Entomol. Derg. 2015, 39, 333–343. [Google Scholar] [CrossRef]
- Trzciński, P.; Piekarska-Boniecka, H. Dynamics of predatory Syrphidae in the apple orchard and neighbouring shrubberies. J. Plant Prot. Res. 2013, 53, 119–123. [Google Scholar] [CrossRef]
- Rossi, J.; Gamba, U.; Pinna, M.; Spagnolo, S.; Visentin, J.; Alma, A. Hoverflies in organic apple orchards in north-western Italy. Bull. Insectol. 2006, 59, 111–114. [Google Scholar]
- Miñarro, M.; Hemptinne, J.L.; Dapena, E. Colonization of apple orchards by predators of Dysaphis plantaginea: Sequential arrival, response to prey abundance and consequences for biological control. BioControl 2005, 50, 403–414. [Google Scholar] [CrossRef]
- Dib, H.; Sauphanor, B.; Capowiez, Y. Effect of management strategies on arthropod communities in the colonies of rosy apple aphid, Dysaphis plantaginea Passerini (Hemiptera: Aphididae) in south-eastern France. Agric. Ecosyst. Environ. 2016, 216, 203–206. [Google Scholar] [CrossRef]
- Kumari, M. Biology and feeding potential Episyrphus balteatus De Geer (Diptera: Syrphidae) on green apple aphid Aphis pomi De Geer (order Hemiptera: Aphididae) in Hills of Shimla, (H.P.), India. Environ. Conserv. 2020, 21, 147–150. [Google Scholar] [CrossRef]
- Aparicio, Y.; Riudavets, J.; Gabarra, R.; Agustín, N.; Rodríguez-Gasol, N.; Alins, G.; Blasco-Morenoa., A.; Arnó, J. Can Insectary Plants Enhance the Presence of Natural Enemies of the Green Peach Aphid (Hemiptera: Aphididae) in Mediterranean Peach Orchards? J. Econ. Entomol. 2021, 114, 784–793. [Google Scholar] [CrossRef]
- Khan, A.A.; Riyaz, S. Diversity and Distribution of Syrphid Fly Communities in Temperate Fruit Orchard of Kashmir, India. IJCMAS 2017, 6, 2794–2805. [Google Scholar] [CrossRef]
- Speight, M.C.D. Species accounts of European Syrphidae (Diptera) 2004. In Syrph the Net, the Database of European Syrphidae; Speight, M.C.D., Castella, E., Sarthou, J.-P., Monteil, C., Eds.; Syrph the Net publications: Dublin, Irland, 2010; Volume 44, p. 235. [Google Scholar]
- Naderloo, M.; Rad, S.P. Diversity of Hoverfly (Diptera: Syrphidae) Communities in Different Habitat Types in Zanjan Province, Iran. Int. Sch. Res. Not. 2014, 2014, 162343. [Google Scholar] [CrossRef]
- Dib, H.; Simon, S.; Sauphanor, B.; Capowiez, Y. The role of natural enemies on the population dynamics of the rosy apple aphid, Dysaphis plantaginea Passerini (Hemiptera: Aphididae) in organic apple orchards in south-eastern France. Biol. Control 2010, 55, 97–109. [Google Scholar] [CrossRef]
- Pfiffner, L.; Luka, H.; Schlatter, C.; Juen, A.; Traugott, M. Impact of wild flower strips on biological control of cabbage lepidopterans. Agric. Ecosyst. Environ. 2009, 129, 310–314. [Google Scholar] [CrossRef]
- Tschumi, M.; Albrecht, M.; Enthling, M.H.; Jacot, K. High effec-tiveness of tailored flower strips in reducing pests and crop plantdamage. Proc. R. Soc. B 2015, 282, 20151369. [Google Scholar] [CrossRef] [PubMed]
- Haaland, C.; Naisbit, R.E.; Bersier, L.F. Sown wildflower strips for insect conservation—A review. Insect Conserv. Divers. 2011, 4, 60–80. [Google Scholar] [CrossRef]
- Branquart, E.; Hemptinne, J.L. Selectivity in the exploitation of floral resources by hoverflies (Diptera: Syrphinae). Ecography 2000, 23, 732–742. [Google Scholar] [CrossRef]
- Wojciechowicz-Żytko, E.; Wilk, E. Effects of the Surrounding Environment and Management System in Apple Orchards on the Occurrence of the Ground Beetles (Coleoptera, Carabidae). Pol. J. Environ. Stud. 2019, 28, 3489–3496. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Traps in Orchards | Traps in Surroundings | Aphid Colonies | |
|---|---|---|---|
| Mean number of specimens | |||
| Site1 | 27.00 ± 16.92 | 54.33 ± 13.38 | 18.00 ± 2.08 |
| Site 2 | 52.00 ± 25.18 | 77.33 ± 52.68 | 15.00 ± 2.52 |
| Site 3 | 50.67 ± 20.70 | 81.33 ± 38.83 | 11.67 ± 2.19 |
| Mean number of species | |||
| Site1 | 6.00 ± 2.08 | 9.00 ± 1.73 | 6.33 ± 0.33 |
| Site 2 | 11.00 ± 1.73 | 8.67 ± 1.76 | 5.67 ± 0.88 |
| Site 3 | 4.67 ± 0.33 | 11.33 ± 1.45 | 3.67 ± 0.33 |
| Species | Orchard 1 | Orchard 2 | Orchard 3 | Total | ||||
|---|---|---|---|---|---|---|---|---|
| No | % | No | % | No | % | No | % | |
| Epistrophe eligans (Harr.) | 3 | 5.5 D | 3 | 6.7 D | 6 | 4.5 Sd | ||
| Epistrophe melanostoma (Zett.) | 1 | 1.9 R | 1 | 0.7 Sr | ||||
| Episyrphus balteatus (Deg.) | 24 | 44.4 Eu | 20 | 44.4 Eu | 19 | 54.3 Eu | 63 | 47.0 Eu |
| Eupeodes corollae (Fabr.) | 9 | 16.7 Eu | 6 | 13.3 Eu | 9 | 25.7 Eu | 24 | 17.9 Eu |
| Eupeodes lapponicus (Zett.) | 1 | 1.9 R | 1 | 0.7 Sr | ||||
| Scaeva pyrastri (L.) | 1 | 1.9 R | 2 | 4.4 Sd | 3 | 2.2 Sd | ||
| Sphaerophoria scripta (L.) | 7 | 12.9 Eu | 6 | 13.3 Eu | 3 | 8.6 D | 16 | 11.9 Eu |
| Syrphus ribesii (L.) | 1 | 2.3 R | 1 | 0.7 Sr | ||||
| Syrphus torvus O.-S. | 2 | 3.7 Sd | 2 | 4.4 Sd | 1 | 2.8 Sd | 5 | 3.7 Sd |
| Syrphus vitripennis Meig. | 6 | 11.1 Eu | 5 | 11.2 Eu | 3 | 8.6 D | 14 | 10.4 Eu |
| number of specimens | 54 | 100.0 | 45 | 100.0 | 35 | 100.0 | 134 | 100.0 |
| Number of species | 9 | 8 | 5 | 10 | ||||
| Site 1 | Site 2 | Site 3 | |||||||
|---|---|---|---|---|---|---|---|---|---|
| o | s | a.c. | o | s | a.c. | o | s | a.c. | |
| No of species | 13 | 16 | 9 | 16 | 15 | 8 | 8 | 21 | 5 |
| Species richness | 6.3 | 6.8 | 4.7 | 6.8 | 5.8 | 4.1 | 3.2 | 8.3 | 2.7 |
| Species | Site 1 | Site 2 | Site 3 | Total | ||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Orchard | Surrounding | Orchard | Surrounding | Orchard | Surrounding | |||||||||||||||
| No | % | f | No | % | f | No | % | f | No | % | f | No | % | f | No | % | f | No | % | |
| Chrysotoxum vernale | 3 | 3.7 Sd | 5.6 | 1 | 0.6 Sr | 2.8 | 1 | 0.4 Sr | 2.8 | 1 | 0.7 Sr | 2.8 | 1 | 0.4 Sr | 2.8 | 7 | 0.7 Sr | |||
| Dasysyrphus venustus (Meig.) | 2 | 0.8 Sr | 2.8 | 2 | 0.2 Sr | |||||||||||||||
| Didea fasciata Macq. | 1 | 0.4 Sr | 2.8 | 1 | 0.1 Sr | |||||||||||||||
| Didea intermedia Loew | 1 | 0.6 Sr | 2.8 | 1 | 0.1 Sr | |||||||||||||||
| Epistrophe eligans (Harr.) | 6 | 2.6 Sd | 2.8 | 6 | 0.6 Sr | |||||||||||||||
| Epistrophe melanostoma (Zett.) | 2 | 1.2 R | 5.6 | 31 | 13.4 Eu | 8.3 | 1 | 0.7 Sr | 2.8 | 6 | 2.5 Sd | 2.8 | 40 | 3.9 Sd | ||||||
| Episyrphus balteatus (Deg.) | 44 | 54.3 Eu | 38.9 | 81 | 49.7 Eu | 36.1 | 39 | 24.7 Eu | 36.1 | 43 | 18.6 Eu | 25 | 88 | 57.9 Eu | 33.3 | 125 | 51.2 Eu | 50 | 420 | 40.8 Eu |
| Eupeodes corollae (Fabr.) | 13 | 16.0 Eu | 22.2 | 26 | 16.0 Eu | 22.2 | 29 | 18.4 Eu | 33.3 | 20 | 8.7 | 25 | 27 | 17.8 Eu | 8.3 | 22 | 9.0 | 33.3 | 137 | 13.3 Eu |
| Eupeodes lapponicus (Zett.) | 3 | 3.7 Sd | 5.6 | 1 | 0.6 Sr | 2.8 | 12 | 7.6 D | 11.1 | 2 | 0.9 Sr | 5.6 | 1 | 0.4 Sr | 2.8 | 19 | 1.8 R | |||
| Eupeodes latifasciatus (Macq.) | 1 | 1.2 R | 2.8 | 2 | 1.2 R | 2.8 | 8 | 5.1 D | 8.3 | 14 | 6.1 D | 19.4 | 2 | 1.3 R | 5.6 | 11 | 4.5 Sd | 13.8 | 38 | 3.7 Sd |
| Eupeodes luniger (Meig.) | 2 | 2.5 Sd | 2.8 | 1 | 0.6 Sr | 2.8 | 9 | 5.7 D | 8.3 | 1 | 0.4 Sr | 2.8 | 13 | 1.3 R | ||||||
| Melangyna lasiophthalma (Zett.) | 8 | 4.9 Sd | 2.8 | 1 | 0.4 Sr | 2.8 | 9 | 0.9 Sr | ||||||||||||
| Melanostoma mellinum (L.) | 2 | 2.5 Sd | 5.6 | 6 | 3.7 Sd | 11.1 | 20 | 12.7 Eu | 16.7 | 3 | 1.3 R | 2 | 1.3 R | 5.6 | 2 | 0.8 Sr | 5.6 | 35 | 3.4 Sd | |
| Parasyrphus annulatus (Zett.) | 1 | 0.6 Sr | 2.8 | 30 | 13.0 Eu | 2.8 | 12 | 4.9 Sd | 5.6 | 43 | 4.2 Sd | |||||||||
| Platycheirus discimanus Loew | 1 | 0.4 Sr | 2.8 | 1 | 0.1 Sr | |||||||||||||||
| Platycheirus melanopsis Loew | 1 | 1.2 R | 2.8 | 1 | 0.6 Sr | 2.8 | 1 | 0.6 Sr | 2.8 | 1 | 0.4 Sr | 4 | 0.4 Sr | |||||||
| Platycheirus scutatus (Meig.) | 2 | 1.2 R | 5.6 | 3 | 1.9 R | 2.8 | 2 | 0.8 Sr | 5.6 | 7 | 0.7 Sr | |||||||||
| Scaeva pyrastri (L.) | 1 | 1.2 R | 2.8 | 2 | 1.3 R | 5.6 | 2 | 0.9 Sr | 5.6 | 2 | 0.8 Sr | 5.6 | 7 | 0.7 Sr | ||||||
| Scaeva selenitica (Meig.) | 1 | 1.2 R | 2.8 | 1 | 0.6 Sr | 2.8 | 2 | 0.2 Sr | ||||||||||||
| Sphaerophoria menthastri (L.) | 1 | 0.6 Sr | 2.8 | 1 | 0.4 Sr | 11.1 | 2 | 0.2 Sr | ||||||||||||
| Sphaerophoria scripta (L.) | 6 | 7.4 D | 5.6 | 15 | 9.2 D | 27.8 | 17 | 10.8 Eu | 22.2 | 4 | 1.7 R | 8.3 | 8 | 3.3 Sd | 13.9 | 50 | 4.9 Sd | |||
| Syrphus ribesii (L.) | 2 | 1.3 R | 5.6 | 10 | 4.3 Sd | 5.6 | 3 | 1.2 R | 2.8 | 15 | 1.5 R | |||||||||
| Syrphus torvus O.-S. | 1 | 1.2 R | 2.8 | 5 | 3.1 Sd | 11.1 | 6 | 3.8 Sd | 5.6 | 27 | 11.7 Eu | 5.6 | 4 | 2.6 Sd | 5.6 | 24 | 9.8 D | 16.7 | 67 | 6.5 D |
| Syrphus vitripennis Meig. | 2 | 2.5 Sd | 5.6 | 10 | 0.6 Sr | 16.7 | 7 | 4.4 Sd | 13.9 | 37 | 16.0 Eu | 8.3 | 27 | 17.8 Eu | 16.7 | 17 | 7.0 D | 16.7 | 100 | 9.7 D |
| Xanthandrus comtus (Harr.) | 1 | 1.2 R | 2.8 | 1 | 0.6 Sr | 2.8 | 2 | 0.2 Sr | ||||||||||||
| Xanthogramma pedissequum Harr. | 1 | 0.4 Sr | 2.8 | 1 | 0.1 Sr | |||||||||||||||
| Number of specimens | 81 | 163 | 158 | 231 | 152 | 244 | 1029 | |||||||||||||
| Number of species | 13 | 16 | 16 | 15 | 8 | 21 | ||||||||||||||
| a.c. 1 | a.c. 2 | a.c.3 | |||
|---|---|---|---|---|---|
| O 1 | 0.47 | O 2 | 0.41 | O 3 | 0.44 |
| S 1 | 0.39 | S 2 | 0.5 | S 3 | 0.24 |
| Habitat | Site 1 | Site 2 | Site 3 |
|---|---|---|---|
| Episyrphus balteatus (Deg.) | |||
| orchard | 14.67 ± 8.41 ab | 16.00 ± 6.00 ab | 9.33 ± 6.44 ab |
| surroundings | 37.67 ± 9.70 abc | 12.67 ± 4.67 ab | 41.67 ± 14.23 bc |
| aphid colonies | 8.00 ± 0.58 a | 5.00 ± 1.00 a | 6.33 ± 1.33 a |
| df = 11 F = 6.05 p = 0.000117 | |||
| Eupeodes corollae | |||
| orchard | 4.33 ± 2.85 ab | 9.67 ± 4.63 ab | 2.33 ± 1.33 ab |
| surroundings | 8.67 ± 6.67 ab | 6.67 ± 2.91 ab | 7.33 ± 1.45 ab |
| aphid colonies | 3.00 ± 0.58 ab | 2.00 ± 0.58 ab | 3.00 ± 0.58 ab |
| df = 11 F = 2.55 p = 0.026636 | |||
| Syrphus vitripennis | |||
| orchard | 1.0 ± 0.0 a | 2.33 ± 0.33 a | 2.33 ± 0.88 a |
| surroundings | 3.33 ± 2.33 a | 12.33 ± 8.95 a | 5.67 ± 3.28 a |
| aphid colonies | 2.00 ± 0.58 a | 1.67 ± 0.33 a | 1.00 ± 0.0 a |
| df = 11 F = 1.76 p = 0.119848 | |||
| Syrphid Specimens | ||||
|---|---|---|---|---|
| o/s | o/c | s/c | ||
| Site 1 | r2 p | 0.990596 0.087376 | 0.998353 0.036541 | 0.981116 0.123917 |
| Site 2 | r2 p | 0.356919 0.592379 | 0.568072 0.615379 | 0.320584 0.792242 |
| Site 3 | r2 p | 0.197369 0.706932 | 0.319044 0.617876 | 0.990232 0.089056 |
| Syrphid species | ||||
| Site1 | r2 p | 0.076923 0.821088 | 0.923077 0.178912 | 0.0 1.0 |
| Site 2 | r2 p | 0.428571 0.545629 | 0.964286 0.121038 | 0.617347 0.424591 |
| Site 3 | r2 p | 0.013158 0.926814 | 1.000000 | 0.013158 0.926814 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wojciechowicz-Żytko, E.; Wilk, E. Surrounding Semi-Natural Vegetation as a Source of Aphidophagous Syrphids (Diptera, Syrphidae) for Aphid Control in Apple Orchards. Agriculture 2023, 13, 1040. https://doi.org/10.3390/agriculture13051040
Wojciechowicz-Żytko E, Wilk E. Surrounding Semi-Natural Vegetation as a Source of Aphidophagous Syrphids (Diptera, Syrphidae) for Aphid Control in Apple Orchards. Agriculture. 2023; 13(5):1040. https://doi.org/10.3390/agriculture13051040
Chicago/Turabian StyleWojciechowicz-Żytko, Elżbieta, and Edyta Wilk. 2023. "Surrounding Semi-Natural Vegetation as a Source of Aphidophagous Syrphids (Diptera, Syrphidae) for Aphid Control in Apple Orchards" Agriculture 13, no. 5: 1040. https://doi.org/10.3390/agriculture13051040
APA StyleWojciechowicz-Żytko, E., & Wilk, E. (2023). Surrounding Semi-Natural Vegetation as a Source of Aphidophagous Syrphids (Diptera, Syrphidae) for Aphid Control in Apple Orchards. Agriculture, 13(5), 1040. https://doi.org/10.3390/agriculture13051040