
Evaluation of Introgressed Lines of Sunflower (Helianthus annuus L.) under Contrasting Water Treatments

 , ,
, ,  and
and
Abstract
:1. Introduction
2. Materials and Methods
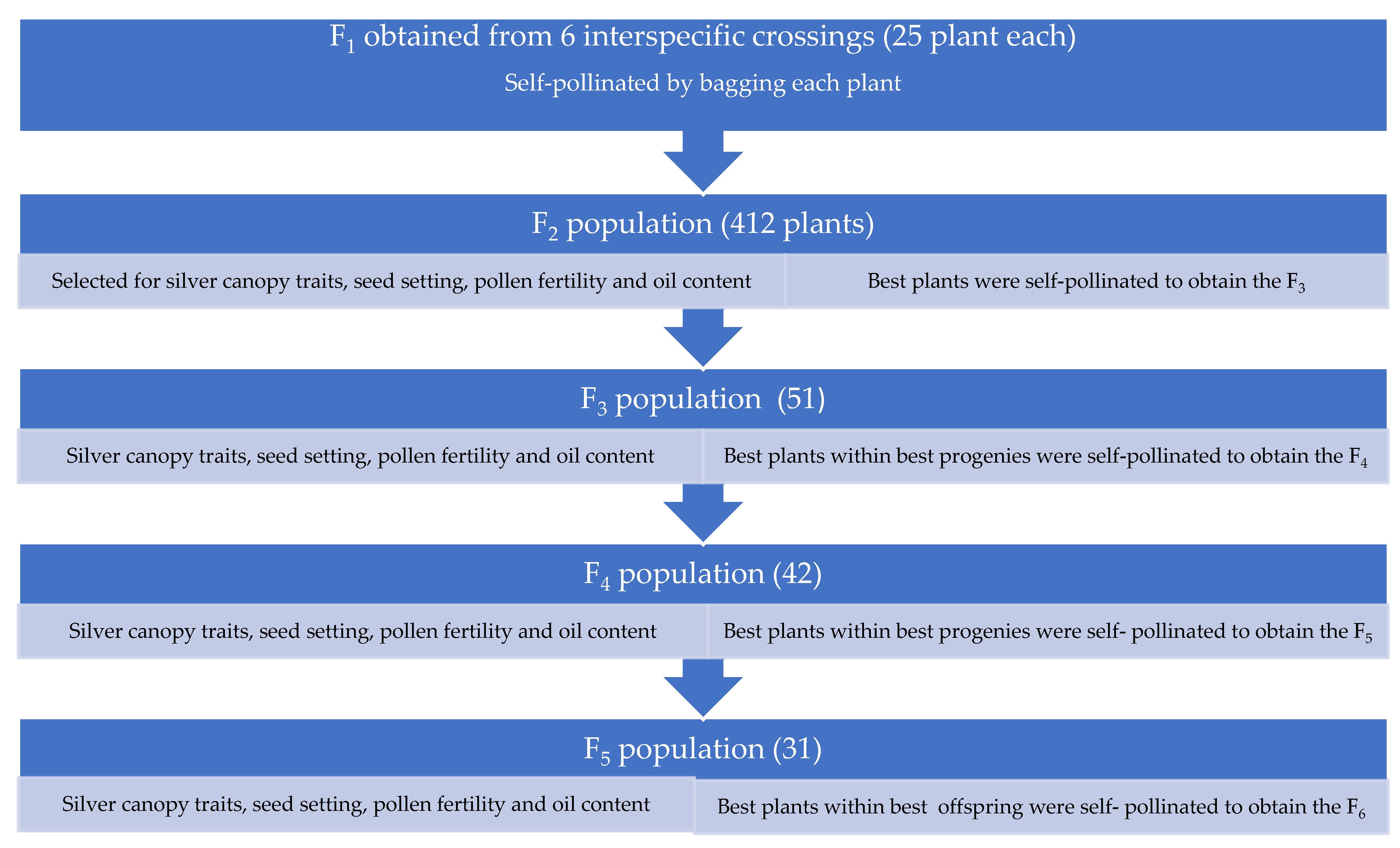
2.1. Development of Drought-Tolerant Breeding Lines
2.2. Evaluation of Drought-Tolerant Plant Selections under Contrasting Water Treatments in Polythene Bags (Figure 3)
- Leaf area: It was determined using a CID Bio-Science CI-202 portable leaf area meter. Leaf area of three leaves from each plant was used to determine the average leaf area plant−1. There were five plants within each replication.
- Root length (cm): Polythene bags were gently dissected and roots were washed with constant gentle water pressure to avoid breakage of roots. The roots were dried to remove any surface moisture and the primary root length was measured.
- Shoot length (cm): It was determined from the base to the tip of the meristematic tissues.
- Fresh root and shoot biomass: The above ground shoot biomass and root biomass of five plants within each replication were recorded.
- Root shoot ratio: The root biomass was divided by shoot biomass to estimate the root shoot ratio.
- Drought resistance index (DRI): It was calculated using the following formula:
- 7.
- Cuticular wax: It was determined following work by Hussain et al. [10]. Leaf disc of 8 cm2 was obtained from 15-day-old leaf from top of canopy. Adaxial leaf discs were dipped in chloroform for 15 s at 25 °C. Extract was filtered and chloroform was evaporated. About 5 mL of reagent was prepared using 20 g of potassium dichromate dissolved in 40 mL of distilled water. The reagent solution was then mixed in concentrated H2SO4 for 1 h. Thereafter, 1 mL of reagent was added to develop chrome. Readings were obtained at optical density of 590 nm and noted. A standard solution was prepared by mixing known concentration of cuticular wax in the solution ranging from 0 to 100 µg L−1.
2.3. Combining Ability Test
2.4. Statistical Analysis
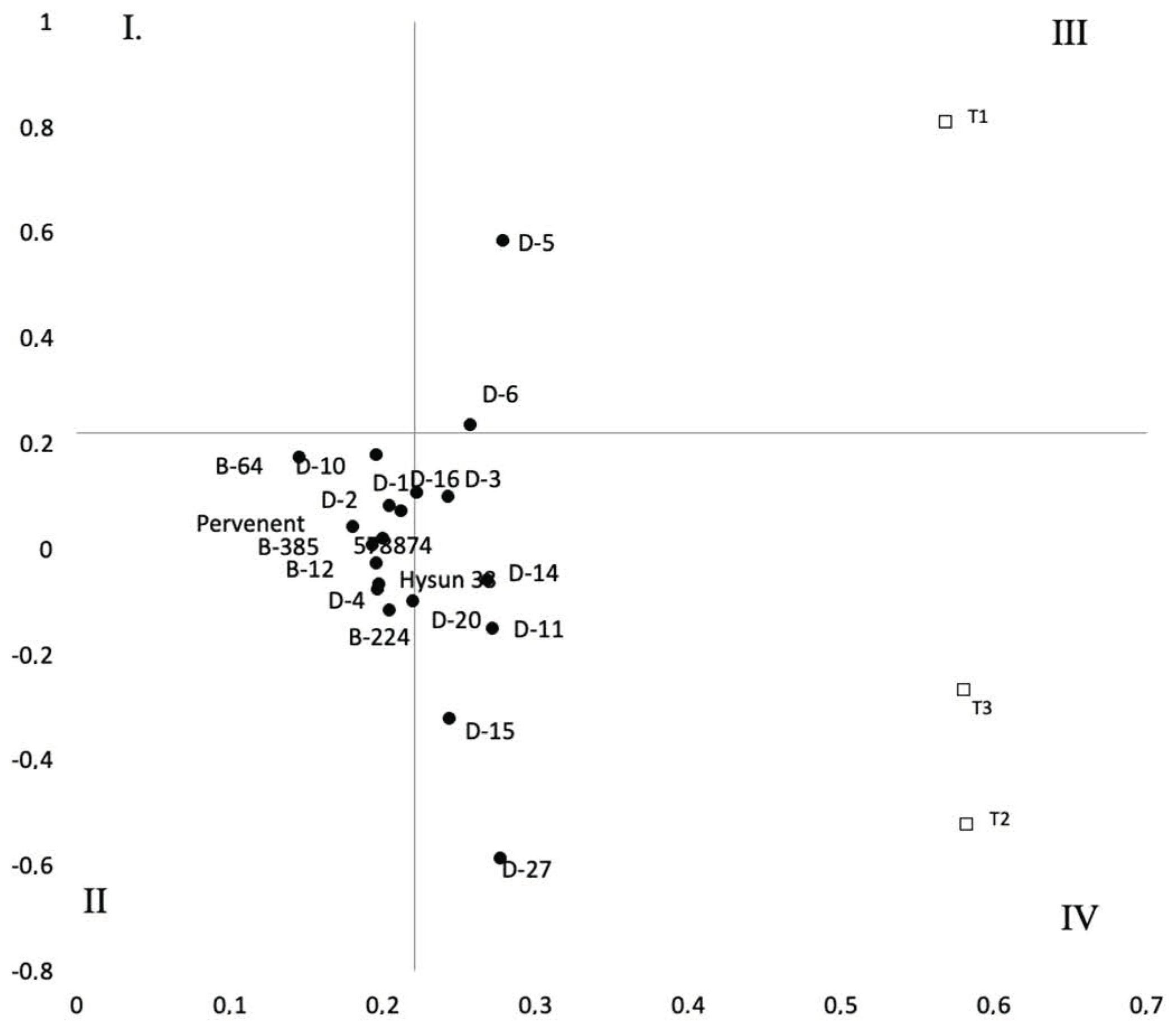
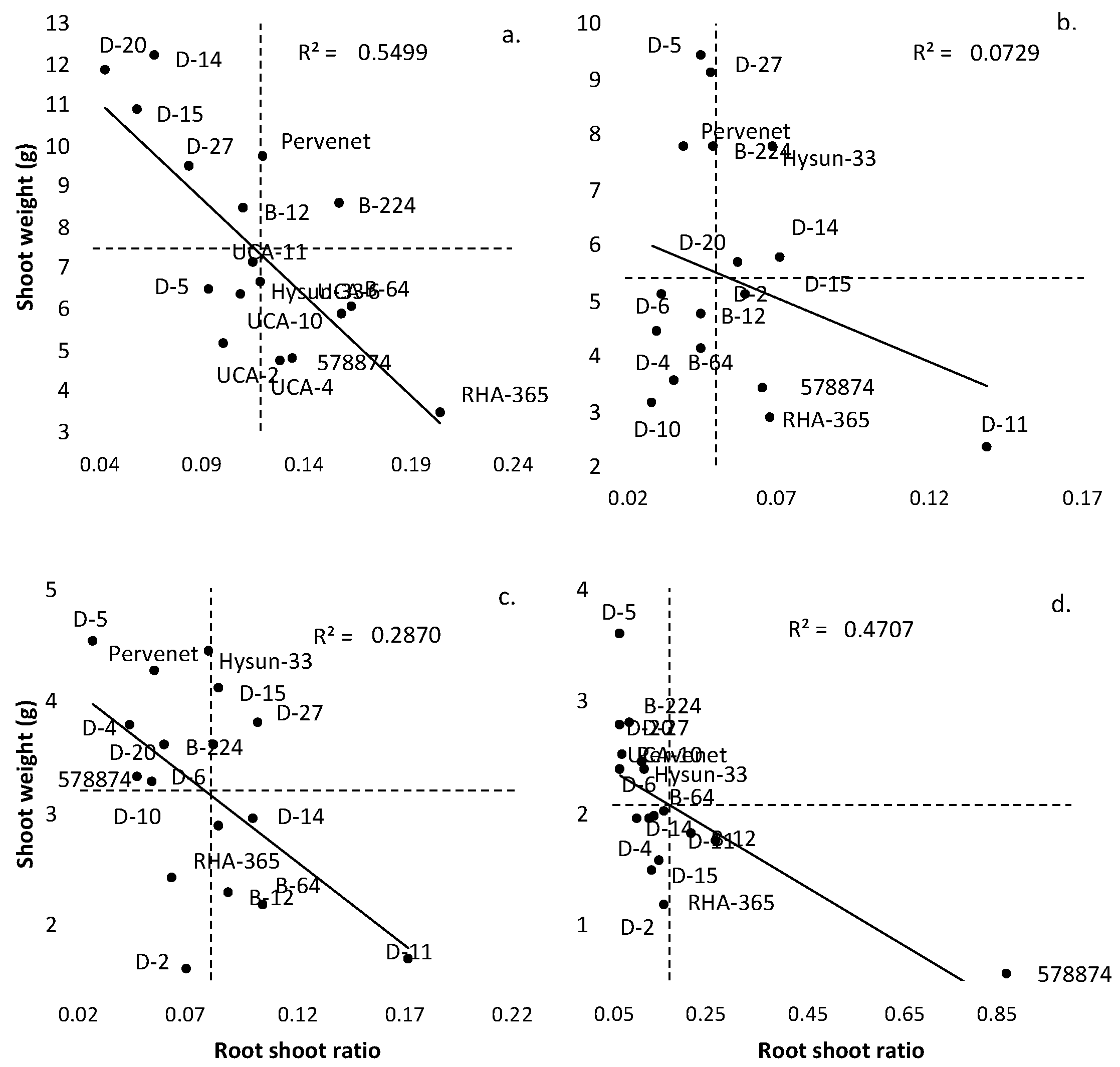
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Rauf, S.; Al-Khayri, J.M.; Zaharieva, M.; Monneveux, P.; Khalil, F. Breeding strategies to enhance drought tolerance in crops. In Advances in Plant Breeding Strategies: Agronomic, Abiotic and Biotic Stress Traits; Al-Khayri, J.M., Jain, S.M., Johnson, D.V., Eds.; Springer: Cham, Switzerland; New York, NY, USA, 2016; pp. 397–445. [Google Scholar]
- Rauf, S.; Sadaqat, H.A. Effect of osmotic adjustment on root length and dry matter partitioning in sunflower (Helianthus annuus L.) under drought stress. Acta Agric. Scand. B Soil Plant Sci. 2008, 58, 252–260. [Google Scholar]
- Food and Agriculture Organization (FAO). FAOSTATs; FAO: Rome, Italy, 2016; Available online: http://www.fao.org/faostat/en/#home (accessed on 16 July 2020).
- Rauf, S. Breeding sunflower (Helianthus annuus L.) for drought tolerance. Commun. Biometry Crop Sci. 2008, 3, 29–44. [Google Scholar]
- Ebrahimian, E.; Seyyedi, S.M.; Bybordi, A.; Damalas, C.A. Seed yield and oil quality of sunflower, safflower, and sesame under different levels of irrigation water availability. Agric. Water Manag. 2019, 218, 149–157. [Google Scholar] [CrossRef]
- Zamani, S.; Naderi, M.R.; Soleymani, A.; Nasiri, B.M. Sunflower (Helianthus annuus L.) biochemical properties and seed components affected by potassium fertilization under drought conditions. Ecotoxicol. Environ. Saf. 2020, 190, 110017. [Google Scholar] [CrossRef]
- Khalil, F.; Rauf, S.; Monneveux, P.; Anwar, S.; Iqbal, Z. Genetic analysis of proline concentration under osmotic stress in sunflower (Helianthus annuus L.). Breed. Sci. J. 2016, 66, 463–470. [Google Scholar] [CrossRef] [Green Version]
- Warburton, M.L.; Rauf, S.; Marek, L.; Hussain, M.; Ogunola, O. The use of crop wild relatives in maize and sunflower breeding. Crop Sci. 2017, 57, 1227–1440. [Google Scholar] [CrossRef]
- Rauf, S. Breeding strategies for sunflower (Helianthus annuus L.) genetic improvement. In Advances in Plant Breeding Strategies: Industrial and Food Crops; Al-Khayri, J.M., Jain, S.M., Johnson, D.V., Eds.; Springer: Cham, Switzerland; New York, NY, USA, 2019; pp. 637–673. [Google Scholar]
- Hussain, M.M.; Rauf, S.; Riaz, M.A.; Al-Khayri, J.M.; Monneveux, P. Determination of drought tolerance related traits in Helianthus argophyllus, Helianthus annuus, and their hybrids. Breed. Sci. J. 2017, 67, 257–267. [Google Scholar] [CrossRef] [Green Version]
- Shehbaz, M.; Rauf, S.; Al-Sadi, A.M.; Nazir, S.; Bano, S. Introgression and inheritance of charcoal rot (Macrophomina phaseolina) resistance from silver sunflower (Helianthus argophyllus Torr. & A. Gray) into cultivated sunflower (Helianthus annuus L.). Australas. Plant Pathol. 2018, 47, 413–420. [Google Scholar]
- Hussain, M.M.; Rauf, S.; Paderewski, J.; ul Haq, I.; Sienkiewics-Paderwska, D.; Monneveux. Multitraits evaluation of Pakistani ecotypes of berseem clover (Trifolium alexandrinum L.) under full irrigation and water restriction conditions. J. Appl. Bot. Food Qual. 2015, 88, 127–133. [Google Scholar]
- Rauf, S.; Warburton, M.; Naeem, A.; Kainat, W. Validated markers for sunflower (Helianthus annuus L.) breeding. OCL 2020, 27, 47. [Google Scholar] [CrossRef]
- Wu, Y.; Shi, H.; Yu, H.; Ma, Y.; Hu, H.; Han, Z.; Hou, J. Combined GWAS and transcriptome analyses provide new insights into the response mechanisms of sunflower against drought stress. Front. Plant Sci. 2022, 13, 847435. [Google Scholar] [CrossRef] [PubMed]
- Makhtoum, S.; Sabouri, H.; Gholizadeh, A.; Ahangar, L.; Katouzi, M.; Mastinu, A. Mapping of QTLs controlling barley agronomic traits (Hordeum vulgare L.) under normal conditions and drought and salinity stress at reproductive stage. Plant Gene 2022, 31, 100375. [Google Scholar] [CrossRef]
- Sajid, H.B.; Luqman, M.; Faisal, M.A.; Zohra, T. Accelerated breeding for Helianthus annuus (sunflower) through genome editing. Int. J. Res. Adv. Agric. Sci. 2022, 1, 13–27. [Google Scholar]
- Reynolds, S.G. The gravimetric method of soil moisture determination Part IA study of equipment, and methodological problems. J. Hydrol. 1970, 11, 258–273. [Google Scholar] [CrossRef]
- Acquaah, G. Principles of Plant Genetics and Breeding; Blackwell Pub. Ltd.: Oxford, UK, 2007. [Google Scholar]
- Kempthorne, O. An Introduction to Genetic Statistics; John Wiley and Sons Inc.: New York, NY, USA, 1957. [Google Scholar]
- Rengel, D.; Arribat, S.; Maury, P.; Martin-Magniette, M.L.; Hourlier. A gene-phenotype network based on genetic variability for drought responses reveals key physiological processes in controlled and natural environments. PLoS ONE 2012, 7, e45249. [Google Scholar] [CrossRef]
- El–Bially, M.E.; Saudy, H.S.; Hashem, F.A.; El–Gabry, Y.A.; Shahin, M.G. Salicylic acid as a tolerance inducer of drought stress on sunflower grown in sandy soil. Gesunde Pflanz. 2022, 74, 603–613. [Google Scholar] [CrossRef]
- Kosar, F.; Alshallash, K.S.; Akram, N.A.; Sadiq, M.; Ashraf, M.; Alkhalifah, D.H.M.; Elkelish, A. trehalose-induced regulations in nutrient status and secondary metabolites of drought-stressed sunflower (Helianthus annuus L.) plants. Plants 2022, 11, 2780. [Google Scholar] [CrossRef]
- Vancostenoble, B.; Blanchet, N.; Langlade, N.B.; Bailly, C. Maternal drought stress induces abiotic stress tolerance to the progeny at the germination stage in sunflower. Environ. Exp. Bot. 2022, 201, 104939. [Google Scholar] [CrossRef]
- Jan, A.U.; Hadi, F.; Ditta, A.; Suleman, M.; Ullah, M. Zinc-induced anti-oxidative defense and osmotic adjustments to enhance drought stress tolerance in sunflower (Helianthus annuus L.). Environ. Exp. Bot. 2022, 193, 104682. [Google Scholar] [CrossRef]
- Hussain, M.M.; Kausar, M.; Rauf, S.; Khan, M.F.Z.; Paderweski, J. Selection for some functional markers for adaptability of Helianthus argophyllus × Helianthus annuus derived population under abiotic stress conditions. Helia 2018, 41, 83–108. [Google Scholar] [CrossRef]
- Safdar, T.; Tahir, M.H.N.; Ali, Z.; ur Rahman, M.H. Exploring the role of HaTIPs genes in enhancing drought tolerance in sunflower. Res. Sq. 2023. [Google Scholar] [CrossRef]
- Isbell, F.; Adler, P.R.; Eisenhauer, N.; Fornara, D.; Kimmel, K. Benefits of increasing plant diversity in sustainable agroecosystems. J. Ecol. 2017, 105, 871–879. [Google Scholar] [CrossRef] [Green Version]
- Henn, J.J.; Buzzard, V.; Enquist, B.J.; Halbritter, A.H.; Klanderud, K. Intraspecific trait variation and phenotypic plasticity mediate alpine plant species response to climate change. Front. Plant Sci. 2018, 9, 1548. [Google Scholar] [CrossRef]
- Fasahat, P.; Rajabi, A.; Rad, J.M.; Derera, J. Principles and utilization of combining ability in plant breeding. Biom. Biostat. Intl. J. 2016, 4, 1–24. [Google Scholar] [CrossRef]
- Moschen, S.; Bengoa Luoni, S.; Di Rienzo, J.A.; Caro, M.D.P.; Tohge, T.; Watanabe, M.; Hollmann, J.; González, S.; Rivarola, M.; García-García, F.; et al. Integrating transcriptomic and metabolomic analysis to understand natural leaf senescence in sunflower. Plant Biotechnol. J. 2016, 14, 719–734. [Google Scholar] [CrossRef] [Green Version]
- Gody, L.; Duruflé, H.; Blanchet, N.; Carré, C.; Legrand, L.; Mayjonade, B. Transcriptomic data of leaves from eight sunflower lines and their sixteen hybrids under water deficit. OCL 2020, 27, 48. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Breeding Material | Status | Oil Content (%) | Days to Flowering | Canopy Color | Cuticular Wax (µg g−1) |
|---|---|---|---|---|---|
| 578874 | OPV * | 34.12 | 52.46 | Green | 3.19 |
| B.224 | B-Line ** | 33.38 | 50.58 | Green | 3.3 |
| B.64 | B-Line | 34.12 | 50.13 | Green | 2.8 |
| Pervenent | OPV | 35.24 | 53.39 | Green | 2.14 |
| B.385 | B-Line | 40.15 | 56.34 | 3.65 | |
| B.12 | OPV | 38.63 | 52.37 | Green | 2.13 |
| UCA-1-DR | F5 Progeny *** | 37.83 | 68.36 | Silver | 12.13 |
| UCA-2-DR | F5 Progeny | 31.29 | 64.37 | Silver | 9.89 |
| UCA-3-DR | F5 Progeny | 35.13 | 70.35 | Silver | 13.14 |
| UCA-4-DR | F5 Progeny | 38.17 | 72.34 | Silver | 12.16 |
| UCA-5-DR | F5 Progeny | 38.24 | 70.31 | Silver | 10.24 |
| UCA-6-DR | F5 Progeny | 36.58 | 62.36 | Silver | 17.19 |
| UCA-10-DR | F5 Progeny | 36.13 | 69.34 | Silver | 12.34 |
| UCA-11-DR | F5 Progeny | 35.19 | 70.32 | Silver | 10.09 |
| UCA-14-DR | F5 Progeny | 28.35 | 62.32 | Silver | 11.12 |
| UCA-15-DR | F5 Progeny | 38.91 | 71.38 | Silver | 10.16 |
| UCA-16-DR | F5 Progeny | 36.19 | 73.27 | Silver | 13.12 |
| UCA-20-DR | F5 Progeny | 35.15 | 61.29 | Silver | 8.72 |
| UCA-27-DR | F5 Progeny | 35.23 | 70.35 | Silver | 9.12 |
| Hysun 33 | 34.56 | 74.35 | Green | 10.13 | |
| FH 331 | Hybrid | 35.38 | 70.18 | Green | 9.34 |
| Source of Variation | Degrees of Freedom | Mean Sum of Squares | |||||
|---|---|---|---|---|---|---|---|
| LA | RL | SL | SW | RW | R/S | ||
| Lines | 19 | 14.81 ** | 10.54 ** | 608.08 ** | 18.67 ** | 0.07 ** | 0.03 ** |
| Water regimes (DL) | 3 | 489.57 ** | 10.14 ** | 2235.11 ** | 251.43 ** | 3.03 ** | 0.14 ** |
| Lines × DL interaction | 57 | 24.00 ** | 4.77 ** | 99.13 ** | 6.82 ** | 0.05 ** | 0.03 ** |
| Residual | 160 | 5.41 | 1.12 | 36.47 | 1.27 | 0.01 | 0.00 |
| σ2 Genotype | 0.00 | 0.64 | 56.55 | 1.32 | 0.00 | 0.01 | |
| σ2 Environment | 5.41 | 1.12 | 36.47 | 1.27 | 0.01 | 0.01 | |
| σ2 Phenotype | 10.59 | 2.98 | 113.91 | 4.44 | 0.03 | 0.03 | |
| Broad-sense heritability | 0.00 | 0.22 | 0.50 | 0.30 | 0.10 | 0.58 | |
| Breeding Lines | Leaf Area (cm2) | Root Length (cm) | ||||||
|---|---|---|---|---|---|---|---|---|
| Control | T1 | T2 | T3 | Control | T1 | T2 | T3 | |
| Tolerant inbred lines | 22.63b | 22.98a | 20.60ab | 17.72a | 6.12b | 6.53ab | 6.61a | 7.19b |
| Phenotypic coefficient of variation (%) | 9.04 | 11.11 | 12.90 | 15.82 | 13.38 | 20.82 | 21.57 | 20.85 |
| Range | 19.13–26.21 | 19.97–28.57 | 17.16–25.33 | 13.50–23.18 | 5.00–7.37 | 3.87–8.70 | 3.60–8.67 | 4.50–9.89 |
| Elite germplasm | 28.71a | 22.76a | 19.10b | 17.22a | 6.10b | 5.45a | 6.63a | 6.55b |
| Phenotypic coefficient of variation (%) | 10.16 | 15.92 | 12.50 | 23.37 | 27.50 | 19.48 | 32.36 | 23.37 |
| Range | 23.12–29.56 | 17.67–27.80 | 15.04–21.29 | 15.31–19.36 | 4.30–8.10 | 2.83–7.73 | 4.67–7.90 | 3.67–9.40 |
| Hybrids | 29.12a | 24.95a | 23.85a | 15.58a | 7.83a | 8.07a | 6.93a | 9.37a |
| Breeding Lines | Shoot Weight (g) | Root Weight (g) | ||||||
|---|---|---|---|---|---|---|---|---|
| Control | T1 | T2 | T3 | Control | T1 | T2 | T3 | |
| Tolerant inbred lines | 7.49a | 5.66b | 3.61a | 2.09a | 0.63b | 0.29b | 0.25a | 0.24a |
| Phenotypic coefficient of variation (%) | 33.34 | 40.41 | 26.93 | 30.88 | 18.91 | 44.74 | 40.19 | 34.69 |
| Range | 4.23–11.67 | 2.57–9.63 | 2.00–4.93 | 1.37–3.50 | 0.47–0.80 | 0.10–0.56 | 0.13–0.43 | 0.13–0.42 |
| Elite germplasm | 6.31a | 5.26b | 3.40a | 1.67a | 0.83a | 0.26b | 0.21a | 0.23a |
| Phenotypic coefficient of variation (%) | 39.44 | 38.61 | 25.12 | 48.64 | 34.88 | 30.94 | 18.99 | 29.74 |
| Range | 2.93–9.23 | 3.13–8.00 | 2.57–4.67 | 0.45–2.70 | 0.47–1.27 | 0.13–0.36 | 0.16–0.25 | 0.14–0.42 |
| Hybrids | 6.13a | 8.00a | 4.83a | 2.33a | 0.58b | 0.48a | 0.27a | 0.29a |
| Breeding Lines | Shoot Length (cm) | Root to Shoot Ratio | ||||||
|---|---|---|---|---|---|---|---|---|
| Control | T1 | T2 | T3 | Control | T1 | T2 | T3 | |
| Tolerant inbred lines | 50.35a | 45.55b | 39.97b | 36.22ab | 0.15a | 0.05a | 0.06a | 0.30a |
| Phenotypic coefficient of variation (%) | 17.67 | 21.25 | 9.83 | 10.29 | 23.21 | 60.48 | 69.98 | 78.56 |
| Range | 35.73–65.62 | 29.17–59.17 | 35–49.17 | 27.92–41.67 | 0.05–0.16 | 0.03–0.14 | 0.03–0.17 | 0.07–0.26 |
| Elite germplasm | 48.83a | 40.29b | 35.56b | 32.13b | 0.10a | 0.06a | 0.07a | 0.12b |
| Phenotypic coefficient of variation (%) | 25.27 | 22.16 | 30.47 | 35.30 | 35.45 | 22.08 | 30.63 | 89.71 |
| Range | 26.33–60.28 | 31.83–58.75 | 15.00–47.08 | 9.58–46.67 | 0.11–0.21 | 0.04–0.07 | 0.05–0.10 | 0.07–0.26 |
| Hybrids | 51.78a | 56.25a | 52.83a | 45.21a | 0.12a | 0.07a | 0.08a | 0.10b |
| Breeding Lines | Sargodha | GCA * | Faisalabad | GCA | ||
|---|---|---|---|---|---|---|
| R.365 | R.SIN.82 | R.365 | R.SIN.82 | |||
| Head Diameter (cm) | ||||||
| D2 | 19.33 ± 1.53 | 22.33 ± 0.58 | −1.95 | 18.33 ± 2.08 | 21.0 ± 1.21 | −1.21 |
| D5 | 17.67 ± 2.08 | 22.00 ± 2.65 | −2.95 | 14.00 ± 1.35 | 22.0 ± 2.65 | −2.88 |
| D20 | 23.00 ± 1.13 | 26.00 ± 1.39 | 1.71 | 19.67 ± 1.53 | 21.7 ± 1.53 | −0.21 |
| D26 | 23.33 ± 2.11 | 23.67 ± 0.58 | 0.71 | 18.33 ± 2.10 | 21.3 ± 1.61 | −1.05 |
| D27 | 23.00 ± 1.00 | 26.00 ± 1.32 | 1.71 | 19.00 ± 1.50 | 22.0 ± 1.52 | −0.38 |
| B.224 | 22.67 ± 1.53 | 22.67 ± 1.53 | −0.12 | 22.00 ± 1.25 | 23.7 ± 0.58 | 1.95 |
| 577874 | 24.00 ± 1.39 | 23.33 ± 1.69 | 0.88 | 24.00 ± 1.35 | 25.3 ± 1.19 | 3.79 |
| Oil content (%) | ||||||
| D2 | 33.00 ± 2.00 | 34.33 ± 2.08 | −1.95 | 35.00 ± 1.39 | 35.7 ± 0.58 | −0.36 |
| D5 | 36.67 ± 1.53 | 37.33 ± 1.53 | 1.38 | 35.67 ± 1.53 | 36.3 ± 0.67 | 0.31 |
| D20 | 36.00 ± 1.57 | 36.67 ± 1.69 | 0.71 | 36.67 ± 1.61 | 35.0 ± 1.35 | 0.14 |
| D26 | 35.67 ± 2.08 | 37.67 ± 1.53 | 1.05 | 36.67 ± 2.15 | 35.7 ± 0.89 | 0.48 |
| D27 | 35.67 ± 1.65 | 37.00 ± 1.21 | 0.71 | 36.00 ± 2.03 | 36.3 ± 0.63 | 0.48 |
| B.224 | 34.33 ± 0.58 | 33.67 ± 1.53 | −1.62 | 35.33 ± 0.58 | 35.0 ± 1.21 | −0.52 |
| 577874 | 35.67 ± 1.08 | 35.00 ± 1.19 | −0.29 | 35.00 ± 1.35 | 35.3 ± 0.61 | −0.52 |
| Seed yield (g plant−1) | ||||||
| D2 | 42.22 ± 3.61 | 54.21 ± 2.80 | −8.71 | 46.97 ± 1.20 | 55.9 ± 1.38 | −8.30 |
| D5 | 47.40 ± 1.82 | 49.63 ± 2.93 | −8.41 | 55.30 ± 1.94 | 60.3 ± 0.97 | −1.93 |
| D20 | 54.97 ± 1.56 | 56.67 ± 1.49 | −1.11 | 60.00 ± 0.58 | 62.4 ± 1.02 | 1.45 |
| D26 | 65.96 ± 3.46 | 63.95 ± 2.03 | 8.03 | 66.49 ± 4.05 | 68.7 ± 1.76 | 7.88 |
| D27 | 62.67 ± 3.53 | 65.33 ± 3.64 | 7.07 | 62.89 ± 2.62 | 58.0 ± 1.57 | 0.71 |
| B.224 | 51.92 ± 1.47 | 59.31 ± 0.94 | −1.31 | 56.31 ± 2.00 | 58.6 ± 1.95 | −2.29 |
| 577874 | 58.61 ± 3.69 | 64.10 ± 1.56 | 4.43 | 62.17 ± 2.70 | 62.3 ± 2.13 | 2.48 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hussain, M.M.; Rauf, S.; Noor, M.; Bibi, A.; Ortiz, R.; Dahlberg, J. Evaluation of Introgressed Lines of Sunflower (Helianthus annuus L.) under Contrasting Water Treatments. Agriculture 2023, 13, 1250. https://doi.org/10.3390/agriculture13061250
Hussain MM, Rauf S, Noor M, Bibi A, Ortiz R, Dahlberg J. Evaluation of Introgressed Lines of Sunflower (Helianthus annuus L.) under Contrasting Water Treatments. Agriculture. 2023; 13(6):1250. https://doi.org/10.3390/agriculture13061250
Chicago/Turabian StyleHussain, Muhammad Mubashar, Saeed Rauf, Muqadas Noor, Amir Bibi, Rodomiro Ortiz, and Jeff Dahlberg. 2023. "Evaluation of Introgressed Lines of Sunflower (Helianthus annuus L.) under Contrasting Water Treatments" Agriculture 13, no. 6: 1250. https://doi.org/10.3390/agriculture13061250
APA StyleHussain, M. M., Rauf, S., Noor, M., Bibi, A., Ortiz, R., & Dahlberg, J. (2023). Evaluation of Introgressed Lines of Sunflower (Helianthus annuus L.) under Contrasting Water Treatments. Agriculture, 13(6), 1250. https://doi.org/10.3390/agriculture13061250