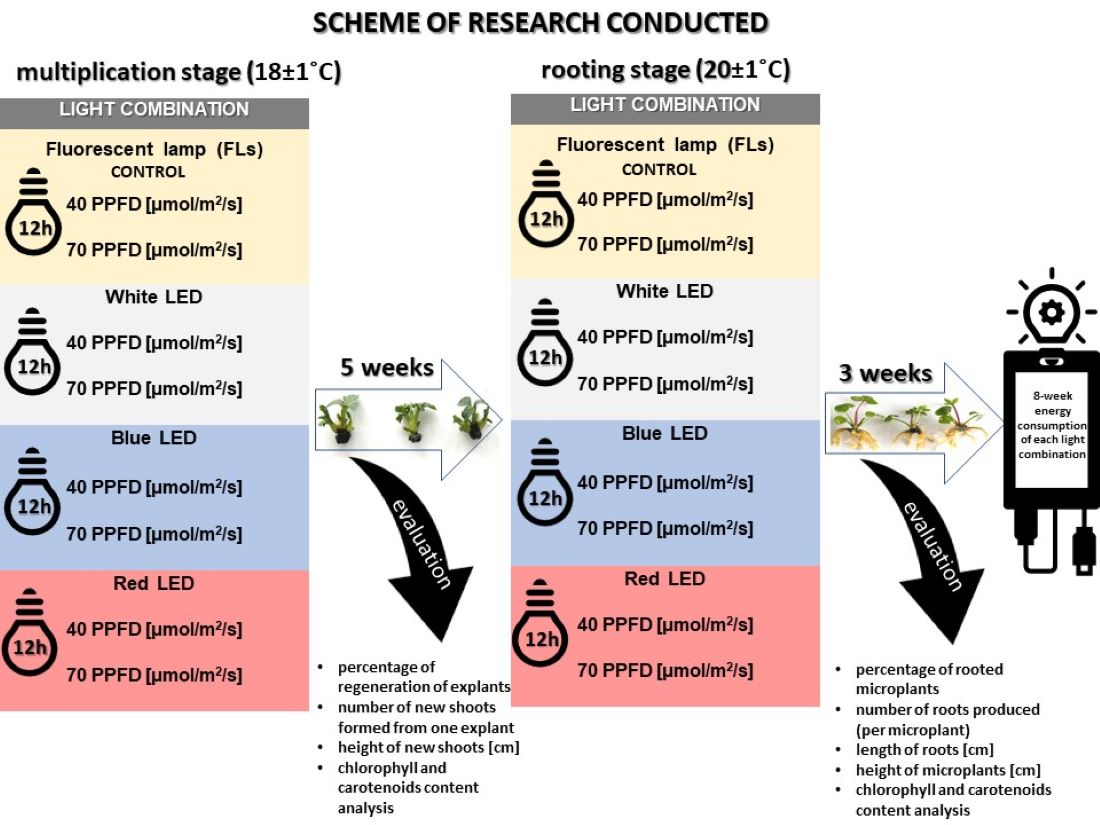
1. Introduction
Hellebores have remained of interest to humans for years. They belong to the
Ranunculaceae family, and their natural habitat is Europe and Asia. The majority of
Helleborus species are concentrated in the Balkan region. Poland is inhabited by
H. purpurescens, which is under legal protection due to the threat of extinction [
1]. In addition to its decorative value, hellebore is also a medicinal plant. In its composition, it includes heleborin, which exhibits narcotic and hemolytic effects, and heleborein, which exhibits cardiac effects similar to digitalis. Depending on dosage and treatment, the extract, as well as individual fractions of hellebore rhizomes and roots, exhibits multiple bioactive effects and can act in several directions. In most cases, the mechanisms of action are not sufficiently studied and described or are unknown [
2].
As has been repeatedly shown, micropropagation is currently the most modern and promising method of vegetative propagation of both ornamental [
3,
4] and endangered plant species [
5,
6,
7]. Due to the different requirements of individual species, and even their varieties, as well as the problems that arise at different stages of conducting in vitro cultures, protocols should be developed for each specific plant. It is also necessary to keep looking for new solutions to make the method as cost-effective as possible and the regenerating plants of the best possible quality [
8]. Many factors affect plant regeneration under sterile, practically fully controlled in vitro conditions. Among the most important are the type of explant; the composition of the medium, including the concentration and type of plant growth regulators; and of course, light [
9,
10]. Light, including its source and quality, is a fundamental factor in the proper growth and development of plants [
11,
12]. Plants use light in a number of vital processes, including the primary one of photosynthesis, which affects their nutrition and appearance, and hence the quality of the material. Depending on the parameters of the light available during growth, plants can respond in completely different ways. With excessively strong lighting, overly intense, elongated growth is usually observed. On the other hand, in response to too little light, growth is often reduced. This ability of plants and chloroplasts to adapt to light is the main basic form of photomorphogenetic response, which is associated with specific changes in plant morphology, physiology and biochemistry [
13,
14].
Due to the significant impact of light on plant development, people began to improve its source and quality in horticultural production, wanting to obtain high-quality propagation material. Lamp producers began to market products tailored specifically to the needs of plants. Fluorescent lamps (FLs) are the traditional light source used in phytotrons for micropropagation. Nevertheless, their disadvantages, such as high power consumption and the production of a wide range of wavelengths (350–750 nm) unnecessary in plant development, have been reported for years [
15]. Light-emitting diodes (LEDs) have emerged as an alternative and were quickly accepted positively in commercial production. LEDs have many advantages over fluorescent lamps—lower heat emission, monochromatic spectrum, longer life and low energy consumption—which contributes to their pro-environmental nature. LED lighting systems for in vitro cultures make it possible to control this important parameter and provide light in the spectral range that is used in photosynthesis and positively affects photomorphogenic reactions in plants [
16,
17]. The colors or combinations of LEDs commonly used in in vitro cultures are white (W), red (R), blue (B) and a mix of blue and red (B+R). It has been reported that red light is important for elongation growth of shoots and stems, phytochrome reactions and changes in plant anatomy [
18], while blue light is important in chlorophyll biosynthesis, opening of stomata, chloroplast maturation and photosynthesis. Together, blue and red have been used in different combinations of LEDs in many studies on photosynthesis and chlorophyll synthesis [
19]. In experiments conducted for 8 weeks under LED light (white, red, blue, R+B combination) on three different ornamental plant species,
Cordyline australis,
Ficus benjamina and
Sinningia speciosa, authors showed that blue and R+B resulted in higher maximum quantum yield in all species and higher plant biomass in this light combination [
20]. During micropropagation of
Zantedeschia, it was found that blue light affected plant height and chlorophyll [
21]. LED light affected potted chrysanthemum production differently. Plants under R+B light had the highest leaf greenness index (SPAD) value and the shortest cuttings with the longest roots, while the W+B LED combination significantly affected most growth parameters, with the exception of plant height and number of leaves [
22].
Therefore, the response to LED lighting depends on the wavelength to which plants are exposed and varies from species to species [
23]. Therefore, there is a need for research to learn more about the response of important species, such as hellebores, to LED lighting, which will replace fluorescent light sources entirely in the near future. We hypothesized that LED light would produce comparable or even better effects on hellebore micropropagation than traditional fluorescent lighting (FLs) at a significant reduction in production costs. Thus, the aim of the study was to increase the micropropagation efficiency of the hellebore ‘Molly’s White’ using energy-efficient light-emitting diodes (LEDs) in white, blue and red, which were compared with traditionally used fluorescent lights (FLs).
4. Discussion
Light is one of the most important external factors affecting all plant development, due to its regulatory role in photosynthetic, biochemical and molecular processes. Choosing the optimal light intensity to support plant proliferation and growth in vitro is particularly important. Explants in vitro are exposed to much lower light intensities compared to plants grown in the field. Artificial, poor and often poor-quality lighting in in vitro cultures has been recognized as a limiting factor for photosynthesis, so sucrose is added to the medium as a standard carbohydrate source. In vitro plants are also very sensitive to high light conditions and susceptible to photoinhibition [
27,
28,
29].
In the presented study, it was shown that during micropropagation of hellebore, better rates of most of the parameters studied were obtained with more intense light. Similar results, especially when increasing the intensity to 94 μmol·m
−2·s
−1, were previously obtained by Lazzarini et al. [
30] for
Lippia gracilis, Eun-A et al. [
31] at 30 μmol/m
2/s for
Alocasia amazonica and Chen et al. [
32] for
Haworthia. Light intensity and quality are the most critical environmental factors for plant physiology and biochemistry. According to available research, many basic parameters of plant production, including dry weight of roots, leaves and the whole plant, as well as the rate of photosynthesis, transpiration and stomatal conductance, were reduced under conditions of insufficient light [
33,
34].
In addition to increased plant height during multiplication, higher intensity (70 μmol/m
2/s) light increased the content of assimilation pigments in regenerating and rooting hellebore plants. Lazzarini et al. [
30] reported that an increase in carotenoid synthesis was observed in plants grown in high light intensities and was related to the photoprotection exerted by these pigments within photosystems. Carotenoids are synthesized in the plastids of all photosynthesizing organisms, hence their important function in photosynthesis [
35]; however, it should not be forgotten that they play important roles in the non-enzymatic protection of plants against oxidative stress and are the basic molecules that protect against photo-oxidative damage [
36]. In accordance with this, it is worth noting that the carotenoid content of hellebore tissues was highest during rooting under fluorescent light, which led to the weakest rooting (the fewest roots), but, above all, the least plant growth during the whole cycle. This may indicate that the plants are adversely affected by this light source, which, with higher concentrations of carotenoids, may be perceived as a stress factor. This theory seems to be correct insofar as white LED light, which allowed plants to achieve the best multiplication and produce the highest number of roots (above seven) at the rooting stage, resulted in the lowest synthesis of carotenoids compared to traditional FL light. In a study on
Lippia filifolia, white LED light provided higher content of both chlorophylls and carotenoids [
37]. The white LED was least effective in inducing hellebore plant growth rate compared to other light types, in spite of its positive influence in generating the highest shoot and root numbers. Often, faster elongation growth, or gain, is a plant response to a stress factor. For example, the leaf elongation rate of cereals is one of the most expressive plant responses to stress. Other plants may show increased root growth during mild drought or increased stem growth in response to low light. In such cases, the stress-induced growth is usually achieved by sacrificing the growth of other parts of that plant [
38]. Hence, there are probably more regenerated shoots and roots under white LED light, with weaker hellebore growth during micropropagation, compared to the other colors.
It is easy to see that most of the parameters studied during hellebore micropropagation are more significantly influenced by the source or color of the light used than by its intensity. The colors or LED combinations commonly used in in vitro cultures are white, red, blue and a mixture of blue and red [
17], hence the choice of these three basic LED colors in this research. Many studies find that blue LEDs are a good light source for inducing chlorophyll synthesis, while red LEDs reduce chlorophyll content. This is confirmed, for example, by Jao et al. [
21], who found that blue LEDs promote growth and increase chlorophyll content in
Zantedeschia jucunda, or by Verma et al. [
39] on
Digitalis purpurea, where chlorophyll a content was higher under blue light. In the present study, in hellebore, the effect of light color on chlorophyll content was significant, but it should be noted that there were definite differences depending on the stage of micropropagation. During multiplication, the highest content of pigments (both chlorophyll and carotenoids) was characterized by plants regenerating under red LED light, while during rooting, such an effect was achieved by traditional FL light or blue LED as indicated by other publications. This may be related to the condition of the plants, which have a different need for particular light spectrums during the intensive shoot proliferation phase than during rooting or as complete plants. Zheng et al. [
20] indicated that both red and blue light significantly increased the chlorophyll a/b ratio. However, more detailed studies using electron microscopy showed that blue light caused severe damage to the fine structure of chloroplasts in the early stages of leaf aging, while the degradation of chloroplast ultrastructure was apparently delayed in red light throughout the experiment compared to other treatments. LED red light perhaps sufficiently protects aging leaves from photoinhibition so that leaf aging can be effectively delayed. Perhaps during the intensive multiplication of hellebore, a higher content of pigments, including chlorophyll, was observed under red light. The positive effect of red light on chlorophyll b content compared to blue LED (more than three times greater) was noted in
Digitalis [
39], and twice as much chlorophyll a+b content was observed in the micropropagation of
Vanilla planifolia [
40]. Red LED light also influenced the elongation growth of micropropagated hellebores, both during propagation and rooting. Similarly,
Lippia filifolia plants reached greater height under red and white LED light conditions and achieved higher biomass accumulation [
37]. Neither the color of the light nor its intensity affected the percentage of rooting or the number of roots, which is an adaptive mechanism of the underground parts of the plant. This indicates that light has no effect on hellebore rhizogenesis, although the more intense elongation growth of roots at lower intensities suggests that they receive light stimuli and stronger light weakens their elongation.
White LED light is the most favorable in terms of obtaining hellebore ‘Molly’s White’ in in vitro culture, giving the highest multiplication rate of longer shoots and, at a lower concentration, affecting the regeneration of the largest number of roots. Thus, it is the best alternative to traditional fluorescent lights, which showed a much weaker effect on hellebore micropropagation. Similarly, the growth of shoots and leaves of
Pyrus communis was more affected by LED lamps compared to fluorescent lamps (control) [
23]. This is probably due to the fact that fluorescent lamps produce a wide range of wavelengths (350–750 nm) that are unnecessary for plant development [
17]. Light-emitting diodes (LEDs) have recently emerged as an alternative to commercial micropropagation because they have a monochromatic spectrum, less thermal radiation and, most importantly for in vitro cultures, they provide light in the spectral region that is involved in photosynthesis and photomorphogenic reactions in plants [
17]. In addition to increasing the efficiency of in vitro hellebore multiplication, LED lighting is more environmentally friendly and allows significant electricity savings, as the electricity requirements of LED lamps are 10–100 times lower than conventional light sources [
41]. Despite the high setup costs, with daily, long-term use of this lighting, these costs are recouped [
16,
41,
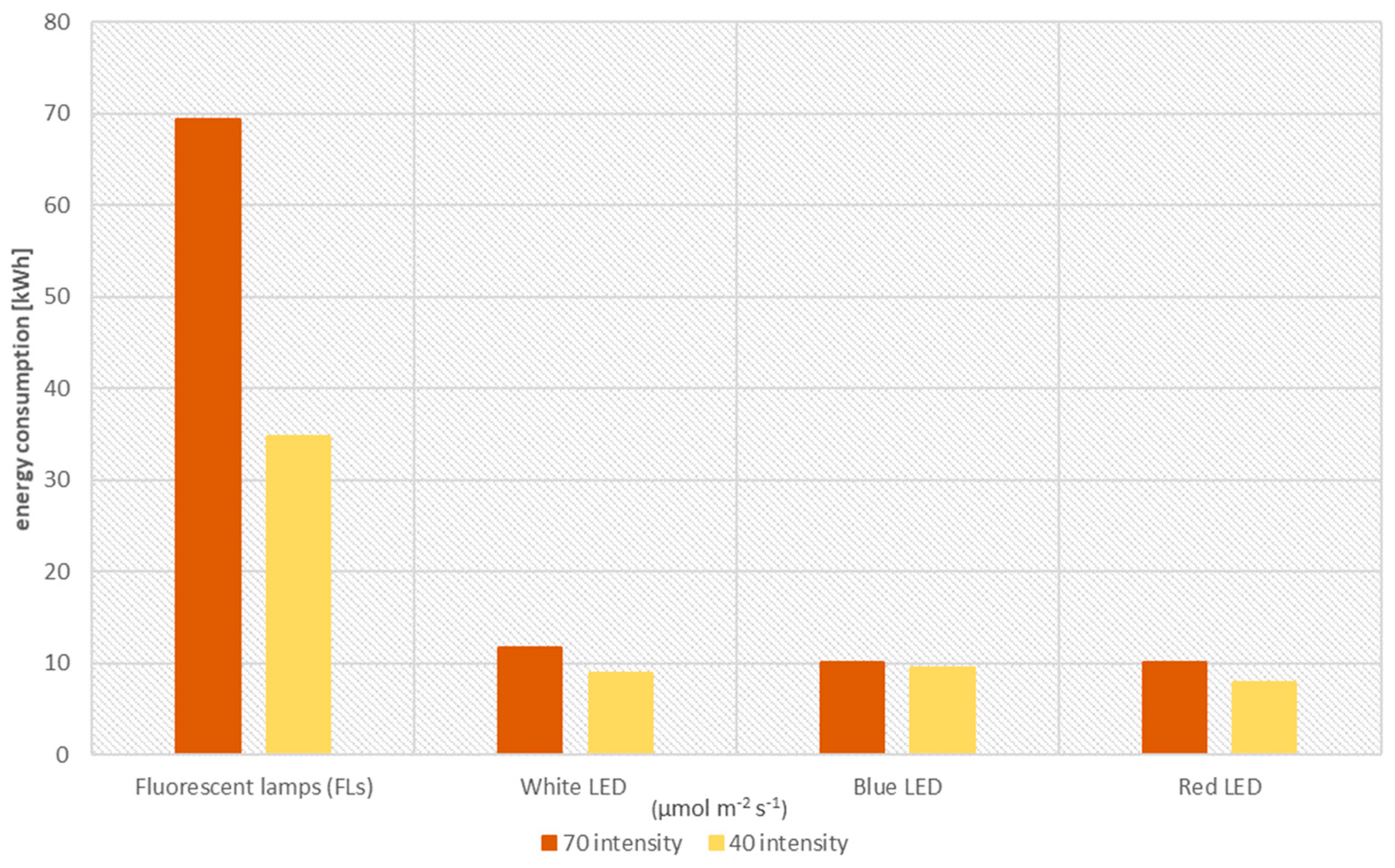
42]. As shown in our study, the differences between the higher and lower intensities of the LEDs used are not large. However, a huge difference is found when comparing any combination of LED light color with fluorescent lamps. LEDs used in our experiments consumed only 14.5–27.4% of what fluorescent lamps do in an 8-week hellebore production cycle (5 weeks of multiplication and 3 weeks of rooting). Even the highest intensity of 70 μmol·m
−2·s
−1 saved more than 57 kWh.
Due to the different responses of plants to the various light parameters and the significant impact of LED light on crop production, further research into its use is recommended. It is also extremely important from the point of view of environmental policies and the use of the latest developments in sustainable economic development.
5. Conclusions
The aim of the presented research was to compare the effects of traditional fluorescent lamps to LED lighting on the micropropagation of a valuable plant such as hellebore. In addition, the actual energy consumption of the different light combinations used during the whole in vitro production cycle of this plant was analyzed. Our studies showed that most of the parameters studied are more influenced by the source or color of the light used during micropropagation than its intensity. However, the higher intensity (70 μmol/m2/s) has a significant effect on increasing the content of assimilation pigments in regenerating and rooting plants. We have shown that red LED light impacts the elongation growth of micropropagated hellebores, but we generally recommend the use of white LED light in laboratories. It is most favorable in terms of obtaining hellebore ‘Molly’s White’ in in vitro cultures, giving the highest rate of propagation of longer shoots and also, at a lower intensity, influencing the regeneration of the largest number of roots. An economically significant result is the demonstration that LED lighting at 70 μmol·m−2·s−1, regardless of color, requires seven times less energy during hellebore micropropagation than fluorescent lamps (FLs).
In vitro cultures are producing more and more plants every year, as the obtained material is of high quality and disease-free. Therefore, it is a good planting material for establishing crops and plantations. Due to new agro-environmental policies, the results presented are extremely important for plant production. Regardless of the plants produced in in vitro culture (whether ornamental species, crops or vegetables), the goal is to reduce the cost of this production, especially its significant component in the form of electricity, while maintaining high-quality material. Importantly, the results of our research show that LED lighting is not only beneficial to the environment, as it is several times more energy-efficient, but also allows a high micropropagation coefficient to be obtained while maintaining the analyzed parameters at a good level. The obtained results allow us to conclude that LEDs are the future of plant production, both those in in vitro culture and in other types of production under cover where plant lighting is required.








{kind=link}
{kind=link}
{kind=link}
{kind=link}