
Nutritional and Antinutritional Potentials of Sorghum: A Comparative Study among Different Sorghum Landraces of Tigray, Northern Ethiopia

 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
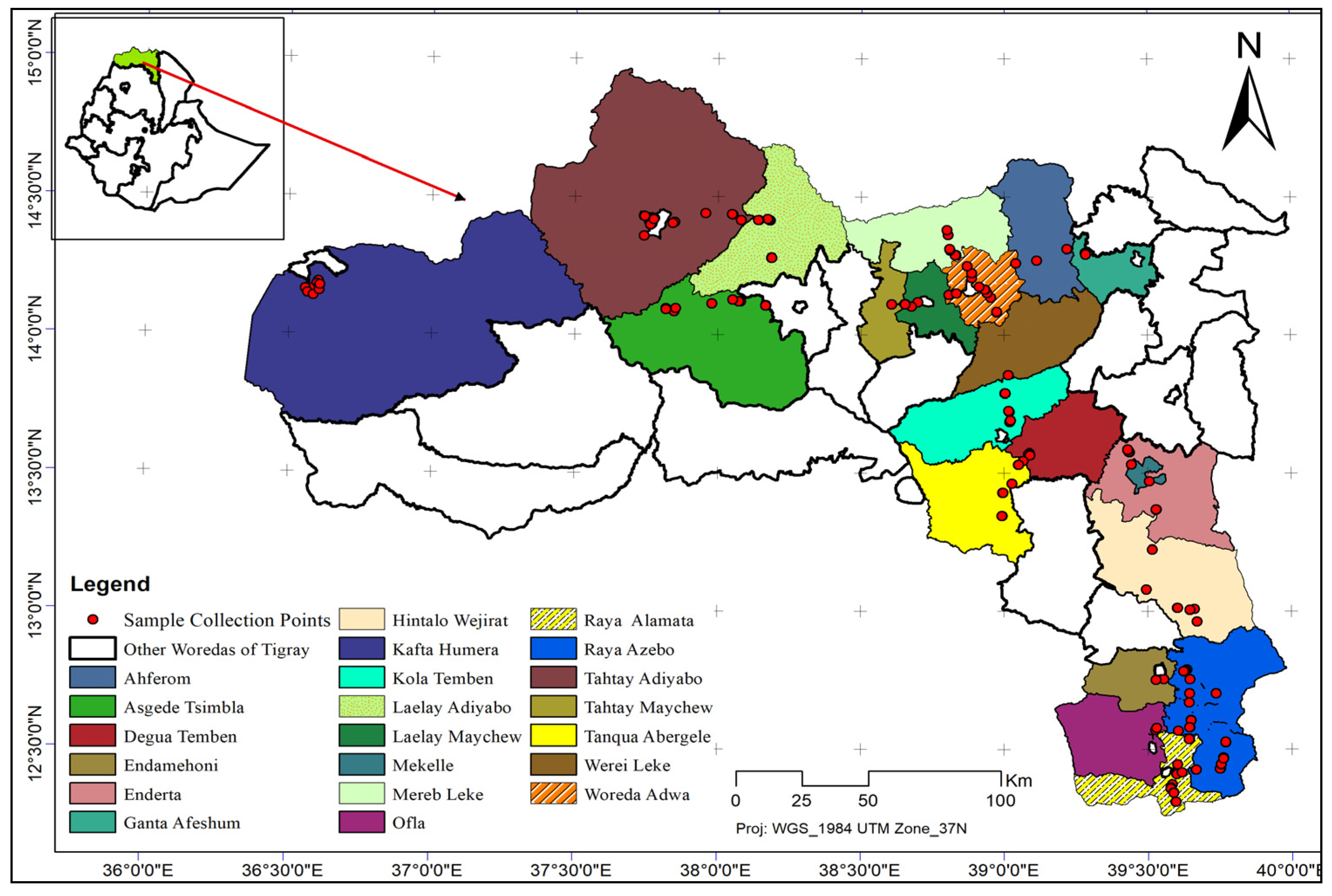
2.1. Plant Materials
2.2. Sample Preparation and Extraction
2.3. Determination of Protein and Starch
2.4. Estimation of Mineral Concentration
2.5. Determination of Phenolic Compounds
2.5.1. Determination of Total Flavonoid Content
2.5.2. Determination of Total Tannin Content
2.6. Determination of Antioxidant Capacity
2.6.1. DPPH Radical Scavenging Assay
2.6.2. Ferric Reducing Antioxidant Power
2.6.3. Total Antioxidant Capacity with Phosphomolybdenum Assay
2.7. Statistical Data Analysis
3. Results
3.1. Variabilities for Protein and Starch
3.2. Variabilities for Mineral Elements
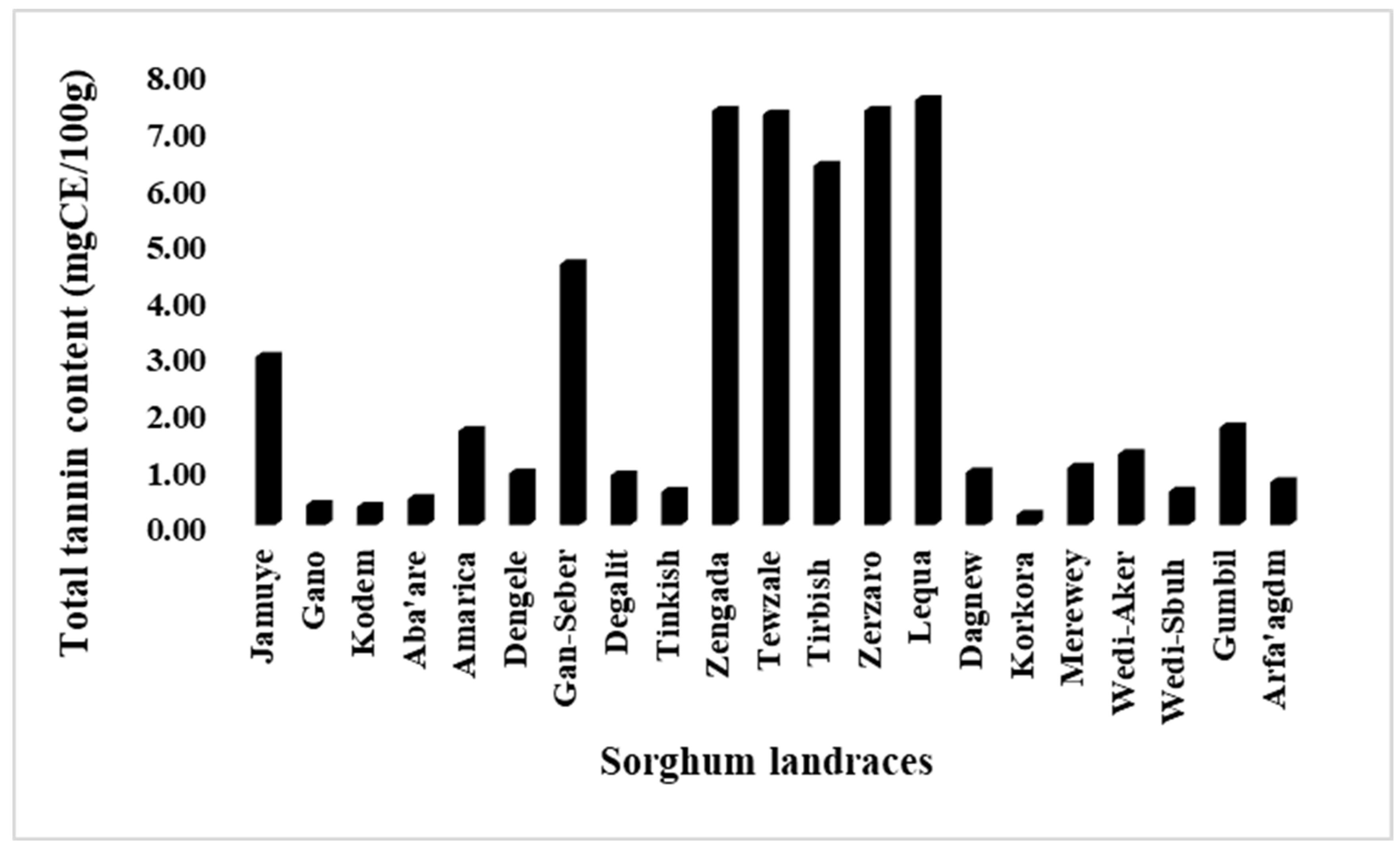
3.3. Variabilities for Total Flavonoid and Tannin Contents
3.4. Variabilities for Antioxidant Activities
3.5. Relationships of Seed Color with Phenolics and Antioxidant Activities
3.6. Grouping and Ordination of Genotypes
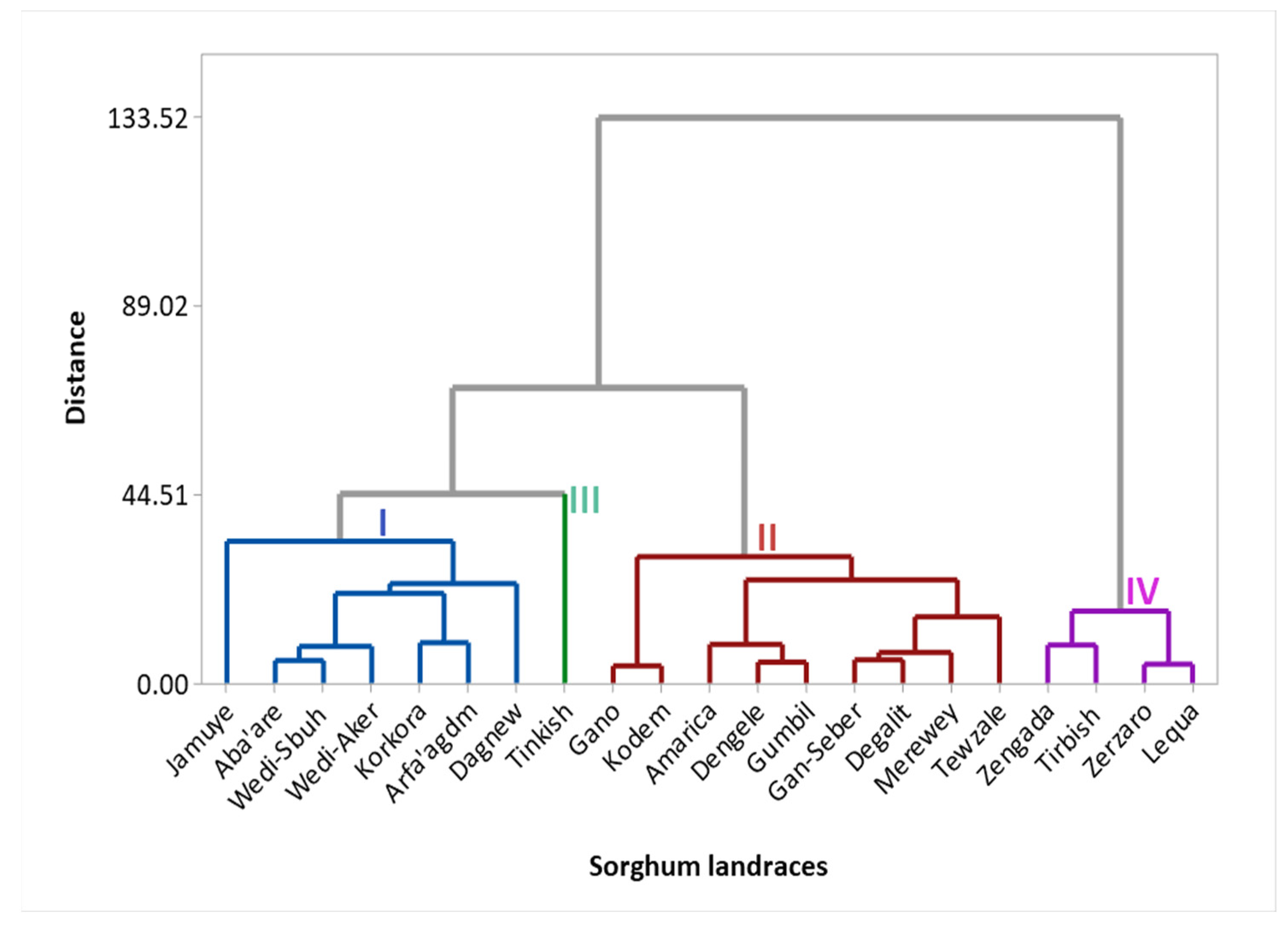
3.6.1. Cluster Analysis
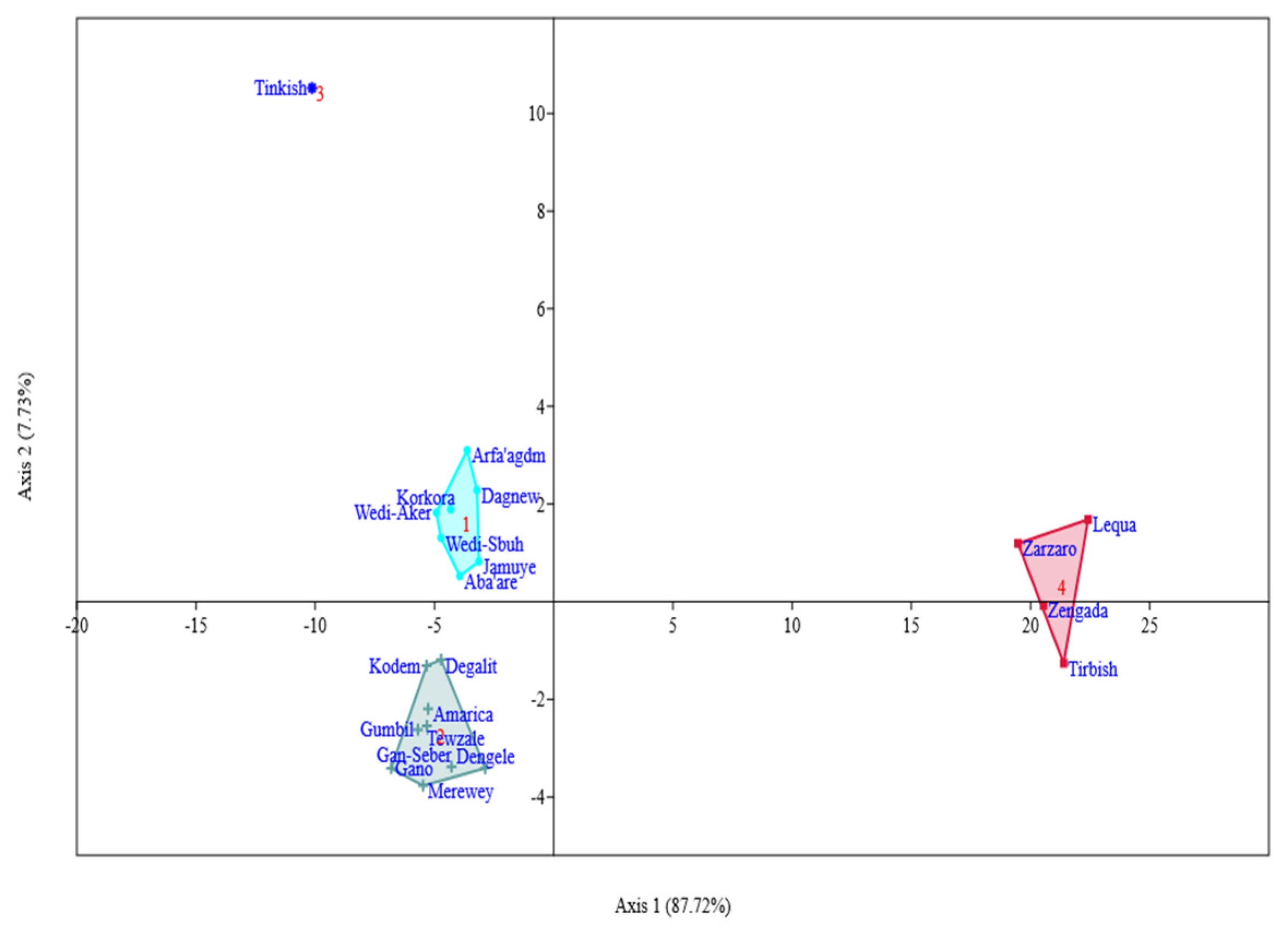
3.6.2. Principal Component Analysis
4. Discussion
4.1. Proximate Analysis
4.2. Phenolic Compounds and Antioxidant Activities
4.3. Relationship of Seed Color with Phenolic Compounds and Antioxidant Activities
4.4. Genotypes Grouping
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Singh, R.; Govindan, V. Zinc-Biofortified Wheat: Harnessing Genetic Diversity for improved Nutritional Quality. In Science Brief: Biofortification; No. 1. CIMMYT; HarvestPlus, and the Global Crop Trust: Bonn, Germany, 2017. [Google Scholar]
- Development Initiatives. Global Nutrition Report 2017: Nourishing the SDGs; Development Initiatives: Bristol, UK, 2017. [Google Scholar]
- Maberly, G.F.; Trowbridge, F.L.; Yip, R.; Sullivan, K.M.; West, C.E. Programs against micronutrient malnutrition: Ending hidden hunger. Annu. Rev. Public Health 1994, 15, 277–301. [Google Scholar] [CrossRef] [PubMed]
- Mayer, J.E.; Pfeiffer, W.H.; Beyer, P. Biofortified crops to alleviate micronutrient malnutrition. Curr. Opin. Plant Biol. 2008, 11, 166–170. [Google Scholar] [CrossRef]
- Teshome, A.; Baum, B.R.; Fahrig, L.; Torrance, J.K.; Arnason, T.J.; Lambert, J.D. Sorghum (Sorghum bicolor (L.) Moench) landrace variation and classification in North Shewa and South Welo, Ethiopia. Euphytica 1997, 97, 255–263. [Google Scholar] [CrossRef]
- Villa, T.C.C.; Maxted, N.; Scholten, M.; Ford-Lloyd, B. Defining and identifying crop landraces. Plant Genet. Resour. 2005, 3, 373–384. [Google Scholar] [CrossRef]
- Seboka, B.; van Hintum, T. The dynamics of on-farm management of sorghum in Ethiopia: Implication for the conservation and improvement of plant genetic resources. Genet. Resour. Crop Evol. 2006, 53, 1385–1403. [Google Scholar] [CrossRef]
- Newton, A.C.; Akar, T.; Baresel, J.P.; Bebeli, P.J.; Bettencourt, E.; Bladenopoulos, K.V.; Czembor, J.H.; Fasoula, D.A.; Katsiotis, A.; Koutis, K.; et al. Cereal landraces for sustainable agriculture. Sustain. Agric. 2011, 2, 147–186. [Google Scholar]
- Central Statistical Agency (CSA). Agricultural Sample Survey 2015/2016 Volume 1, a Report on Area and Production of Major Crops; Statistical Bulletin: Addis Ababa, Ethiopia, 2016. [Google Scholar]
- Teshome, A.; Patterson, D.; Asfew, Z.; Torrance, J.K.; Arnason, J.T. Changes of Sorghum bicolor landrace diversity and farmers’ selection criteria over space and time, Ethiopia. Genet. Resour. Crop Evol. 2007, 54, 1219–1233. [Google Scholar] [CrossRef]
- Tsehaye, Y.; Abera, Z.; Kebede, A.; Ghebremichael, B. A dynamic sorghum (Sorghum bicolor (L.) Moench) diversity management in situ and livelihood resilience in South and Central Tigray Region, Ethiopia. Momona Ethiop. J. Sci. 2009, 1. [Google Scholar] [CrossRef]
- Duodu, K.G.; Taylor, J.R.N.; Belton, P.S.; Hamaker, B.R. Factors affecting sorghum protein digestibility. J. Cereal Sci. 2003, 38, 117–131. [Google Scholar] [CrossRef]
- Saleh, S.M.; Zhang, Q.; Chen, J.; Shen, Q. Millet grains: Nutritional quality, processing, and potential health benefits. Compr. Rev. Food Sci. Food Saf. 2013, 12, 281–295. [Google Scholar] [CrossRef]
- Rao, S.; Santhakumar, A.B.; Chinkwo, K.A.; Wu, G.; Johnson, S.K.; Blanchard, C.L. Characterization of phenolic compounds and antioxidant activity in sorghum grains. J. Cereal Sci. 2018, 84, 103–111. [Google Scholar] [CrossRef]
- Shegro, A.; Shargie, N.G.; van Biljon, A.; Labuschagne, M.T. Diversity in starch, protein and mineral composition of sorghum landrace accessions from Ethiopia. J. Crop Sci. Biotechnol. 2012, 15, 275–280. [Google Scholar] [CrossRef]
- Shegro, A.; Labuschagne, M.T.; Shargie, N.G.; van Biljon, A. Multivariate analysis of nutriotonal diversity in sorghum accesions from wetsern Ethiopia. J. Biol. Sci. 2013, 13, 67–74. [Google Scholar] [CrossRef]
- Mofokeng, M.A. Diversity Analysis of South African Sorghum Genotypes Using Agronomic Traits, SSR Markers and Protein Content and Amino Acid Composition. Ph.D. Thesis, University of KwaZulu-Natal, Pietermaritzburg, South Africa, 2015. [Google Scholar]
- de Morais Cardoso, L.; Pinheiro, S.S.; Martino, H.S.D.; Pinheiro-Sant’Ana, H.M. Sorghum (Sorghum bicolor L.): Nutrients, bioactive compounds, and potential impact on human health. Crit. Rev. Food Sci. Nutr. 2017, 57, 372–390. [Google Scholar] [CrossRef]
- Nguni, D.; Geleta, M.; Hofvander, P.; Fatih, M.; Bryngelsson, T. Comparative genetic diversity and nutritional quality variation among some important southern African sorghum accessions [Sorghum bicolor (L.) Moench]. Aust. J. Crop Sci. 2012, 6, 56–64. [Google Scholar]
- Shewayrga, H.; Sopade, P.A.; Jordan, D.R.; Godwin, I.D. Characterisation of grain quality in diverse sorghum germplasm using a Rapid Visco-Analyzer and near infrared reflectance spectroscopy. J. Sci. Food Agric. 2012, 92, 1402–1410. [Google Scholar] [CrossRef]
- Jambunathan, R.; Subramanian, V. Grain quality and utilization of sorghum and pearl millet. Biotechnol. Trop. Crop Improv. 1988, 133–139. [Google Scholar]
- Njuguna, V.W.; Cheruiyot, E.K.; Mwonga, S.; Rono, J.K. Effect of genotype and environment on grain quality of sorghum (Sorghum bicolor L. Moench) lines evaluated in Kenya. Afr. J. Biotechnol. 2018, 12, 324–330. [Google Scholar]
- Pietta, P.G. Flavonoids as antioxidants. J. Nat. Prod. 2000, 63, 1035–1042. [Google Scholar] [CrossRef]
- Dykes, L.; Rooney, L.W. Sorghum and millet phenols and antioxidants. J. Cereal Sci. 2006, 44, 236–251. [Google Scholar] [CrossRef]
- Dykes, L.; Rooney, L. Phenolic compounds in cereal grains and their health benefits. Cereal Foods World 2007, 52, 105–111. [Google Scholar] [CrossRef]
- Dykes, L.; Rooney, W.L.; Rooney, L.W. Evaluation of phenolics and antioxidant activity of black sorghum hybrids. J. Cereal Sci. 2013, 58, 278–283. [Google Scholar] [CrossRef]
- Dykes, L.; Rooney, L.W.; Waniska, R.D.; Rooney, W.L. Phenolic compounds and antioxidant activity of sorghum grains of varying genotypes. J. Agric. Food Chem. 2005, 53, 6813–6818. [Google Scholar] [CrossRef]
- Awika, J.M.; Rooney, L.W. Sorghum phytochemicals and their potential impact on human health. Phytochemistry 2004, 65, 1199–1221. [Google Scholar] [CrossRef] [PubMed]
- Available online: https://www.perkinelmer.com/product/da-7250-at-line-nir-instrument-perdab (accessed on 12 February 2022).
- Brauteseth, E.M. The Mutagenesis of Sorghum bicolour (L.) Moench towards Improved Nutrition and Agronomic Performance. Doctoral Dissertation, University of KwaZulu-Natal, Pietermaritzburg, South Africa, 2009. [Google Scholar]
- Melash, A.A.; Mengistu, D.K. Improving grain micronutrient content of durum wheat (Triticum turgidum var. durum) through agronomic biofortification to alleviate the hidden hunger. Adv. Agric. 2020, 2020, 1–6. [Google Scholar] [CrossRef]
- Quettier-Deleu, C.; Gressier, B.; Vasseur, J.; Dine, T.; Brunet, C.; Luyckx, M.; Cazin, M.; Cazin, J.C.; Bailleul, F.; Trotin, F. Phenolic compounds and antioxidant activities of buckwheat (Fagopyrum esculentum Moench) hulls and flour. J. Ethnopharmacol. 2000, 72, 35–42. [Google Scholar] [CrossRef]
- Chew, K.K.; Khoo, M.Z.; Ng, S.Y.; Thoo, Y.Y.; Aida, W.W.; Ho, C.W. Effect of ethanol concentration, extraction time and extraction temperature on the recovery of phenolic compounds and antioxidant capacity of Orthosiphon stamineus extracts. Int. Food Res. J. 2011, 18, 1427. [Google Scholar]
- Katerere, D.R.; Eloff, J. Antibacterial and antioxidant activity of Sutherlandia frutescens (Fabaceae), a reputed anti-HIV/AIDS. Phytomedicine Phytother. Res. 2005, 19, 779–781. [Google Scholar] [CrossRef]
- Engida, D. In Vitro Antioxidant and α-Amylase Inhibition Activities of Spiced Red Chili Paste (Datta) from South Ethiopia. Ethiop. Pharm. J. 2015, 31, 93–106. [Google Scholar] [CrossRef]
- Prieto, P.; Pineda, M.; Aguilar, M. Spectrophotometric quantitation of antioxidant capacity through the formation of a phosphomolybdenum complex: Specific application to the determination of vitamin E. Anal. Biochem. 1999, 269, 337–341. [Google Scholar] [CrossRef]
- Tasie, M.M.; Gebreyes, B.G. Characterization of nutritional, antinutritional, and mineral contents of thirty-five sorghum varieties grown in Ethiopia. Int. J. Food Sci. 2020, 8243617. [Google Scholar] [CrossRef]
- FAO. Sorghum and Pearl Millets in Human Nutrition; Food and Agriculture Organization of the United Nations (FAO) Press: Rome, Italy, 1995. [Google Scholar]
- BSTID-NRC (Board on Science and Technology for International Development-National Research Council). Lost Crops of Africa; Academic Press: Washington, DC, USA, 1996; pp. 127–213. [Google Scholar]
- Singh, R.; Axtell, J.D. High lysine mutant gene (hl) that improves protein quality and biochemical value of grain sorghum. Crop Sci. 1973, 13, 535–539. [Google Scholar] [CrossRef]
- Bing, Y.I.; Zhou, Y.F.; Gao, M.Y.; Zhang, Z.; Yi, H.; Yang, G.D.; Wenjuan, X.; Huang, R.D. Effect of drought stress during flowering stage on starch accumulation and starch synthesis enzymes in sorghum grains. J. Integr. Agric. 2014, 13, 2399–2406. [Google Scholar]
- Abdelhalim, T.S.; Kamal, N.M.; Hassan, A.B. Nutritional potential of wild sorghum: Grain quality of Sudanese wild sorghum genotypes (Sorghum bicolor L. Moench). Food Sci. Nutr. 2019, 7, 1529–1539. [Google Scholar] [CrossRef]
- Hariprasanna, K.; Agte, V.; Elangovan, M.; Patil, J.V. Genetic variability for grain iron and zinc content in cultivars, breeding lines and selected germplasm accessions of sorghum [Sorghum bicolor (L.) Moench]. Indian J. Genet. 2014, 74, 42–49. [Google Scholar] [CrossRef]
- Mohammed, N.; Ahmed, I.; Babiker, E. Nutritional evaluation of sorghum flour (Sorghum bicolor L. Moench) during processing of injera. Int. J. BioLife Sci. 2010, 6, 35–39. [Google Scholar]
- Chan, S.S.; Ferguson, E.L.; Bailey, K.; Fahmida, U.; Harper, T.B.; Gibson, R.S. The concentration of iron, calcium, zinc and phytate in cereals and legumes habitually consumed by infants living in East Lombok, Indones. J. Food Compos. Anal. 2007, 20, 609–617. [Google Scholar] [CrossRef]
- Clemens, S. Zn and Fe biofortification: The right chemical environment for human bioavailability. Plant Sci. 2014, 225, 52–57. [Google Scholar] [CrossRef] [PubMed]
- Alfieri, M.; Balconi, C.; Cabassi, G.; Habyarimana, E.; Redaelli, R. Antioxidant activity in a set of sorghum landraces and breeding lines. Maydica 2017, 62, 1–7. [Google Scholar]
- Habriyanman Habyarimana, E.; Dall’Agata, M.; De Franceschi, P.; Baloch, F.S. Genome-wide association mapping of total antioxidant capacity, phenols, tannins, and flavonoids in a panel of Sorghum bicolor and S. bicolor × S. halepense populations using multi-locus models. PLoS ONE 2019, 14, e0225979. [Google Scholar] [CrossRef]
- Punia, H.; Tokas, J.; Malik, A.; Sangwan, S. Characterization of phenolic compounds and antioxidant activity in sorghum [Sorghum bicolor (L.) Moench] grains. Cereal Res. Commun. 2021, 2021, 343–353. [Google Scholar] [CrossRef]
- Taleon, V.; Dykes, L.; Rooney, W.; Rooney, L. Effect of genotype and environment on flavonoid concentration and profile of black sorghum grains. J. Cereal Sci. 2012, 56, 470–475. [Google Scholar] [CrossRef]
- Awika, J.M. Sorghum: Its Unique Nutritional and Health-Promoting Attributes. In Gluten-Free Ancient Grains: Cereals, Pseudocereals, and Legumes: Sustainable, Nutritious, and Health-Promoting Foods for the 21st Century; 2017. [Google Scholar]
- Kruger, J.; Taylor, J.R.; Du, X.; De Moura, F.F.; Lönnerdal, B.; Oelofse, A. Effect of phytate reduction of sorghum, through genetic modification, on iron and zinc availability as assessed by an in vitro dialysability bioaccessibility assay, Caco-2 cell uptake assay, and suckling rat pup absorption model. Food Chem. 2013, 141, 1019–1025. [Google Scholar] [CrossRef] [PubMed]
- Rathore, S.; Karunakar, S.; Vivek, K. Millet Grain Processing, Utilization and its Role in Helath promotion: A review. Int. J. Nutr. Food Sci. 2016, 5, 318–329. [Google Scholar] [CrossRef]
- Elkin, R.G.; Freed, M.B.; Hamaker, B.R.; Zhang, Y.; Parsons, C.M. Condensed tannins are only partially responsible for variations in nutrient digestibilities of sorghum grain cultivars. J. Agric. Food Chem. 1996, 44, 848–853. [Google Scholar] [CrossRef]
- Sharma, I.; Mathur, M.; Singh, G.P.; Rishi, A. Quantitative estimation of phenolic and flavonoid content ns antioxidant activity of various extracts of different parts of Plumbago zeylanica Linn. Int. J. Drug Dev. Res. 2014, 6, 136–140. [Google Scholar]
- Negash, A.; Derbew, B.; Amanuel, Z.; Mitiku, H.; Girmay, G.; Atkilt, G. Tree aga And Harvesting Season Affected Physico-chemical and Bioactive Compounds of Elite Type of Gunda Gundo Orange (Citrus spp.) in the Nortehrn Ethiopia. Int. J. Fruit Sci. 2021, 21, 26–39. [Google Scholar] [CrossRef]
- Ljiljana, S.; Mihajlo, S.; Vensa, N.; Ljubisa, N.; Dusica, R.; Jasna, C.B.; Vensa, T. Antioxidant Activity and Total Phenolic and Flavonoid Contents of Hieracium pilosella L. Extracts. Sensors 2009, 9, 5702–5714. [Google Scholar] [CrossRef]
- Soualem, Z.M.; ALsayadi, M.; Gaouar, N.; Benammar, C.h.; Beghdada, C.h.; Belarbi, M. Antioxidant activity and nutrient composition of Sorghum bicolor L. and Secale cereale L. in Algeria. Acad. J. Food Res. 2013, 1, 59–65. [Google Scholar]
- Rohman, A.; Riyanto, S.; Yuniarti, N.; Saputra, W.R.; Utami, R.; Mulatsih, W. Antioxidant activity, total phenolic, and total flavaonoid of extracts and fractions of red fruit (Pandanus conoideus Lam). Int. Food Res. J. 2010, 17, 97–106. [Google Scholar]
- Shen, S.; Huang, R.; Li, C.; Wu, W.; Chen, H.; Shi, J.; Chen, S.; Ye, X. Phenolic Compositions and Antioxidant Activities Differ Significantly among Sorghum Grains with Different Applications. Molecules 2018, 23, 1203. [Google Scholar] [CrossRef] [PubMed]
- Awika, J.M. Sorghum Phenols as Antioxidants. Master’s Thesis, Texas A&M University, College Station, TX, USA, 2000. [Google Scholar]
- Semere, T.; Fjellheim, S.; Tsehaye, Y.; Westengen, O.T. Inventory of sorghum landraces diversity on-farm and adaptability responses to changing climate: Implications for sorghum breeding and conservation. Genet. Resour. Crop Evol. 2023, 1–18. [Google Scholar] [CrossRef]
- Tsehaye, Y.; Berg, T.; Tsegaye, B.; Tanto, T. Farmers’ management of finger millet (Eleusine coracana L.) diversity in Tigray, Ethiopia and implications for on-farm conservation. Biodivers. Conserv. 2006, 15, 4289–4308. [Google Scholar] [CrossRef]
- Hikeezi, D.M. The Importance of Sorghum Grain Colour and Hardness, and Their Causes and Measurement. International Sorghum and Millet Collaborative Research Support Program (INTSORMIL CRSP) Presentations. 2010. Available online: https://digitalcommons.unl.edu/intsormilpresent/18 (accessed on 3 May 2022).
- Harris, H.B.; Burns, R.E. Influence of tannin content on preharvest seed germination in sorghum seed molding. Agron. J. 1970, 65, 957–959. [Google Scholar] [CrossRef]
- Asfaw, Z. Variation in the morphology of the spike within Ethiopian barley, Hordeum vulgare L. (Poaceae). Acta Agric. Scand. 1988, 38, 277–288. [Google Scholar] [CrossRef]
- Deepika, K.; Krishna, L.; Santosha, R.; Chandra, M.Y.; Aravind, K.J.; Narender, R.; Subba, R.L.V.; Jyothi, B. Multivariate analysis of geographically diverse rice germplasm for genetic improvement of yield, dormancy and shattering-related traits. Plant Genet. Resour. Charact. Util. 2021, 19, 144–152. [Google Scholar] [CrossRef]
- Seiler, G.J.; Stafford, R.E. Factor analysis of components of yield in Guar. Crop Sci. 1985, 25, 905–908. [Google Scholar] [CrossRef]
- Yan, W.; Fregeau-Reid, J.A. Breeding line selection based on multiple traits. Crop Sci. 2008, 48, 417–423. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Landrace Name | Zone of Origin | Race | Seed Color | Preferred End-Use |
|---|---|---|---|---|---|
| 1 | Jamuye | Southern Tigray | Durra | Yellow | Swa/Enjera |
| 2 | Gano | Southern Tigray | Durra | Yellow | Enjera |
| 3 | Kodem | Southern Tigray | Durra | Yellow | Enjera/Swa |
| 4 | Aba’are | Southern Tigray | Durra | Yellow | Enjera |
| 5 | Amarica | Southern Tigray | Durra | White | Enjera |
| 6 | Dengele | Southern Tigray | Durra | Yellow | Enjera/Swa |
| 7 | Gan-Seber | Southern Tigray | Durra | White | Swa/Enjera |
| 8 | Degalit | Southern Tigray | Durra | Yellow | Swa/Enjera |
| 9 | Tinkish | Southern Tigray | Durra | Yellow | Enjera/popping |
| 10 | Zengada | Southern Tigray | Bicolor | Red | Enjera/Swa |
| 11 | Tewzale | Central Tigray | Caudatum | Red | Enjera/Swa |
| 12 | Tirbish | Central Tigray | Durra | Yellow | Swa/Enjera |
| 13 | Zerzaro | Central Tigray | Durra | Brown | Swa/Enjera |
| 14 | Lequa | Central Tigray | Bicolor | Brown | Swa |
| 15 | Dagnew | Western Tigray | Bicolor | Yellow | Swa/Enjera |
| 16 | Korkora | Western Tigray | Caudatum | White | Enjera |
| 17 | Merewey | Western Tigray | Durra | Yellow | Enjera |
| 18 | Wedi-Aker | Western Tigray | Caudatum | White | Swa |
| 19 | Wedi-Sbuh | Western Tigray | Durra | White | Enjera |
| 20 | Gumbil | Central Tigray | Durra | Yellow | Enjera |
| 21 | Arfa’agdm | Western Tigray | Caudatum | White | Enjera |
| Variable | Protein | Starch | Fe | Zn | Cu | Mn | Cr | TFC | TTC | EC50 | FRAP | TAC |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Max | 18 | 78.3 | 101 | 43 | 5.3 | 20.2 | 1.5 | 665 | 7.5 | 818.4 | 334.8 | 63.9 |
| Min | 6.21 | 33.4 | 32 | 16.9 | 1.5 | 9.21 | 0 | 0 | 0.18 | 29.1 | 17.9 | 1.71 |
| Mean | 11.4 | 69.3 | 51.6 | 27 | 2.9 | 14.7 | 1.01 | 198.4 | 2.61 | 208 | 115 | 23.5 |
| SE | 0.12 | 0.32 | 3.29 | 1.42 | 0.2 | 0.55 | 0.1 | 46.7 | 0.61 | 48.6 | 23.1 | 4.45 |
| SD | 2.2 | 6.12 | 15.1 | 6.5 | 0.9 | 2.51 | 0.35 | 214.2 | 2.78 | 222.6 | 105.9 | 20.4 |
| CV | 19.34 | 8.83 | 29.2 | 24.1 | 32 | 17.1 | 34.3 | 108 | 106.5 | 106.9 | 92.35 | 86.8 |
| t-test | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 |
| Mean of Nutritional Traits | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Cluster | Protein | Starch | Fe | Zn | Cu | Mn | Cr | TFC | TTC | EC50 | FRAP | TAC |
| I | 13.17 | 72.38 | 59.99 | 32.48 | 3.7 | 16.24 | 1.12 | 119.6 | 1.02 | 231.6 | 62 | 12.39 |
| II | 10.27 | 72.8 | 45.01 | 23.13 | 2.32 | 14.02 | 0.85 | 121.3 | 2.08 | 199.6 | 82 | 17.47 |
| III | 14.67 | 53 | 55.49 | 25.19 | 2.91 | 14.24 | 1.39 | 0 | 0.59 | 818.37 | 18 | 1.71 |
| IV | 10.34 | 67.88 | 50.74 | 26.28 | 2.83 | 13.7 | 1.07 | 559.3 | 7.12 | 34.8 | 303 | 61.94 |
| Clusters | I | II | III | IV |
|---|---|---|---|---|
| I | - | |||
| II | 40.18 ** | - | ||
| III | 212.12 ** | 238.15 ** | - | |
| IV | 673.98 ** | 729.15 ** | 1173.79 ** | - |
| Trait | PC 1 | PC 2 | PC 3 | PC 4 |
|---|---|---|---|---|
| Protein | −0.43 | 0.38 | −0.27 | 0.32 |
| Starch | −0.07 | −0.25 | 0.89 | −0.02 |
| Fe | −0.17 | 0.75 | −0.06 | −0.07 |
| Zn | −0.13 | 0.65 | 0.53 | 0.27 |
| Cu | −0.22 | 0.91 | 0.01 | −0.21 |
| Mn | −0.22 | 0.45 | 0.16 | −0.59 |
| Cr | 0.19 | 0.28 | −0.05 | 0.72 |
| TFC | 0.89 | 0.24 | 0.05 | 0.15 |
| TTC | 0.88 | 0.15 | −0.24 | −0.21 |
| EC50 | −0.69 | −0.01 | −0.56 | −0.02 |
| FRAP | 0.97 | 0.17 | −0.03 | 0.01 |
| TAC | 0.96 | 0.10 | −0.14 | −0.11 |
| Eigenvalue | 4.26 | 2.42 | 1.57 | 1.17 |
| % Total variance | 35.54 | 20.16 | 13.08 | 9.71 |
| % Cumulative variance | 35.54 | 55.70 | 68.78 | 78.48 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Semere, T.; Tsehaye, Y.; Tareke, L.; Westengen, O.T.; Fjellheim, S. Nutritional and Antinutritional Potentials of Sorghum: A Comparative Study among Different Sorghum Landraces of Tigray, Northern Ethiopia. Agriculture 2023, 13, 1753. https://doi.org/10.3390/agriculture13091753
Semere T, Tsehaye Y, Tareke L, Westengen OT, Fjellheim S. Nutritional and Antinutritional Potentials of Sorghum: A Comparative Study among Different Sorghum Landraces of Tigray, Northern Ethiopia. Agriculture. 2023; 13(9):1753. https://doi.org/10.3390/agriculture13091753
Chicago/Turabian StyleSemere, Tesfakiros, Yemane Tsehaye, Lijalem Tareke, Ola T. Westengen, and Siri Fjellheim. 2023. "Nutritional and Antinutritional Potentials of Sorghum: A Comparative Study among Different Sorghum Landraces of Tigray, Northern Ethiopia" Agriculture 13, no. 9: 1753. https://doi.org/10.3390/agriculture13091753
APA StyleSemere, T., Tsehaye, Y., Tareke, L., Westengen, O. T., & Fjellheim, S. (2023). Nutritional and Antinutritional Potentials of Sorghum: A Comparative Study among Different Sorghum Landraces of Tigray, Northern Ethiopia. Agriculture, 13(9), 1753. https://doi.org/10.3390/agriculture13091753