
A Glimpse into the Genetic Heritage of the Olive Tree in Malta

 ,
,  ,
,  ,
,  ,
,  , ,
, , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. DNA Extraction
2.3. Olive Genotyping
2.4. References for Comparison
2.5. Data Analysis
2.5.1. Genetic Diversity
2.5.2. Genetic Relationships and Population Structure
3. Results
3.1. Genetic Relationships and Genetic Diversity of Maltese Genotypes
3.2. Genetic Diversity of Maltese Genotypes
3.3. Genetic Relationships of the Maltese Germplasm with the Mediterranean Germplasm
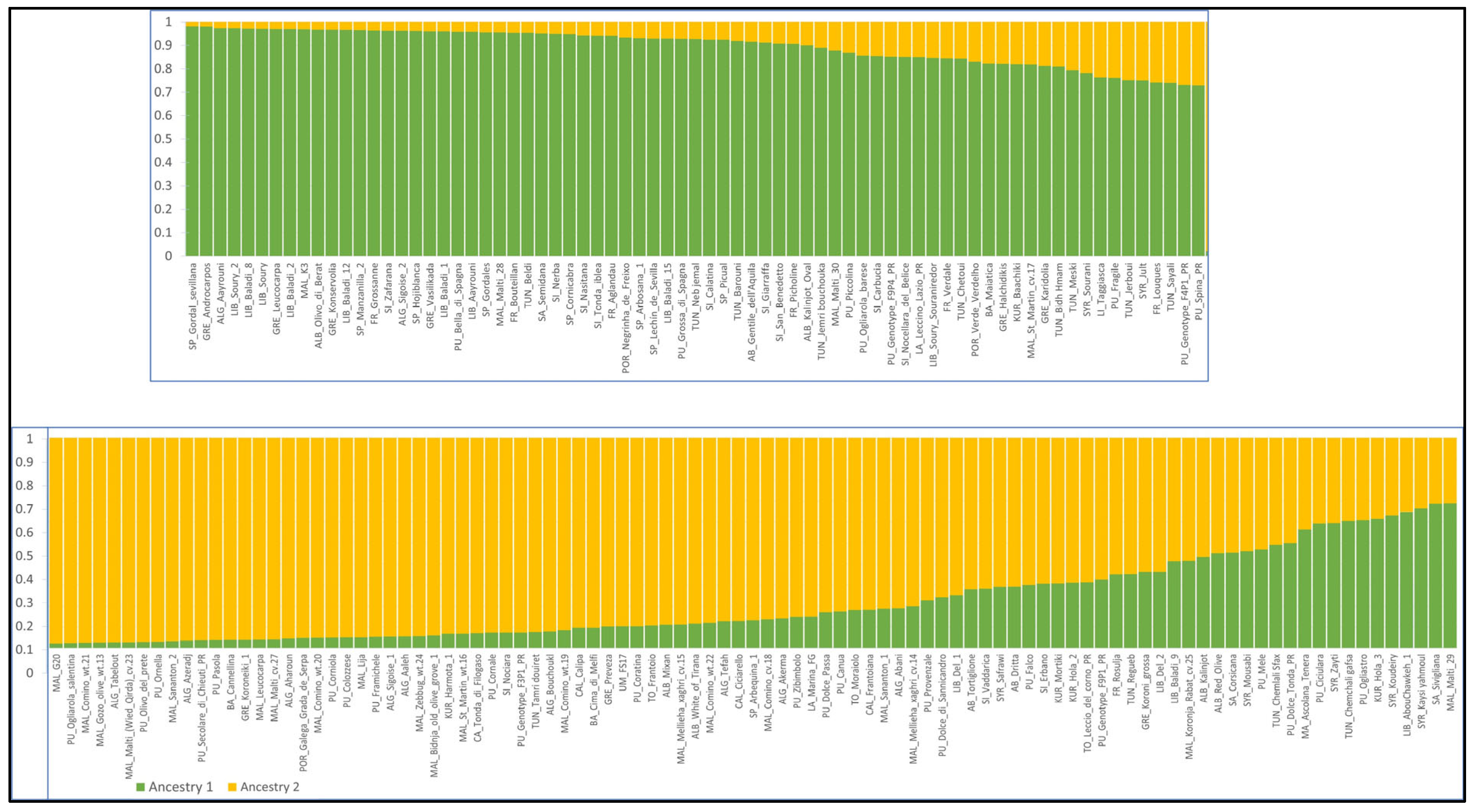
3.4. Population Structure
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Borg, J. Cultivation and Diseases of Fruit Trees in the Maltese Islands; Malta Government Printing Office: Marsa, Malta, 1922; pp. 1–617. [Google Scholar]
- Abela, F.G. Della Descrittione di Malta, Isola Nel Mare Siciliano Con le Sue Antichità, Ed Altre Notitie; Paolo Bonacota: Valletta, Malta, 1647; pp. 1–573. [Google Scholar]
- Valeri, M.C.; Mifsud, D.; Sammut, C.; Pandolfi, S.; Lilli, E.; Bufacchi, M.; Stanzione, V.; Passeri, V.; Baldoni, L.; Mariotti, R.; et al. Exploring Olive Genetic Diversity in the Maltese Islands. Sustainability 2022, 14, 10684. [Google Scholar] [CrossRef]
- Passeri, V.; Sammut, C.; Mifsud, D.; Domesi, A.; Stanzione, V.; Baldoni, L.; Mousavi, S.; Mariotti, R.; Pandolfi, S.; Cinosi, N.; et al. The Ancient Olive Trees (Olea europaea L.) of the Maltese Islands: A Rich and Unexplored Patrimony to Enhance Oliviculture. Plants 2023, 12, 1988. [Google Scholar] [CrossRef]
- Mazzitelli, O.; Calleja, A.; Sardella, D.; Farrugia, C.; Zammit-Mangion, M. Analysis of the molecular diversity of Olea europaea in the Mediterranean Island of Malta. Genet. Resour. Crop Evol. 2015, 62, 1021–1027. [Google Scholar] [CrossRef]
- Lageard, J.G.; Sultana, D.; Brearley, F.Q. Veteran trees in an historic landscape: The Bidnija olive grove, Malta. J. Archaeol. Sci. Rep. 2021, 38, 103094. [Google Scholar] [CrossRef]
- Buhagiar, J. Perspectives on olive cultivation and pro-cessing in Maltese roman antiquity. In The Zejtuna Roman Villa—Research, Conservation, Management; Wirt iz-Zejtun Publication, Gutenberg Press Ltd.: Tarxien, Malta, 2012. [Google Scholar]
- Gambin, T. A drop in the Ocean: Malta’s Trade in olive oil during the Roman Period. In The Zejtuna Roman Villa—Research, Conservation, Management; Wirt iz-Zejtun Publication, Gutenberg Press Ltd.: Tarxien, Malta, 2012. [Google Scholar]
- Gatt, L.; Lia, F.; Zammit-Mangion, M.; Thorpe, S.J.; Scembri-Wismayer, P. First Profile of Phenolic Compounds from Maltese Extra Virgin Olive Oils Using Liquid-Liquid Extraction and Liquid Chromatography-Mass Spectrometry. J. Oleo Sci. 2021, 70, 145–153. [Google Scholar] [CrossRef] [PubMed]
- Di Rienzo, V.; Sion, S.; Taranto, F.; D’Agostino, N.; Montemurro, C.; Fanelli, V.; Sabetta, W.; Boucheffa, S.; Tamendjari, A.; Pasqualone, A.; et al. Genetic flow among olive populations within the Mediterranean basin. PeerJ 2018, 6, e5260. [Google Scholar] [CrossRef] [PubMed]
- Pasqualone, A.; Di Rienzo, V.; Nasti, R.; Blanco, A.; Gomes, T.; Montemurro, C. Traceability of Italian protected designation of origin (PDO) table olives by means of microsatellite molecular markers. J. Agric. Food Chem. 2013, 61, 3068–3073. [Google Scholar] [CrossRef]
- Pasqualone, A.; Di Rienzo, V.; Miazzi, M.M.; Fanelli, V.; Caponio, F.; Montemurro, C. High resolution melting, analysis of DNA microsatellites in olive pastes and virgin olive oils obtained by talc addition. Eur. J. Lipid Sci. Technol. 2015, 117, 2044–2048. [Google Scholar] [CrossRef]
- Sabetta, W.; Miazzi, M.M.; Di Rienzo, V.; Fanelli, V. Development and application of protocols to certify the authenticity and traceability of Apulian typical products in olive sector. Riv. Ital. Sost. Grasse 2016, 94, 37–43. [Google Scholar]
- Di Rienzo, V.; Miazzi, M.M.; Fanelli, V.; Sabetta, W.; Montemurro, C. The preservation and characterization of Apulian olive germplasm biodiversity. Acta Hortic. 2018, 1199, 1–6. [Google Scholar] [CrossRef]
- Muzzalupo, I.; Vendramin, G.G.; Chiappetta, A. Genetic biodiversity of Italian olives (Olea europaea) germplasm analyzed by SSR markers. Sci. World J. 2014, 2014, 296590. [Google Scholar] [CrossRef]
- Taranto, F.; D’Agostino, N.; Pavan, S.; Fanelli, V.; Di Renzo, V.; Sabetta, W.; Miazzi, M.M.; Zelasco, S.; Perri, E.; Montemurro, C. Single nucleotide polymorphism (SNP) diversity in an olive germplasm collection. Acta Hortic. 2016, 1199, 27–31. [Google Scholar] [CrossRef]
- Mousavi, S.; Mariotti, R.; Bagnoli, F.; Costantini, L.; Cultrera, N.G.M.; Arzani, K.; Pandolfi, S.; Vendramin, G.G.; Torkzaban, B.; Hosseini-Mazinani, M.; et al. The eastern part of the Fertile Crescent concealed an unexpected route of olive (Olea europaea L.) differentiation. Ann. Bot. 2017, 119, 1305–1318. [Google Scholar] [CrossRef]
- Boucheffa, S.; Tamendjari, A.; Sanchez-Gimeno, A.C.; Rovellini, P.; Venturini, S.; Di Rienzo, V.; Miazzi, M.M.; Montemurro, C. Diversity assessment of Algerian wild and cultivated olives (Olea europaea L.) by molecular, morphological, and chemical traits. Eur. J. Lipid Sci. Technol. 2019, 121, 1800302. [Google Scholar] [CrossRef]
- Saddoud Debbabi, O.; Miazzi, M.M.; Elloumi, O.; Fendri, M.; Ben Amar, F.; Savoia, M.; Sion, S.; Souabni, H.; Mnasri, S.R.; Ben Abdelaali, S.; et al. Recovery, Assessment, and Molecular Characterization of Minor Olive Genotypes in Tunisia. Plants 2020, 9, 382. [Google Scholar] [CrossRef]
- Spadoni, A.; Sion, S.; Gadaleta, S.; Savoia, M.A.; Piarulli, L.; Fanelli, V.; Di Rienzo, V.; Taranto, F.; Miazzi, M.M.; Montemurro, C.; et al. A simple and rapid method for genomic DNA extraction and microsatellite analysis in tree plants. J. Agric. Sci. Technol. 2019, 21, 1215–1226. [Google Scholar]
- Sefc, K.M.; Lopes, M.S.; Lefort, F.; Botta, R.; Roubelakis-Angelakis, K.A.; Ibáñez, J.; Pejić, I.; Wagner, H.W.; Glössl, J.; Steinkellner, H. Microsatellite variability in grapevine cultivars from different European regions and evaluation of assignment testing to assess the geographic origin of cultivars. Theor. Appl. Genet. 2000, 100, 498–505. [Google Scholar] [CrossRef]
- Carriero, F.; Fontanazza, G.; Cellini, F.; Giorio, G. Identification of simple sequence repeats (SSRs) in olive (Olea europaea L.). Theor. Appl. Genet. 2002, 104, 301–307. [Google Scholar] [CrossRef]
- De La Rosa, R.; James, C.M.; Tobutt, K.R. Isolation and characterization of polymorphic microsatellites in olive (Olea europaea L.) and their transferability to other genera in the Oleaceae. Mol. Ecol. Notes 2002, 2, 265–267. [Google Scholar] [CrossRef]
- Savoia, M.A.; Fanelli, V.; Miazzi, M.M.; Taranto, F.; Procino, S.; Susca, L.; Montilon, V.; Potere, O.; Nigro, F.; Montemurro, C. Apulian Autochthonous Olive Germplasm: A Promising Resource to Restore Cultivation in Xylella fastidiosa-Infected Areas. Agriculture 2023, 13, 1746. [Google Scholar] [CrossRef]
- Peakall, R. GENALEX 6: Genetic analysis in Excel. Population genetic software for teaching and research. Mol. Ecol. Notes 2006, 6, 288–295. [Google Scholar] [CrossRef]
- Gower, J.C. Wiley StatsRef: Statistics Reference Online; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2014; Volume 70. [Google Scholar]
- Lynch, M.; Ritland, K. Estimation of pairwise relatedness with molecular markers. Genetics 1999, 152, 1753–1766. [Google Scholar] [CrossRef]
- Perrier, X.; Flori, A.; Bonnot, F. Data Analysis Methods. In Genetic Diversity of Cultivated Tropical Plants; Hamon, P., Seguin, M., Perrier, X., Glaszmann, J.C., Eds.; Science Publishers: Enfield, NH, USA, 2003; pp. 43–76. [Google Scholar]
- Felsenstein, J. Confidence limits on phylogenies: An approach using the bootstrap. Evolution 1985, 39, 783–791. [Google Scholar] [CrossRef]
- Pritchard, J.K.; Stephens, M.; Donnelly, P. Inference of population structure using multilocus genotype data. Genetics 2000, 155, 945–959. [Google Scholar] [CrossRef]
- Earl, D.A.; Vonholdt, B. Structure Harverest: A website and program for visualizing STRUCTURE output and implementing the Evanno method. Conserv. Genet. Res. 2012, 4, 359–361. [Google Scholar] [CrossRef]
- Evanno, G.; Regnaut, S.; Goudet, J. Detecting the number of clusters of individuals using the software STRUCTURE: A simulation study. Mol. Ecol. 2005, 14, 2611–2620. [Google Scholar] [CrossRef]
- Díez, C.M.; Moral, J.; Cabello, D.; Morello, P.; Rallo, L.; Barranco, D. Cultivar and tree density as key factors in the long-term performance of super high-density olive orchards. Front. Plant Sci. 2016, 7, 1226. [Google Scholar] [CrossRef] [PubMed]
- Gros-Balthazard, M.; Besnard, G.; Sarah, G.; Holtz, Y.; Leclercq, J.; Santoni, S.; Wegmann, D.; Glémin, S.; Khadari, B. Evolutionary transcriptomics reveals the origins of olives and the genomic changes associated with their domestication. Plant J. 2019, 100, 143–157. [Google Scholar] [CrossRef] [PubMed]
- Farrugia, N.; Attard, E. Autochthonous and international olive cultivars in Malta: Morphometric characteristics and physicochemical analysis. Arch. Agric. Environ. Sci. 2021, 8, 15–22. [Google Scholar]
- Topi, D.; Guclu, G.; Kelebek, H.; Selli, S. Olive Oil Production in Albania, Chemical Characterization, and Authenticity. In Olive Oil—New Perspectives and Applications; BoD—Books on Demand; IntechOpen: London, UK, 2021; ISBN 1839684143 9781839684142. [Google Scholar]
- Dervishi, A.; Jakše, J.; Ismaili, H.; Javornik, B.; Štajner, N. Genetic Structure and Core Collection of Olive Germplasm from Albania Revealed by Microsatellite Markers. Genes 2021, 12, 256. [Google Scholar] [CrossRef]
- Breton, C.; Terral, J.F.; Pinatel, C.; Médail, F.; Bonhomme, F.; Bervillé, A. The origins of the domestication of the olive tree. Comptes Rendus Biol. 2009, 332, 1059–1064. [Google Scholar] [CrossRef]
- Díez, C.M.; Trujillo, I.; Martinez-Urdiroz, N.; Barranco, D.; Rallo, L.; Marfil, P.; Gaut, B.S. Olive domestication and diversification in the Mediterranean Basin. New Phytol. 2015, 206, 436–447. [Google Scholar] [CrossRef]
- Sion, S.; Taranto, F.; Montemurro, C.; Mangini, G.; Camposeo, S.; Falco, V.; Gallo, A.; Mita, G.; Saddoud Debbabi, O.; Ben Amar, F.; et al. Genetic Characterization of Apulian Olive Germplasm as Potential Source in New Breeding Programs. Plants 2019, 8, 268. [Google Scholar] [CrossRef]
- Chalak, L.; Noun, A.; Youssef, H.; Hamadeh, B. Diversity of loquats (Eriobotrya japonica Lindl.) cultivated in Lebanon as assessed by morphological traits. Sci. Hortic. 2014, 167, 135–144. [Google Scholar] [CrossRef]
- Terral, J.-F.; Alonso, N.; Buxó I Capdevila, R.; Chatti, N.; Fabre, L.; Fiorentino, G.; Marinval, P.; Jordá, G.P.; Pradat, B.; Rovira, N.; et al. Historical biogeography of olive domestication (Olea europaea L.) as revealed by geometrical morphometry applied to biological and archaeological material. J. Biogeogr. 2004, 31, 63–77. [Google Scholar] [CrossRef]
- Rosati, A.; Zipanćičn, M.; Caporali, S.; Padula, G. Fruit weight is related to ovary weight in olive (Olea europaea L.). Sci. Hortic. 2009, 122, 399–403. [Google Scholar] [CrossRef]
- Julca, I.; Marcet-Houben, M.; Cruz, F.; Gómez-Garrido, J.; Gaut, B.S.; Díez, C.M.; Gut, I.G.; Alioto, T.S.; Vargas, P.; Gabaldón, T. Genomic evidence for recurrent genetic admixture during the domestication of Mediterranean olive trees (Olea europaea L.). BMC Biol. 2020, 18, 148. [Google Scholar] [CrossRef] [PubMed]
- Besnard, G.; Terral, J.F.; Cornille, A. On the origins and domestication of the olive: A review and perspectives. Ann. Bot. 2018, 121, 385–403. [Google Scholar] [CrossRef] [PubMed]
- Belaj, A.; Dominguez-García, M.D.C.; Atienza, S.G.; Urdíroz, N.M.; De la Rosa, R.; Satovic, Z.; Martín, A.; Kilian, A.; Trujillo, I.; Valpuesta, V.; et al. Developing a core collection of olive (Olea europaea L.) based on molecular markers (DArTs, SSRs, SNPs) and agronomic traits. Tree Genet. Genomes 2012, 8, 365–378. [Google Scholar] [CrossRef]
- Debbabi, O.S.; Amar, F.B.; Rahmani, S.M.; Taranto, F.; Montemurro, C.; Miazzi, M.M. The Status of Genetic Resources and Olive Breeding in Tunisia. Plants 2022, 11, 1759. [Google Scholar] [CrossRef] [PubMed]
- Moret, M.; Ramírez-Tejero, J.A.; Serrano, A.; Ramírez-Yera, E.; Cueva-López, M.D.; Belaj, A.; León, L.; de la Rosa, R.; Bombarely, A.; Luque, F. Identification of Genetic Markers and Genes Putatively Involved in Determining Olive Fruit Weight. Plants 2023, 12, 155. [Google Scholar] [CrossRef] [PubMed]
- Saponari, M.; Boscia, D.; Altamura, G.; Loconsole, G.; Zicca, S.; D’Attoma, G.; Morelli, M.; Palmisano, F.; Saponari, A.; Tavano, D.; et al. Isolation and pathogenicity of Xylella fastidiosa associated to the olive quick decline syndrome in southern Italy. Sci. Rep. 2017, 7, 17723. [Google Scholar] [CrossRef] [PubMed]
- Lia, F.; Zammit-Mangion, M.; Farrugia, C. A First Description of the Phenolic Profile of EVOOs from the Maltese Islands Using SPE and HPLC: Pedo-Climatic Conditions Modulate Genetic Factors. Agriculture 2019, 9, 107. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| N. | Code | Sampling Location | Type | Sampling Point | Use |
|---|---|---|---|---|---|
| 1 | B1 | Bidnija old olive grove | Cultivated | Lower Crown | Oil |
| 2 | B2 | Bidnija old olive grove | Cultivated | Lower Crown | Oil |
| 3 | B3 | Bidnija old olive grove | Cultivated | Lower Crown | Oil |
| 4 | B4 | Bidnija old olive grove | Cultivated | Lower Crown | Oil |
| 5 | B5a | Bidnija old olive grove | Cultivated | Lower Crown | Oil |
| 6 | B5b | Bidnija old olive grove | Cultivated | Lower Crown | Oil |
| 7 | B6a | Bidnija old olive grove | Cultivated | Lower Crown | Oil |
| 8 | B6b | Bidnija old olive grove | Cultivated | Lower Crown | Oil |
| 9 | B7 | Bidnija old olive grove | Cultivated | Lower Crown | Oil |
| 10 | B8 | Bidnija old olive grove | Cultivated | Lower Crown | Oil |
| 11 | B9 | Bidnija old olive grove | Cultivated | Lower Crown | Oil |
| 12 | B10 | Bidnija old olive grove | Cultivated | Lower Crown | Oil |
| 13 | B11 | Bidnija old olive grove | Cultivated | Lower Crown | Oil |
| 14 | B12 | Bidnija old olive grove | Cultivated | Lower Crown | Oil |
| 15 | G1 | Bidnija old olive grove | Cultivated | Upper Crown | Oil |
| 16 | G2 | Bidnija old olive grove | Cultivated | Upper Crown | Oil |
| 17 | G3 | Bidnija old olive grove | Cultivated | Upper Crown | Oil |
| 18 | G4 | Bidnija old olive grove | Cultivated | Upper Crown | Oil |
| 19 | G5 | Bidnija old olive grove | Cultivated | Upper Crown | Oil |
| 20 | G6 | Bidnija old olive grove | Cultivated | Upper Crown | Oil |
| 21 | G7 | Bidnija old olive grove | Cultivated | Upper Crown | Oil |
| 22 | G8 | Bidnija old olive grove | Cultivated | Upper Crown | Oil |
| 23 | G9 | Bidnija old olive grove | Cultivated | Upper Crown | Oil |
| 24 | G10 | Bidnija old olive grove | Cultivated | Upper Crown | Oil |
| 25 | G11 | Bidnija old olive grove | Cultivated | Upper Crown | Oil |
| 26 | G12 | Bidnija old olive grove | Cultivated | Upper Crown | Oil |
| 27 | G14 | Bidnija old olive grove | Cultivated | Upper Crown | Oil |
| 28 | G15 | Bidnija old olive grove | Cultivated | Upper Crown | Oil |
| 29 | G16 | Bidnija old olive grove | Cultivated | Upper Crown | Oil |
| 30 | G17 | Bidnija old olive grove | Cultivated | Upper Crown | Oil |
| 31 | G19 | Bidnija old olive grove | Cultivated | Upper Crown | Oil |
| 32 | G20 | Bidnija old olive grove | Cultivated | Upper Crown | Oil |
| 33 | W.t. 13 | Gozo (south-west coast) | Wild | Crown | - |
| 34 | Cv. 14 | Mellieħa Xaghri | Cultivated | Crown | Oil |
| 35 | Cv. 15 | Mellieħa Xaghri | Cultivated | Crown | Oil |
| 36 | W.t. 16 | St. Martin | Wild | Crown | - |
| 37 | Cv. 17 | St. Martin | Cultivated | Crown | Oil |
| 38 | Cv. 18 | Comino | Cultivated | Crown | Oil |
| 39 | W.t. 19 | Comino | Wild | Crown | - |
| 40 | W.t. 20 | Comino | Wild | Crown | - |
| 41 | W.t. 21 | Comino | Wild | Crown | - |
| 42 | W.t. 22 | Comino | Wild | Crown | - |
| 43 | Malti 23 | Wied Qirda | Cultivated | Crown | Oil |
| 44 | W.t. 24 | Żebbuġ | Wild | Crown | - |
| 45 | Malti 25 | Koronia-Rabat | Cultivated | Crown | Oil |
| 46 | Malti 26 | Żebbuġ | Cultivated | Crown | Oil |
| 47 | Malti 27 | Lija old tree | Cultivated | Crown | Oil |
| 48 | Malti 28 | Unknown | Cultivated | Crown | Oil |
| 49 | Malti 29 | Unknown | Cultivated | Crown | Oil |
| 50 | Malti 30 | Unknown | Cultivated | Crown | Oil |
| 51 | K1 | Old garden | Cultivated | Crown | Oil |
| 52 | K2 | Old garden | Cultivated | Crown | Oil |
| 53 | K3 | Old garden | Cultivated | Crown | Oil |
| 54 | K4 | Old garden | Cultivated | Crown | Oil |
| 55 | K5 | Old garden | Cultivated | Crown | Oil |
| 56 | K6 | Old garden | Cultivated | Crown | Oil |
| 57 | K7 | Old garden | Cultivated | Crown | Oil |
| SSR ID | Bibliographic Reference | Repeat Motif | Ta |
|---|---|---|---|
| DCA03 | Sefc et al. (2000) [21] | (GA)19 | 50 °C |
| DCA05 | Sefc et al. (2000) [21] | (GA)15 | 50 °C |
| DCA09 | Sefc et al. (2000) [21] | (GA)23 | 55 °C |
| DCA13 | Sefc et al. (2000) [21] | (CA)15 | 55 °C |
| DCA17 | Sefc et al. (2000) [21] | (GT)9(AT)7AGATA(GA)38 | 50 °C |
| DCA18 | Sefc et al. (2000) [21] | (CA)4CT(CA)3(GA)19 | 50 °C |
| GAPU71b | Carriero et al. (2002) [22] | GA(AG)6(AAG)8 | 59 °C |
| GAPU101 | Carriero et al. (2002) [22] | (GA)8(G)3(AG)3 | 59 °C |
| EMOL | De la Rosa et al. (2002) [23] | (GA)12 | 50 °C |
| EMO90 | De la Rosa et al. (2002) [23] | (CA)10 | 50 °C |
| Na | Ne | I | Ho | He | uHe | F | |
|---|---|---|---|---|---|---|---|
| Total | 84.0 | 32.8 | |||||
| Mean | 7.6 | 2.9 | 1.323 | 0.765 | 0.651 | 0.657 | −0.167 |
| SE | 0.8 | 0.191 | 0.065 | 0.065 | 0.022 | 0.022 | 0.088 |
| Sample 1 | Sample 2 | LRM |
|---|---|---|
| ALG_Sigoise_2 | FR_Grossanne | 0.50 |
| ALB_Olivo_di_Berat | GRE_Konservolia | 0.50 |
| GRE_Androcarpos | SP_Gordal_sevillana | 0.50 |
| ALG_Aayrouni | LIB_Soury_2 | 0.50 |
| LI_Taggiasca | GRE_Karidolia | 0.50 |
| LIB_Baladi_8 | LIB_Baladi_12 | 0.48 |
| LIB_AbouChawkeh_1 | LIB_AbouChawkeh_2 | 0.47 |
| LIB_Baladi_1 | LIB_Soury | 0.45 |
| LIB_Baladi_8 | LIB_Aayrouni | 0.44 |
| LIB_Del_1 | LIB_Del_2 | 0.43 |
| GRE_Koroneiki_1 | POR_Galega_Grada_de_Serpa | 0.43 |
| LIB_Aayrouni | LIB_Baladi_12 | 0.42 |
| MAL_Bidnija_olive_grove_1 | MAL_Mellieha_xaghri_cv.14 | 0.42 |
| LA_Leccino_Lazio_PR | ALB_Kalinjot_Oval | 0.41 |
| Population | Na | Ne | I | Ho | He | uHe | F |
|---|---|---|---|---|---|---|---|
| Italy | 11.9 | 5.4 | 1.881 | 0.681 | 0.786 | 0.792 | 0.140 |
| Albania | 4.9 | 3.7 | 1.378 | 0.783 | 0.689 | 0.752 | −0.122 |
| France | 6.2 | 4.1 | 1.554 | 0.692 | 0.738 | 0.795 | 0.088 |
| Greece | 4.7 | 3.6 | 1.349 | 0.843 | 0.691 | 0.744 | −0.232 |
| Kurdistan | 6.0 | 4.1 | 1.490 | 0.770 | 0.711 | 0.748 | −0.065 |
| Syria | 5.4 | 4.0 | 1.465 | 0.709 | 0.725 | 0.762 | 0.015 |
| Spain/Portugal | 5.5 | 3.7 | 1.376 | 0.731 | 0.670 | 0.697 | −0.073 |
| Tunisia | 4.6 | 3.0 | 1.157 | 0.531 | 0.571 | 0.594 | 0.107 |
| Malta | 10.4 | 5.8 | 1.922 | 0.735 | 0.801 | 0.819 | 0.085 |
| Lebanon | 4.1 | 2.6 | 1.065 | 0.774 | 0.577 | 0.599 | −0.319 |
| Grand Mean | 6.3 | 4.0 | 1.453 | 0.728 | 0.692 | 0.73 | −0.042 |
| Total | 637 | 402 | - | - | - | - | - |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Miazzi, M.M.; Pasqualone, A.; Zammit-Mangion, M.; Savoia, M.A.; Fanelli, V.; Procino, S.; Gadaleta, S.; Aurelio, F.L.; Montemurro, C. A Glimpse into the Genetic Heritage of the Olive Tree in Malta. Agriculture 2024, 14, 495. https://doi.org/10.3390/agriculture14030495
Miazzi MM, Pasqualone A, Zammit-Mangion M, Savoia MA, Fanelli V, Procino S, Gadaleta S, Aurelio FL, Montemurro C. A Glimpse into the Genetic Heritage of the Olive Tree in Malta. Agriculture. 2024; 14(3):495. https://doi.org/10.3390/agriculture14030495
Chicago/Turabian StyleMiazzi, Monica Marilena, Antonella Pasqualone, Marion Zammit-Mangion, Michele Antonio Savoia, Valentina Fanelli, Silvia Procino, Susanna Gadaleta, Francesco Luigi Aurelio, and Cinzia Montemurro. 2024. "A Glimpse into the Genetic Heritage of the Olive Tree in Malta" Agriculture 14, no. 3: 495. https://doi.org/10.3390/agriculture14030495
APA StyleMiazzi, M. M., Pasqualone, A., Zammit-Mangion, M., Savoia, M. A., Fanelli, V., Procino, S., Gadaleta, S., Aurelio, F. L., & Montemurro, C. (2024). A Glimpse into the Genetic Heritage of the Olive Tree in Malta. Agriculture, 14(3), 495. https://doi.org/10.3390/agriculture14030495