
Phosphorus Bioavailability: A Key Aspect for Conserving this Critical Animal Feed Resource with Reference to Broiler Nutrition

Abstract
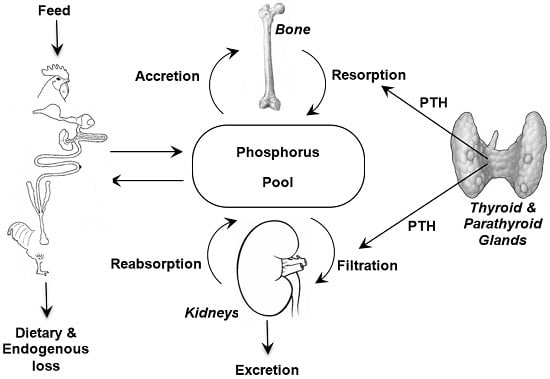
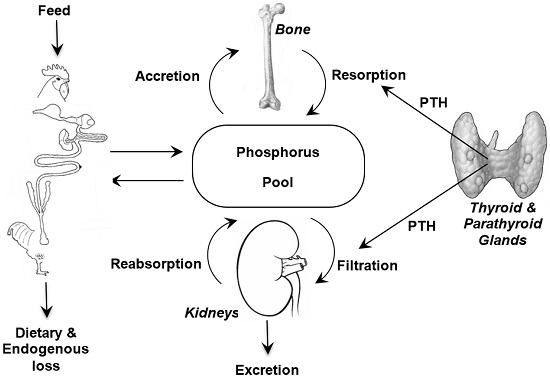
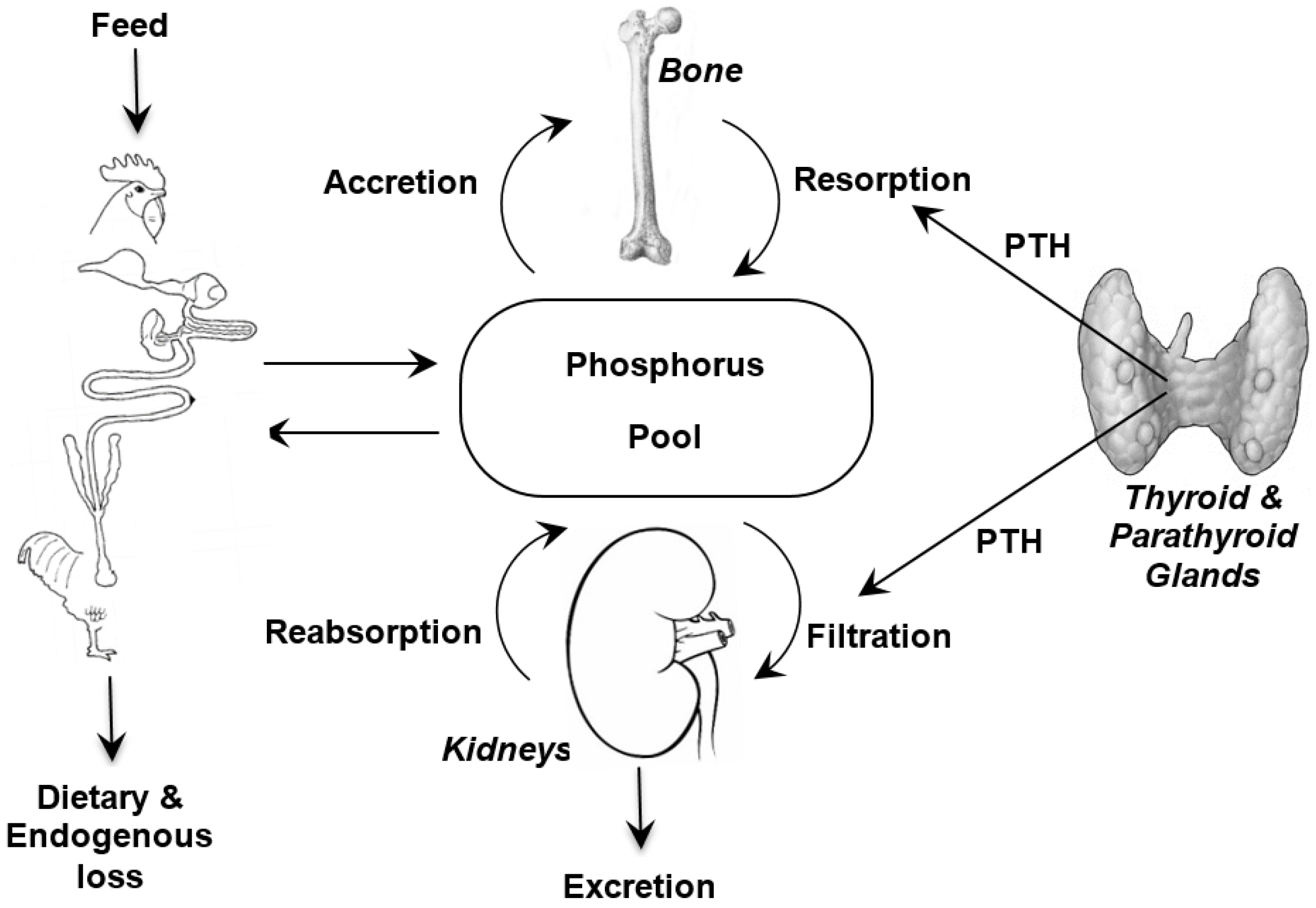
:
1. Introduction
2. Phosphorus Metabolism
3. Phosphorus Requirements
4. Evaluation of Procedures that Determine Bioavailable Phosphorus
4.1. Biologically Determined Values
4.2. Chemically Determined Values
4.3. Ileal Digestibility as an Estimate of Phosphorus Bioavailability
5. Terms Used to Describe Bioavailable Phosphorus
6. Phosphorus Sources and Availability
6.1. Plant Feedstuffs
6.2. Animal Feedstuffs
6.3. Inorganic Phosphorus Sources
7. Factors Affecting Phosphorus Availability
7.1. Phosphorus and Phytate
7.2. Calcium
7.3. Vitamin D
7.4. Phytase
7.5. Birds
8. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Asimov, I. Asimov on Chemistry; Doubleday: Garden City, NY, USA, 1974. [Google Scholar]
- Van der Klis, J.D.; Versteegh, H.A.J. Phosphorus nutrition of poultry. In Recent Developments in Poultry Nutrition 2; Garnsworthy, P.C., Wiseman, J., Eds.; Nottingham University Press: Nottingham, UK, 1999; pp. 309–320. [Google Scholar]
- Shastak, Y.; Witzig, M.; Hartung, K.; Rodehutscord, M. Comparison of retention and prececal digestibility measurements in evaluating mineral phosphorus sources in broilers. Poult. Sci. 2012, 91, 2201–2209. [Google Scholar] [CrossRef] [PubMed]
- Cordel, D.; Drangert, J.; White, S. The story of phosphorus: Global food security and food for thought. Glob. Environ. Chang. 2009, 19, 292–305. [Google Scholar] [CrossRef]
- Neset, T.S.; Cordell, D. Global phosphorus scarcity: Identifying synergies for a sustainable future. J. Sci. Food Agric. 2012, 92, 2–6. [Google Scholar] [CrossRef] [PubMed]
- Devereux, C.; Smart, M.; Kalt, F.P.; Takei, N. Animal feeds: Phosphate supplements. In CEH Marketing Research Report; HIS Chemical: Englewood, CO, USA, 1994. [Google Scholar]
- Waldroup, P.W. Nutritional approaches to reducing phosphorus excretion by poultry. Poult. Sci. 1999, 78, 683–691. [Google Scholar] [CrossRef] [PubMed]
- Van der Klis, J.D.; Versteegh, H.A.J. Phosphorus nutrition in broilers. In Recent Advances in Animal Nutrition; Garnsworty, P.C., Wiseman, J., Haresign, W., Eds.; Nottingham University press: Nottingham, UK, 1996; pp. 71–83. [Google Scholar]
- Cossa, J.; Oloffs, K.; Kluge, H.; Jeroch, H. Investigation into the TP and PP content in different varieties of grain maize. In Proceedings of the 11th European Symposium on Poultry Nutrition; World’s Poultry Science Association: Faaberg, Denmark, 1997; pp. 444–446. [Google Scholar]
- Applegate, T.J.; Angel, R. Feasibility versus Practicality of Phosphorus Reduction in Poultry: Progress and Future Needs. USDA National Agricultural Library: West Lafayette, IN, USA. Available online: http://www.cals.ncsu.edu/waste_mgt/natlcenter/sanantonio/ApplegateandAngel.pdf (accessed on 8 December 2014).
- Schindler, D.W. Evolution of phosphorus limitation in lakes. Sciences 1977, 195, 260–262. [Google Scholar] [CrossRef] [PubMed]
- Sharpley, A.N.; Chapra, S.C.; Wedepohl, R.; Sims, J.T.; Daniel, T.C.; Reddy, K.R. Managing agricultural phosphorus for protection of surface waters: Issues and options. J. Environ. Qual. 1994, 23, 437–451. [Google Scholar] [CrossRef]
- Leske, K.; Coon, C.N. The development of feedstuff retainable phosphorus values for broilers. Poult. Sci. 2002, 81, 1681–1693. [Google Scholar] [CrossRef] [PubMed]
- Angel, R. Calcium and phosphorus requirements in poultry. In Proceedings of the 1st International Phytase Summit, Washington, DC, USA, 28–30 September 2010; pp. 65–71.
- Li, X.; Zhang, D.; Huang, K.H.; Bryden, W.L. Available phosphorus requirement of broilers fed sorghum based diet from day 22 to 49. In Proceedings of the 19th European Symposium on Poultry Nutrition, Potsdam, Germany, 26–29 August 2013; p. 200.
- Li, X.; Zhang, D.; Huang, K.H.; Bryden, W.L. Dietary calcium levels and non-phytate phosphorus requirement of broilers from days 1 to 21. Aust. Poult. Sci. Symp. 2013, 24, 50. [Google Scholar]
- Yang, T.Y.; Setiyawan, E.; Li, X.; Zhang, D.; Bryden, W.L. Dietary calcium levels and non-phytate phosphorus requirement of broilers from days 22–49. In Proceedings of Recent Advances in Animal Nutrition in Australia 2013; University of New England: Armidale, New South Wales, Australia, 2013. [Google Scholar]
- Zhang, D.; Li, X.; Huang, K.H.; Bryden, W.L. Available phosphorus requirement of broilers fed sorghum based diet from day 1 to 21. In Proceedings of the 19th European Symposium on Poultry Nutrition, Potsdam, Germany, 26–29 August 2013; p. 196.
- Li, X.; Zhang, D.; Bryden, W.L. Bioavailability of phosphorus in feedstuffs for broilers. In Final Report for Rural Industries Research and Development Corporation; Rural Industries Research and Development Corporation: Kingston, Australia, 2015. [Google Scholar]
- Klasing, K.C. Comparative Avian Nutrition; CAB International: Wallingford, UK, 1998. [Google Scholar]
- Vitti, D.M.S.S.; Kebreab, E. (Eds.) Phosphorus and Calcium Utilization and Requirements in Farm Animals; CAB International: Wallingford, UK, 2010.
- Yan, F.; Kersey, J.H.; Waldroup, P.W. Phosphorus requirements of broiler chicks three to six weeks of age as influenced by phytase supplementation. Poult. Sci. 2001, 80, 455–459. [Google Scholar] [CrossRef] [PubMed]
- Persia, M.E.; Saylor, W.W. Effects of broiler strain, dietary nonphytate phosphorus, and phytase supplementation on chick performance and tibia ash. J. Appl. Poult. Res. 2006, 15, 72–81. [Google Scholar] [CrossRef]
- Angel, R.; Applegate, T.J.; Christman, M. Effect of dietary non-phytate phosphorus (nPP) on performance and bone measurements in broilers fed on a four-phase feeding system. Poult. Sci. 2000, 79 (Suppl. 1), 21–22. [Google Scholar]
- Angel, R.; Applegate, T.J.; Christman, M.; Mitchell, A.D. Effect of dietary non-phytate phosphorus (nPP) level on broiler performance and bone measurements in the starter and grower phase. Poult. Sci. 2000, 79 (Suppl. 1), 22. [Google Scholar]
- Moran, E.T., Jr.; Todd, M.C. Continuous submarginal phosphorus with broilers and the effect of preslaughter transportation: Carcass defects, further-processing yields, and tibia-femur integrity. Poult. Sci. 1994, 73, 1448–1457. [Google Scholar] [CrossRef] [PubMed]
- Waldroup, P.W.; Kersey, J.H.; Saleh, E.A.; Fritts, C.A.; Yan, F.; Stilborn, H.L.; Crum, R.C., Jr.; Raboy, V. Non-phytate phosphorus requirement and phosphorus excretion of broilers fed diets composed of normal or high available phosphate corn with and without microbial phytase. Poult. Sci. 2000, 79, 1451–1459. [Google Scholar] [CrossRef] [PubMed]
- Yan, F.; Waldroup, P.W. Non-phytate phosphorus requirement and phosphorus excretion of broiler chicks fed diets composed normal or high available phosphate corn as influenced by phytase supplementation and vitamin D source. Int. J. Poult. Sci. 2006, 5, 219–228. [Google Scholar]
- NRC. Nutrient Requirements of Poultry, 9th ed.; National Academy Press: Washington, DC, USA, 1994. [Google Scholar]
- Yan, F.; Kersey, J.H.; Fritts, C.A.; Waldroup, P.W. Phosphorus requirements of broiler chicks six to nine weeks of age as influenced by phytase supplementation. Poult. Sci. 2003, 83, 294–300. [Google Scholar] [CrossRef]
- Dhandu, A.S.; Angel, R. Broiler non-phytin phosphorus requirement in the finisher and withdrawal phases of a commercial four-phase feeding system. Poult. Sci. 2003, 82, 1257–1265. [Google Scholar] [CrossRef] [PubMed]
- Gillis, M.B.; Norris, L.C.; Heuser, G.F. Studies on the biological value of inorganic phosphates. J. Nutr. 1954, 52, 115–125. [Google Scholar] [PubMed]
- Grau, C.R.; Zweigart, P.A. Phosphatic clay as a phosphorus source for chicks. Poult. Sci. 1953, 32, 500–503. [Google Scholar] [CrossRef]
- Fritz, J.C.; Roberts, T.; Bochne, J.W.; Hove, E.L. Factors affecting the chick’s requirement for phosphorus. Poult. Sci. 1969, 48, 307–320. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, M.; Hoshii, H. Improvement of biological assay to determine available phosphorus with growing chicks. J. Poult. Sci. 1977, 14, 33–43. [Google Scholar] [CrossRef]
- Hoshii, H.; Yoshida, M. Comparison of phosphorus availabilities estimated biologically based on toe ash content and carcass phosphorus retention. J. Poult. Sci. 1977, 14, 279–283. [Google Scholar] [CrossRef]
- Potchanacom, M.; Potter, L.M. Biological values of phosphorus from various sources for young turkeys. Poult. Sci. 1987, 66, 505–513. [Google Scholar]
- Potter, L.M. Bio-availability of phosphorus from various phosphates based on body weight and toe ash measurements. Poult. Sci. 1988, 67, 96–102. [Google Scholar] [CrossRef] [PubMed]
- Potter, L.M.; Potchanacom, M.; Ravindran, V.; Kornegay, E.T. Bio-availability of phosphorus in various phosphate sources using body weight and toe ash as response criteria. Poult. Sci. 1995, 74, 813–820. [Google Scholar] [CrossRef] [PubMed]
- Ketels, E.; De Groote, G. The relative bio-availability and the ileal digestibility of phosphorus in mineral and animal sources. In Proceedings of the 18th World’s Poultry Congress, Nagoya, Japan, 4–9 September 1988; pp. 873–875.
- De Groote, G.; Ketels, P.; Seynaeve, P. Relative bio-availability of phosphorus in meat meals as influenced by particle size. In Proceedings of the International Poultry Congress, Istanbul, Turkey, 6–10 September 1991; pp. 42–52.
- De Groote, G. Biological availability of phosphorus in feed phosphates for broilers. In Proceedings of the 4th European Symposium on Poultry Nutrition, Tours, France, 17–20 October 1983; Larbier, M., Ed.; pp. 91–102.
- Sauveur, B. Bio-availability to poultry of plant-origin phosphorus-Methodological criticisms and results. In Proceedings of the 4th European Symposium on Poultry Nutrition, Tours, France, 17–20 October 1983; Larbier, M., Ed.; pp. 103–113.
- Payne, S.G. The Phosphorus Availability of Feed Phosphates in Broilers. MS Thesis, University of Stellenbosh, Stellenbosh, South Africa, 2005. [Google Scholar]
- Edwards, H.M., Jr.; Gillis, M.B. A chromic oxide balance method for determining phosphate availability. Poult. Sci. 1959, 38, 569–574. [Google Scholar] [CrossRef]
- Sibbald, I.R. Measurement of mineral bio-availability: Extension of true metabolisable energy methodology. Poult. Sci. 1982, 61, 485–487. [Google Scholar] [CrossRef]
- Simons, P.C.M.; Versteegh, H.A.J.; Jongbloed, A.W.; Kemme, P.A.; Slump, P. Improvement of phosphorus availability by microbial phytase in broilers and pigs. Br. J. Nutr. 1990, 64, 525–540. [Google Scholar] [CrossRef] [PubMed]
- NRC. Nutrient Requirements of Poultry, 7th ed.; National Academy Press: Washington, DC, USA, 1977. [Google Scholar]
- Ankra-Badu, G.A.; Aggrey, S.E.; Pesti, G.M.; Bakalli, R.I.; Edwards, H.M., Jr. Modelling of parameters affecting phytate phosphorus bioavailability in growing birds. Poult. Sci. 2004, 83, 1083–1088. [Google Scholar] [CrossRef] [PubMed]
- Thompson, L.U.; Yoon, H. Starch digestibility as affected by poly phenols and phytic acid. J. Food Sci. 1984, 49, 1228–1229. [Google Scholar] [CrossRef]
- Walk, C.L.; Bedford, M.R.; McElroy, A.P. Influence of limestone and phytase on broiler performance, gastrointestinal pH, and apparent ileal nutrient digestibility. Poult. Sci. 2011, 91, 1371–1378. [Google Scholar] [CrossRef] [PubMed]
- Ravindran, V.; Bryden, W.L.; Kornegay, E.T. Phytates: Occurrence, bioavailability and implications in poultry nutrition. Poult. Avian Biol. Rev. 1995, 6, 125–143. [Google Scholar]
- Selle, P.H.; Ravindran, V. Microbial phytase in poultry nutrition. Anim. Feed Sci. Technol. 2007, 135, 1–41. [Google Scholar] [CrossRef]
- Angel, R.; Saylor, W.W.; Mitchell, A.D.; Powers, W.; Applegate, T.J. Effect of dietary phosphorus, phytase, and 25-hydroxycholecalciferol on broiler chicken bone mineralization, litter phosphorus, and processing yields. Poult. Sci. 2006, 85, 1200–1211. [Google Scholar] [CrossRef] [PubMed]
- Rodehutscord, M.; Dieckmann, A.; Witzig, M.; Shastak, Y. A note on sampling digesta from the ileum of broilers in phosphorus digestibility studies. Poult. Sci. 2012, 91, 965–971. [Google Scholar] [CrossRef] [PubMed]
- Axe, D. Phosphorus value in ingredient sources examined. Feedstuffs 1998, 70, 17–28. [Google Scholar]
- Savage, J.E.; Yohe, J.M.; Pickett, E.E.; O’Dell, B.L. Zinc metabolism in the growing chick. Tissue concentration and effect of phytate on absorption. Poult. Sci. 1964, 43, 420–426. [Google Scholar] [CrossRef]
- Wise, A. Dietary factors determining the biological activities of phytase. Nutr. Abs. Rev. 1983, 53, 791–806. [Google Scholar]
- Shastak, Y.; Rodehtscord, M. Determination and estimation of phosphorus availability in growing poultry and their historical development. World's Poult. Sci. J. 2013, 69, 569–586. [Google Scholar] [CrossRef]
- Rodehutscord, M.; WPSA (Working Group No. 2: Nutrition of the European Federation of Branches of World Poultry Science Association). Determination of phosphorus availability in poultry. World's Poult. Sci. J. 2013, 69, 687–698. [Google Scholar]
- Allison, P. When Can You Safely Ignore Multicollinearity? 2012. Available online: http://statisticalhorizons.com/multicollinearity (accessed on 4 May 2015).
- Bryden, W.L.; Li, X. Amino acid digestibility and poultry feed formulation: Expression, limitations and application. Rev. Bras. Zootec. 2010, 39 (Suppl.), 279–287. [Google Scholar] [CrossRef]
- Adeola, O.; Applegate, T.J. Phosphorus and calcium equivalency of phytase. In Proceedings of the 1st International Phytase Summit, Washington, DC, USA, 28–30 September 2010; pp. 45–64.
- Adedokun, S.A.; Sands, J.S.; Adeola, O. Determining the equivalent phosphorus released by an Escherichia coli derived phytase in broiler chicks. Can. J. Anim. Sci. 2004, 84, 437–444. [Google Scholar] [CrossRef]
- Dilger, R.N.; Adeola, O. Estimation of true phosphorus digestibility and endogenous phosphorus loss in growing chicks fed conventional and low-phytate soybean meals. Poult. Sci. 2006, 85, 661–668. [Google Scholar] [CrossRef] [PubMed]
- Jendza, J.A.; Dilger, R.N.; Sands, J.S.; Adeola, O. Efficacy and equivalency of an Escherichia coli-derived phytase for replacing inorganic phosphorus in the diets of broiler chickens and young pigs. J. Anim. Sci. 2006, 84, 3364–3374. [Google Scholar] [CrossRef] [PubMed]
- Nyannor, E.K.D.; Adeola, O. Corn expressing an Escherichia coli-derived phytase gene: Comparative evaluation study in broiler chicks. Poult. Sci. 2008, 87, 2015–2022. [Google Scholar] [CrossRef] [PubMed]
- Nelson, T.S. The hydrolysis of phytate phosphorus by chicks and laying hens. Poult. Sci. 1976, 55, 2262–2264. [Google Scholar] [CrossRef] [PubMed]
- Barrier-Guillot, B.; Casado, P.; Maupetit, P.; Jondreville, C.; Gatel, F. Wheat phosphorus availability: 1-In vitro study; Factors affecting endogenous phytasic activity and phytic phosphorus content. J. Sci. Food Agric. 1996, 70, 62–68. [Google Scholar] [CrossRef]
- Li, X.; Zhang, D.; Bryden, W.L. University of Queensland: Gatton, Australia, Unpublished Work. 2015.
- De Groote, G.; Huyghebaert, G. The bio-availability of phosphorus from feed phosphates for broilers as influenced by bio-assay method, dietary Ca-level and feed form. Anim. Feed Sci. Technol. 1997, 69, 329–340. [Google Scholar] [CrossRef]
- Skurray, G.R. The nutritional evaluation of meat meals for poultry. World's Poult. Sci. J. 1974, 30, 129–136. [Google Scholar] [CrossRef]
- Waldroup, P.W. Phosphorus, phytase, and the environment—A retrospective look. Proc. MD Nutr. Conf. Feed Manuf. 2002, 49, 195–202. [Google Scholar]
- Coon, C.N.; Seo, S.; Manangi, M.K. The determination of retainable phosphorus, relative biological availability, and relative biological value of phosphorus sources for broiler. Poult. Sci. 2007, 86, 857–868. [Google Scholar] [CrossRef] [PubMed]
- Gillis, M.B.; Edwards, H.M., Jr.; Young, R.J. Studies on the availability of calcium orthophosphates for chickens and turkeys. J. Nutr. 1962, 78, 155–162. [Google Scholar] [PubMed]
- Lima, F.R.; Mendonca, C.X., Jr.; Alvarez, J.C.; Garzillo, J.M.F.; Ghion, E.; Leal, P.M. Biological evaluations of commercial dicalcium phosphates as sources of available phosphorus for broiler chicks. Poult. Sci. 1997, 76, 1707–1713. [Google Scholar] [CrossRef] [PubMed]
- Sanders, A.M.; Edwards, H.M., Jr.; Roland, G.N. Calcium and phosphorus requirements of the chick as determined response surface analysis. Br. J. Nutr. 1992, 67, 21. [Google Scholar] [CrossRef]
- Ravindran, V.; Cabahug, S.; Ravindran, G.; Selle, P.H.; Bryden, W.L. Response of broiler chickens to microbial phytase supplementation as influenced by dietary phytic acid and non-phytate phosphorus levels. II. Effects on apparent metabolisable energy, nutrient digestibility and nutrient retention. Br. Poult. Sci. 2000, 41, 193–200. [Google Scholar] [CrossRef] [PubMed]
- Bryden, W.L.; Selle, P.H.; Ravindran, V.; Acamovic, T. Phytate: An anti-nutrient factor in animal diets. In Poisonous Plants: Global Research and Solutions; Panter, K.E., Wierenga, T.L., Pfister, J.A., Eds.; CABI Publishing: Wallingford, UK, 2007; pp. 279–284. [Google Scholar]
- Maenz, D.D.; Engle-Schaan, C.M.; Newkirk, R.W.; Classen, H.L. The effect of minerals and mineral chelators on the formation of phytase-resistant and phytase susceptible forms of phytic acid in solution and in a slurry of canola meal. Anim. Feed Sci. Technol. 1999, 81, 177–192. [Google Scholar] [CrossRef]
- Pallauf, J.; Rimbach, G.; Pippig, S.; Schindler, B.; Most, E. Effect of phytase supplementation to a phytate-rich diet based on wheat, barley and soya on the bioavailability of dietary phosphorus, calcium, magnesium, zinc and protein in piglets. Agribiol. Res. 1994, 47, 39–48. [Google Scholar]
- Singh, M.; Krikorian, A.D. Inhibition of trypsin activity in vitro by phytase. J. Agric. Food Chem. 1982, 30, 799–800. [Google Scholar] [CrossRef]
- Deshpande, S.S.; Cheryan, M. Effects of phytate, divalent cations and their implications on alpha-amylase activity. J. Food Sci. 1984, 49, 516–519. [Google Scholar] [CrossRef]
- Liener, I.E. Anti-nutritional factors in legume seeds: State of art. In Recent Advances of Research on Anti-Nutritional Factors in Legume Seeds; Huismen, J., Van Der Poel, T.F.B., Liener, I.E., Eds.; Pudoc: Wageningen, The Netherlands, 1989; pp. 6–13. [Google Scholar]
- Knuckles, B.E.; Betschart, A.A. Effect of phytate and other myo-inositol phosphate esters on alpha-amylase digestion of starch. J. Food Sci. 1987, 52, 719–721. [Google Scholar] [CrossRef]
- Caldwell, R.A. Effect of calcium and phytic acid on the activation of trypsinogen and the stability of trypsin. J. Agric. Food Chem. 1992, 40, 43–46. [Google Scholar] [CrossRef]
- Leeson, S.; Summers, J.D.; Caston, L. Response of brown egg strain layers to dietary calcium or phosphorus. Poult. Sci. 1993, 72, 1510–1514. [Google Scholar] [CrossRef]
- Selle, P.H.; Ravindran, V.; Caldwell, R.A.; Bryden, W.L. Phytate and phytase: Consequences for protein utilisation. Nutr. Res. Rev. 2000, 13, 255–278. [Google Scholar] [CrossRef] [PubMed]
- Champagne, E.T. Effects of pH on mineral-phytate, protein-mineral-phytate and mineral-fiber interactions. Possible consequences of atrophic gastritis on mineral bioavailability from high fiber foods. J. Am. Coll. Nutr. 1988, 38, 199–215. [Google Scholar] [CrossRef]
- Taylor, T.G. Dietary phosphorus and egg shell thickness in the domestic fowl. Br. Poult. Sci. 1965, 6, 79–87. [Google Scholar] [CrossRef] [PubMed]
- Keshavarz, K. The effect of dietary levels of calcium and phosphorus on performance and retention of these nutrients by laying hens. Poult. Sci. 1986, 65, 114–121. [Google Scholar] [CrossRef]
- Keshavarz, K. Phosphorus requirement of laying hens with and without phytase. In Proceedings of the Cornell Nutrition Conference for Feed Manufacturers, Syracuse, NY, USA, 19–21 October 1999; pp. 81–92.
- Hurwitz, S.; Bar, A. Absorption of calcium and phosphorus along the intestinal tract of the laying fowl as influenced by dietary calcium and eggshell formation. J. Nutr. 1965, 86, 433–438. [Google Scholar] [PubMed]
- Guinotte, F.; Gautron, J.; Nys, Y.; Soumarmon, A. Calcium solubilisation and retention in the gastrointestinal tract in chicks (Gallus domesticus) as a function of gastric acid secretion inhibition and of calcium carbonate particle size. Br. J. Nutr. 1995, 73, 125–139. [Google Scholar] [CrossRef] [PubMed]
- Keshavarz, K.; Austic, R.E. Effects of dietary minerals on acid–base balance and eggshell quality in chickens. J. Nutr. 1990, 120, 1360–1369. [Google Scholar] [PubMed]
- Li, X.; Zhang, D.; Bryden, W.L.; Huang, K.H. University of Queensland: Gatton, Australia, Unpublished Work. 2013.
- Selle, P.H.; Ravindran, V.; Bryden, W.L.; Scott, T. Influence of dietary phytate and exogenous phytase on amino acid digestibility in poultry: A review. J. Poult. Sci. 2006, 43, 89–103. [Google Scholar] [CrossRef]
- Shirley, R.B.; Edwards, H.M., Jr. Graded levels of phytase past industry standards improves broiler performance. Poult. Sci. 2003, 82, 671–680. [Google Scholar] [CrossRef] [PubMed]
- Manangi, M.K.; Coon, C. Phytate phosphorus hydrolysis in broilers in response to dietary phytase, calcium, and phosphorus concentrations. Poult. Sci. 2008, 87, 1577–1586. [Google Scholar] [CrossRef] [PubMed]
- Mohammed, A.; Gibney, M.J.; Taylor, T.G. The effects of dietary levels of inorganic phosphorus, calcium and cholecalciferol on the digestion of phytate phosphorus by the chick. Br. Poult. Sci. 1991, 66, 251–259. [Google Scholar]
- Kornegay, E.T. Nutritional, environmental and economic considerations for using phytase in pig and poultry diets. In Nutrient Management of Food Animals to Enhance and Protect the Environment; Kornegay, E.T., Ed.; Lewis Publishers: New York, NY, USA; CRC Press: Boca Raton, FL, USA, 1996; pp. 277–302. [Google Scholar]
- Punna, S.; Roland, D.A., Sr. Influence of supplemental microbial phytase on first cycle laying hens fed phosphorus-deficient diets from day one of age. Poult. Sci. 1999, 78, 1407–1411. [Google Scholar] [CrossRef] [PubMed]
- Tamim, N.M.; Angel, R.; Christman, M. Influence of dietary calcium and phytase on phytate phosphorus hydrolysis in broiler chickens. Poult. Sci. 2004, 83, 1358–1367. [Google Scholar] [CrossRef] [PubMed]
- Augspurger, N.R.; Baker, D.H. Phytase improves dietary calcium utilisation in chicks, and oyster shell, carbonate, citrate, and citrate-malate forms of calcium are equally bioavailable. Nutr. Res. 2004, 24, 293–301. [Google Scholar] [CrossRef]
- Angel, R.; Dhandu, A.S.; Applegate, T.J. Phosphorus requirements of broilers and the impact of exogenous phytases. In Proceedings of the Arkansas Poultry Nutrition Conference; University of Arkansas: Fayetteville, AR, USA, 2002; pp. 1–22. [Google Scholar]
- Applegate, T.J.; Angel, R.; Classen, H.L. Effect of dietary calcium, 25-hydroxycholecalciferol, or bird strain on small intestinal phytase activity in broiler chickens. Poult. Sci. 2003, 82, 1140–1148. [Google Scholar] [CrossRef] [PubMed]
- Nelson, T.S.; Peeler, H.T. The availability of phosphates from single and combined phosphates to chicks. Poult. Sci. 1961, 40, 1321–1327. [Google Scholar] [CrossRef]
- Waldroup, P.W. Bioassays remain necessary to estimate phosphorus, calcium bioavailability. Feedstuffs 1996, 68, 13–23. [Google Scholar]
- Nelson, T.S.; Kirby, L.K. The calcium binding properties of natural phytate in chick diets. Nutr. Rep. Int. 1987, 35, 949–956. [Google Scholar]


{kind=link}
{kind=link}
| AP (g/k) (Bird Age, Day) | |||
|---|---|---|---|
| Criteria of Assessment | 1–21 | 21–42 | Reference |
| Carcass P analysis | 3.7 (day 0–10), 3.0 (day 10–30) | 2.4 (day 30–40) | [2] |
| Bone breaking strength | 3.9 | 3.0 | [13] |
| FCR | 1.63 | [22] | |
| FCR | 3.3–3.9 | [23] | |
| Growth and tibia ash | 3.2–2.8 (day 18–32) | 2.4–1.9 (day 32–42), 1.1 (day 43–49) | [24,25] |
| Growth, bone ash and processing losses | 4.5 | 4.0 | [26] |
| Weight gain | 3.5 (day 0–14) | 3.0 (day 15–49) | [15,18] |
| Weight gain | 3.2–3.4 | [27] | |
| Weight gain | 1.86 | [22] | |
| Weight gain | 3.2–3.5 | [23] | |
| Weight gain | 2.8–2.9 | [28] | |
| Tibia or toe ash | 4.5 | 3.5 | [29] |
| Tibia ash | 3.3 | [22] | |
| Tibia ash | 3.5–3.9 | [23] | |
| Tibia ash | 3.7–3.9 | 3.1 (day 49) | [30] |
| Ingredient | RP (%) | AP (%) | Reference |
|---|---|---|---|
| Low-phosphorus mixed diet | 43 | 29 | [64] 1 |
| Low-phytate soybean meal | 77 | 94 | [65] 2 |
| Conventional soybean meal | 60 | 94 | [65] 2 |
| Low-phosphorus mixed diet | 59 | 52 | [66] 1 |
| Low-phosphorus mixed diet | 45 | 30 | [67] 1 |
| Canola meal | 39 | 66 | [63] 2 |
| Feedstuffs | TP (g/kg) | RP (% TP) * | NPP (g/kg) | PP (g/kg) | PP (% TP) | PP Degradability (%) ** |
|---|---|---|---|---|---|---|
| Beans | 4.9 | 52 | 1.3 | 3.6 | 73.5 | 53 |
| Lupin | 3 | 72 | 1.5 | 1.5 | 50 | 80 |
| Maize | 3 | 29 | 0.7 | 2.3 | 76.7 | 16 |
| Peas | 4.1 | 41 | 1.5 | 2.6 | 63.4 | 23 |
| Rapeseed | 10.9 | 33 | 3.8 | 7.1 | 65.1 | 10 |
| Rice bran | 17.2 | 16 | 3.1 | 14.1 | 82 | 2 |
| Soy bean (heat treated) | 5.5 | 54 | 2 | 3.5 | 63.6 | 49 |
| Soybean meal | 7.1 | 61 | 2.8 | 4.3 | 60.6 | 61 |
| Sunflower meal | 11.9 | 38 | 4.2 | 7.7 | 64.7 | 19 |
| Wheat | 3.4 | 48 | 0.9 | 2.5 | 73.5 | 46 |
| Wheat middlings | 10.8 | 36 | 2.8 | 8 | 74.1 | 26 |
| Animal Feedstuffs | TP (g/kg) | RP (% TP) * |
|---|---|---|
| Bone meal | 76 | 59 |
| Fish meal | 22 | 74 |
| Meat meal | 29 | 65 |
| Meat and bone meal | 60 | 66 |
| Inorganic P Source | TP (g/kg) | RP (% TP) * | Reference |
|---|---|---|---|
| Calcium sodium phosphate | 180 | 59 | [2] * |
| Dicalcium phosphate | 183 | 83 | [74] 1,2 |
| Dicalcium phosphate (anhydrous) | 197 | 55 | [2] * |
| Dicalcium phosphate (hydrous) | 181 | 77 | [2] * |
| Monocalcium phosphate | 226 | 84 | [2] * |
| Monodicalcium phosphate (hydrous) | 213 | 79 | [2] * |
| Monodicalcium phosphate | 203 | 77 | [13] 1 |
| Monodicalcium phosphate | 200 | 80 | [13] 1 |
| Monodicalcium phosphate | 216 | 81 | [13] 1 |
| Monosodium phosphate | 224 | 92 | [2] * |
| Defluorinated phosphate | 182 | 86 | [74] 1,2 |
| Defluorinated phosphate | 179 | 76 | [74] 1,2 |
| Source of Variation | Coefficient of Variation (%) |
|---|---|
| Sampling variation | 5 to 10 |
| Analytical variation | 5 |
| Mixer variation | 5 to 10 |
| Bird utilisation | 16 |
| Ingredient variation (corn and soybean meal) | 8 to 13 |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, X.; Zhang, D.; Yang, T.Y.; Bryden, W.L. Phosphorus Bioavailability: A Key Aspect for Conserving this Critical Animal Feed Resource with Reference to Broiler Nutrition. Agriculture 2016, 6, 25. https://doi.org/10.3390/agriculture6020025
Li X, Zhang D, Yang TY, Bryden WL. Phosphorus Bioavailability: A Key Aspect for Conserving this Critical Animal Feed Resource with Reference to Broiler Nutrition. Agriculture. 2016; 6(2):25. https://doi.org/10.3390/agriculture6020025
Chicago/Turabian StyleLi, Xiuhua, Dagong Zhang, Tsung Y. Yang, and Wayne L. Bryden. 2016. "Phosphorus Bioavailability: A Key Aspect for Conserving this Critical Animal Feed Resource with Reference to Broiler Nutrition" Agriculture 6, no. 2: 25. https://doi.org/10.3390/agriculture6020025
APA StyleLi, X., Zhang, D., Yang, T. Y., & Bryden, W. L. (2016). Phosphorus Bioavailability: A Key Aspect for Conserving this Critical Animal Feed Resource with Reference to Broiler Nutrition. Agriculture, 6(2), 25. https://doi.org/10.3390/agriculture6020025