
The Evaluation of Exogenous Application of Salicylic Acid on Physiological Characteristics, Proline and Essential Oil Content of Chamomile (Matricaria chamomilla L.) under Normal and Heat Stress Conditions

Abstract
:
1. Introduction
2. Material and Methods
2.1. Experimental Site Description
2.2. Plants Growth and Air Temperature
2.3. Salicylic Acid Treatments
2.4. Physiological Characteristics
2.5. Chlorophyll Measurement
2.6. Determination of Free Proline Concentration
2.7. Extraction of Essential Oil
2.8. Data Analysis
3. Results and Discussion
3.1. Plant Height
3.2. Capitol Diameter
3.3. The 1000 Grain Weight
3.4. Fresh Flower Weight
3.5. Dried Flower Weight
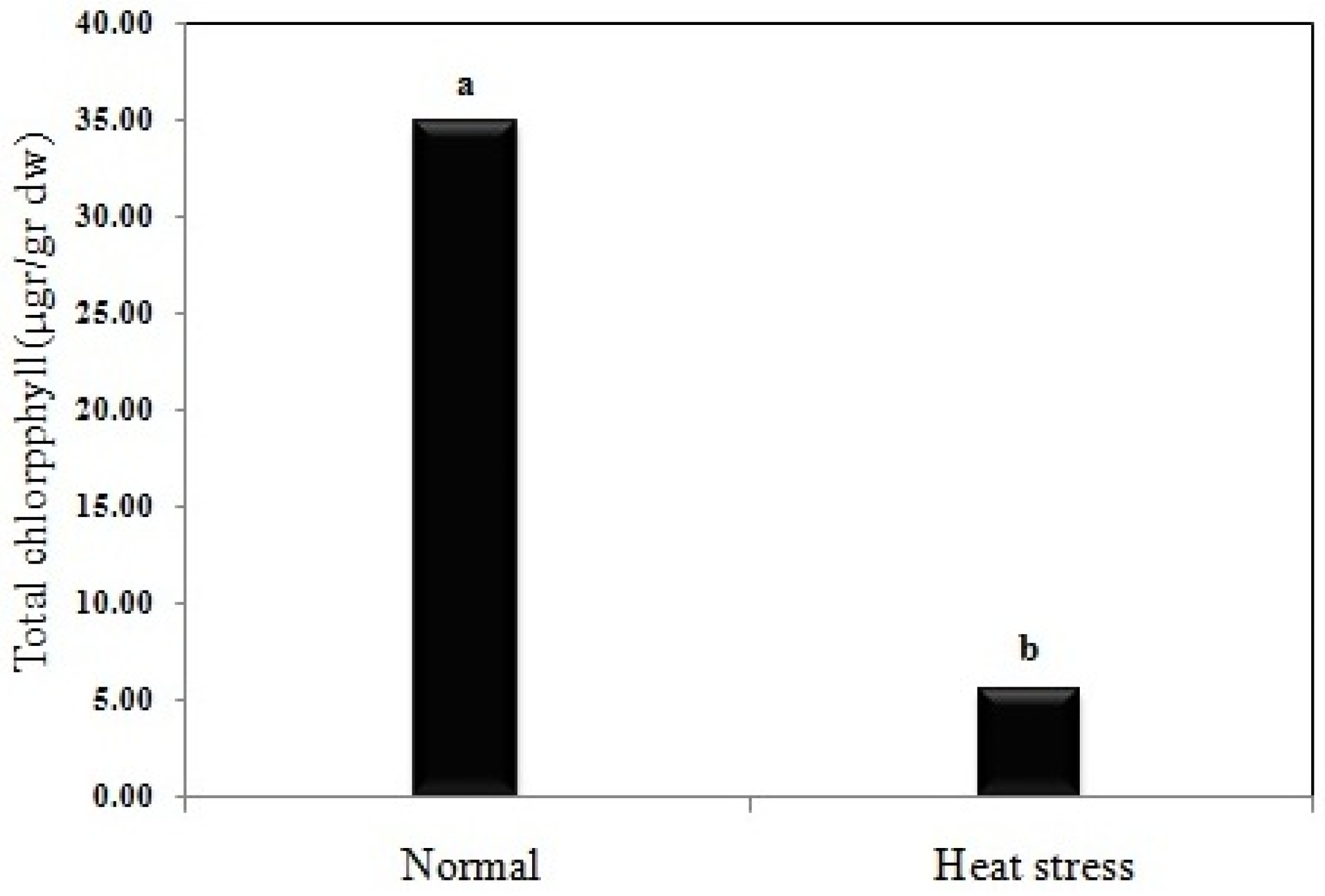
3.6. Total Chlorophyll Content
3.7. Free Proline Concentration
3.8. Essential Oil Content
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Kovacik, J.; Gruz, J.; Backor, M.; Strnad, M.; Repcak, M. Salicylic acid-induced changes to growth and phenolic metabolism in Matricaria chamomilla plants. Plant Cell Rep. 2009. [Google Scholar] [CrossRef]
- Snyman, M.; Cronje, M.J.; Park, A.; Africa, S. Modulation of heat shock factors accompanies salicylic acid-mediated potentiation of Hsp70 in tomato seedlings. J. Exp. Bot. 2008, 59, 2125–2132. [Google Scholar] [CrossRef] [PubMed]
- Larkindale, J.; Huang, B. Effects of abscisic acid, salicylic acid, ethylene and hydrogen peroxide in thermotolerance and recovery for creeping bentgrass. Plant Growth Regul. 2005, 47, 17–28. [Google Scholar] [CrossRef]
- Yousefzadi, M.; Sharifi, M.; Moyano, E.; Palazon, J. Salicylic acid improves podophyllotoxin production in cell cultures of Linum album by increasing the expression of genes related with its biosynthesis. Biotechnol. Lett. 2010, 32, 1739–1743. [Google Scholar] [CrossRef] [PubMed]
- Mustafa, N.; Kim, H.; Choi, Y.; Verpoorte, R. Metabolic changes of salicylic acid-elicited Catharanthus roseus cell suspension cultures monitored by NMR-based metabolomics. Biotechnol. Lett. 2009, 31, 1967–1974. [Google Scholar] [CrossRef] [PubMed]
- Kova, J.; Bac, M. Salicylic acid alleviates NaCl-induced changes in the metabolism of Matricaria chamomilla plants. Ecotoxicology 2009, 18. [Google Scholar] [CrossRef]
- SoloUKi, M.; Mehdikhani, H.; Zeinali, H.; Emamjomeh, A.A. Study of genetic diversity in Chamomile (Matricaria chamomilla) based on morphological traits and molecular markers. Sci. Hortic. J. 2008, 117, 281–287. [Google Scholar] [CrossRef]
- Jeshni, M.G.; MoUSAvinik, M.; Khammari, I.; Rahimi, M. The changes of yield and essential oil components of German Chamomile (Matricaria recutita L.) under application of phosphorus and zinc fertilizers and drought stress conditions. J. Saudi Soc. Agric. Sci. 2015. [Google Scholar] [CrossRef]
- Farhoudi, R.; Lee, D.-J.; Hussain, M. Mild Drought Improves Growth and Flower Oil Productivity of German Chamomile (Matricaria recutita L.). J. Essent. Oil Bear. Plants 2014, 17, 26–31. [Google Scholar] [CrossRef]
- Baghalian, K.; Abdoshah, S.; Khalighi-sigaroodi, F.; Paknejad, F. Physiological and phytochemical response to drought stress of German chamomile (Matricaria recutita L.). Plant Physiol. Biochem. 2011, 49, 201–207. [Google Scholar] [CrossRef] [PubMed]
- Salimi, F.; Shekari, F.; Hamzei, J. Methyl jasmonate improves salinity resistance in German chamomile (Matricaria chamomilla L.) by increasing activity of antioxidant enzymes. Acta Physiol. Plant 2016, 38, 1. [Google Scholar] [CrossRef]
- Rowshan, V.; Bahmanzadegan, A. Effects of salicylic acid on essential oil components in Yarrow (Achillea millefolium Boiss ). Int. J. Basic Sci. Appl. Res. 2013, 2, 347–351. [Google Scholar]
- Rafieiolhossaini, M.; Sodaeizadeh, H.; Adams, A.; De Kimpe, N.; Van Damme, P. Effects of planting date and seedling age on agro-morphological characteristics, essential oil content and composition of German chamomile (Matricaria chamomilla L.) grown in Belgium. Ind. Crops Prod. 2010, 31, 145–152. [Google Scholar]
- Salamon, I. Effect of the internal and external factors on yield and qualitative characteristics of chamomile essential oil. In Proceeding of the I International Symposium on Chamomile Research, Development and Production, Presov, Slovak Republic, 7–10 June 2006; ISHS Acta Horticulturae: Leuven, Belgium.
- Razmjoo, K.; Heydarizadeh, P.; Sabzalian, M.R. Effect of Salinity and Drought Stresses on Growth Parameters and Essential Oil Content of. Int. J. Agric. Biol. 2008, 10, 451–454. [Google Scholar]
- Salamon, I. Chamomile Biodiversity of the Essential oil Qualitative-Quantitative Characteristics; Springer Press: Dordrecht, The Netherlands, 2009; pp. 83–90. [Google Scholar]
- D’Andrea, L. Variation of Morphology, Yield and Essential Oil Components in Common Chamomile (Chamomilla recutita (L.) Rauschert) Cultivars Grown in Southern Italy. J. Herbs Spices. Med. Plants 2002, 37–41. [Google Scholar] [CrossRef]
- Rampino, P.; Lupotto, E.; Marmiroli, N.; Perrotta, C. The effect of heat stress and cadmium ions on the expression of a small Hsp gene in barley and maize. J. Cereal Sci. 2005, 42, 25–31. [Google Scholar]
- Wahid, A.; Gelani, S.; Ashraf, M.; Foolad, M.R. Heat tolerance in plants: An overview. Environ. Exp. Bot. 2007, 61, 199–223. [Google Scholar] [CrossRef]
- Usman, M.G.; Rafii, M.Y.; Ismail, M.R.; Malek, M.A.; Latif, M.A. Expression of Target Gene Hsp70 and Membrane Stability Determine Heat Tolerance in Chili Pepper. J. Am. Soc. Hortic Sci. 2015, 140, 144–150. [Google Scholar]
- Al-whaibi, M.H. Plant heat-shock proteins: A mini review. J. King Saud Univ. Sci. 2011, 23, 139–150. [Google Scholar] [CrossRef]
- Heidari, M.; Sarani, S. Growth, biochemical components and ion content of Chamomile (Matricaria chamomilla L.) under salinity stress and iron deficiency. J. Saudi Soc. Agric. Sci. 2012, 11, 37–42. [Google Scholar] [CrossRef]
- Usman, M.G.; Rafii, M.Y.; Ismail, M.R.; Malek, M.A.; Latif, M.A. Heritability and Genetic Advance among Chili Pepper Genotypes for Heat Tolerance and Morphophysiological Characteristics. Sci. World J. 2014, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Hayat, S.; Masood, A.; Mohammad, Y.; Ahmad, A.; Fariduddin, Q. Growth of Indian mustard (Brassica juncea L.) in response to salicylic acid under high-temperature stress. Brazil. J. Plant Physiol. 2009, 21, 187–195. [Google Scholar] [CrossRef]
- Mohammadi, V.; Bihamta, M.-R.; Zali, A.-A. Evaluation of screening techniques for heat tolerance in wheat. Pakistan J. Biol. Sci. 2007, 10, 887–892. [Google Scholar] [CrossRef]
- Modarresi, M.; Mohammadi, V.; Zali, A.; Mardi, M. Response of Wheat Yield and Yield Related Traits to High Temperature. Cereal Res. Commun. 2010, 38, 23–31. [Google Scholar] [CrossRef]
- Modhej, A.; Naderi, A.; Emam, Y.; Aynehband, A. Effects of post-anthesis heat stress and nitrogen levels on grain yield in wheat (T. durum and T. aestivum) genotypes. Int. J. Plant Prod. 2008, 2, 257–268. [Google Scholar]
- Ayeneh, A.; Van Ginkel, M.; Reynolds, M.P.; Ammar, K. Comparison of leaf, spike, peduncle and canopy temperature depression in wheat under heat stress. Field Crop. Res. 2002, 79, 173–184. [Google Scholar] [CrossRef]
- Midmore, D.J.; Cartwright, P.M.; Fischer, R.A. Wheat in tropical environments. II. Crop growth and grain yield. Field Crop. Res. 1984, 8, 207–227. [Google Scholar] [CrossRef]
- Lichtenthaler, H.K. Chlorophylls and carotenoids: Pigments of photosynthetic biomembranes. Methods Enzymol. 1987, 148, 350–382. [Google Scholar]
- Bates, L.S.; Waldern, R.P.; Teave, I.D. Rapid determination of free proline for water stress studies. Plant Soil 1973, 39, 205–207. [Google Scholar] [CrossRef]
- Kumar, R.R.; Goswami, S.; Sharma, S.K.; Singh, K.; Gadpayle, K.A.; Kumar, N.; Rai, G.K.; Singh, M.; Rai, R.D. Protection against heat stress in wheat involves change in cell membrane stability, antioxidant enzymes, osmolyte, H2O2 and transcript of heat shock protein. Int. J. Plant Physiol. Biochem. 2012, 4, 83–91. [Google Scholar]
- Khalid, K.A.; Teixeira, J.A. Yield, essential oil and pigment content of Calendula officinalis L. flower heads cultivated under salt stress conditions. Sci. Hortic. (Amsterdam) 2010, 126, 297–305. [Google Scholar] [CrossRef]
- SAS Institute. SAS Users Guide: Statistics Version 6th Edn; SAS Institute Inc.: Cary, NC, USA, 1996. [Google Scholar]
- Alexandra, S. German Chamomile (Matricaria Chamomilla L.) Population Morphological and Chemical Diversity; Budapest University: Budapest, Hungary, 2005. [Google Scholar]
- Hussain, M.; Malik, M.A.; Farooq, M.; Khan, M.B.; Akram, M.; Saleem, M.F. Exogenous glycinebetaine and salicylic acid application improves water relations, allometry and quality of hybrid sunflower under water deficit conditions. J. Agron. Crop Sci. 2009, 195, 98–109. [Google Scholar] [CrossRef]
- Farooq, M.; Wahid, A.; Kobayashi, N.; Fujita, D.; Basra, S.M.A. Plant drought stress: Effects, mechanisms and management. Agron. Sustain. Dev. 2009, 29, 185–212. [Google Scholar]
- Circella, G.; De Mastro, G.; Nano, G.M.; D’Andrea, L. Comparison of Chamomile Biotypes (chamomilla recutal Rauschert). In Proceeding of the WOCMAP I-Medicinal and Aromatic Plants Conference, Maastricht, The Netherlands, 1 April 1993; ISHS Acta Horticulturae: Leuven, Belgium.
- Golparvar, A.R.; Pirbalouti, A.G.; Karimi, M. Determination of the effective traits on essence percent and dry flower yield in German chamomile (Matricaria chamomilla L.) populations. J. Med. Plants Res. 2011, 5, 3242–3246. [Google Scholar]
- Taviana, P. Variation for agronomic and essential oil traits among wild populations of Chamomilla recutita (L.) Rausch. from Central Italy. J. Herbs Spices Med. Plant 2001, 9, 1049–6475. [Google Scholar]
- Amani, A.L. Cadmium induced changes in pigment content, ion uptake, proline content and phosphoenolpyruvate carboxylase activity in Triticum aestivum seedlings. Aust. J. Basic Appl. Sci. 2008, 2, 57–62. [Google Scholar]
- Abd El-Baky, H.H.; Hussein, M.M.; El-Baroty, G.S. Algal extracts improve antioxidant defense abilities and salt tolerance of wheat plant irrigated with sea water. Electron. J. Environ. Agric. Food Chem. 2008, 7, 2812–2832. [Google Scholar]
- El-Khallal, S.M.; Hathout, T.A.; Ashour, A.E.A.; Kerrit, A.A.A. Brassinolide and salicylic acid induced growth, biochemical activities and productivity of maize plants grown under salt stress. Res. J. Agric. Biol. Sci. 2009, 5, 380–390. [Google Scholar]
- Wang, L.-J.; Fan, L.; Loescher, W.; Duan, W.; Liu, G.-J.; Cheng, J.-S.; Luo, H.-B.; Li, S.-H. Salicylic acid alleviates decreases in photosynthesis under heat stress and accelerates recovery in grapevine leaves. BMC Plant Biol. 2010, 10, 34. [Google Scholar] [CrossRef] [PubMed]
- Efeoglu, B.; Terzioglu, S. Photosynthetic responses of two wheat varieties to high temperature. EurAsia. J. BioSci. 2009, 3, 97–106. [Google Scholar] [CrossRef]
- Reda, F.; Mandoura, H.M.H. Response of enzymes activities, photosynthetic pigments, proline to low or high temperature stressed wheat plant (Triticum aestivum L.) in the presence or absence of exogenous proline or cysteine. Int. J. Acad. Res. 2011, 3, 108–115. [Google Scholar]
- Ashraf, M.; Harris, P.J.C. Photosynthesis under stressful environments: An overview. Photosynthetica 2013, 51, 163–190. [Google Scholar] [CrossRef]
- Johari-Pireivatlou, M. Effect of soil water stress on yield and proline content of four wheat lines. Afr. J. Biotechnol. 2010, 9, 36–40. [Google Scholar]
- Baghalian, K.; Haghiry, A.; Naghavi, M.R.; Mohammadi, A. Effect of saline irrigation water on agronomical and phytochemical characters of chamomile (Matricaria recutita L.). Sci. Hortic. 2008, 116, 437–441. [Google Scholar] [CrossRef]
- Bita, C.E.; Gerats, T. Plant tolerance to high temperature in a changing environment: Scientific fundamentals and production of heat stress-tolerant crops. Front. Plant Sci. 2013, 4, 1–18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Presibella, M.M.; Villas-Boas, L.D.B.; Belletti, K.M.D.S.; Santos, C.A.D.M.; Weffort-Santos, A.M. Comparison of chemical constituents of Chamomilla recutita (L.) Rauschert essential oil and its anti-chemotactic activity. Braz. Arch. Biol. Technol. 2006, 49, 717–724. [Google Scholar] [CrossRef]
- Flexas, J.; Medrano, H. Drought-inhibition of photosynthesis in C3 plants: Stomatal and non-stomatal limitations revisited. Ann. Bot. 2002, 89, 183–189. [Google Scholar] [CrossRef] [PubMed]
- Priyadarshi, S.; Borse, B.B. Effect of the Environment on Content and Composition of Essential oil in Coriander. Int. J. Sci. Eng. Res. 2014, 5, 57–65. [Google Scholar]
- Kwiatkowski, C.A. Yield and quality of chamomile (Chamomilla recutita (L.) Rausch.) raw material depending on selected foliar sprays and plant spacing. Acta Sci. Pol. Hortorum Cultus 2015, 14, 143–156. [Google Scholar]


{kind=link}
{kind=link}
| Soil Depth (cm) | EC (ds·m−1) | pH | T.N.V * (%) | O.C ** (%) | Ca2+ + Mg2+ | Na+ | Cl− | HCO3− | SO4− | SAR |
|---|---|---|---|---|---|---|---|---|---|---|
| 0–30 | 6.3 | 7.8 | 60 | 0.33 | meq/L | |||||
| 58 | 25 | 20 | 4 | 59 | 4.6 | |||||
| EC (ds·m−1) | pH | HCO3− | Cl− | SO4− | Ca2+ + Mg2+ | Na+ | SAR |
|---|---|---|---|---|---|---|---|
| meq·L−1 | |||||||
| 3.7 | 7.6 | 4.5 | 8.5 | 36 | 37 | 12 | 2.8 |
| Soil Depth (cm) | Bulk Density g·cm−1 | Humidity (%/w) in F.C Status | Humidity (%/w) in P.W.P Status | Particle Density g·cm−1 | The Average of Penetration Rate (cm·h−1) | Moment Penetration Rate (cm·h−1) | Saturation Percent | Texture | Clay Percent |
|---|---|---|---|---|---|---|---|---|---|
| 0–30 | 1.26 | 16.9 | 6.3 | 2.65 | 10.5 | 6.2 | 32 | S.L. | 12 |
| 30–60 | 1.43 | 15.4 | 6.5 | 2.65 | |||||
| 60–90 | 1.43 | 13.3 | 7.2 | 2.65 | |||||
| Month and Year | Average of Temperature (°C) | Average of Relative Humidity (%) | Precipitation (mm) | Average of Sunny Hours (h) | Evaporation (mm) | ||
|---|---|---|---|---|---|---|---|
| Min | Max | Min | Max | ||||
| November 2011 | 17.8 | 30.6 | 27 | 60 | 39.4 | 7.4 | 5.3 |
| December 2011 | 10.2 | 21.4 | 45 | 82 | 62.2 | 8 | 2.9 |
| January 2012 | 10.6 | 22 | 42 | 76 | 32.1 | 6.7 | 2.9 |
| February 2012 | 10.1 | 21 | 39 | 77 | 33.8 | 5.7 | 3.4 |
| March 2012 | 10.9 | 24.1 | 29 | 69 | 22.1 | 6.8 | 5.2 |
| April 2012 | 18.1 | 31.8 | 21 | 64 | 10.2 | 6.6 | 8.8 |
| May 2012 | 25.7 | 40.6 | 12.2 | 34 | 0 | 7.3 | 11.3 |
| June 2012 | 28.2 | 44.1 | 13 | 36 | 0 | 10 | 14.2 |
| Time of Spraying | Environmental Conditions | |
|---|---|---|
| Normal | Heat Stress | |
| 1 | 21 February 2012 | 8 April 2012 |
| 2 | 8 March 2012 | 23 April 2012 |
| 3 | 22 March 2012 | 8 May 2012 |
| Environmental Conditions | ||||||
|---|---|---|---|---|---|---|
| Normal | Heat Stress | |||||
| Harvest Time | Bushehr Cultivar | Bona Cultivar | Bodegold Cultivar | Bushehr Cultivar | Bona Cultivar | Bodegold Cultivar |
| 1 | 27 Mar 2012 | 19 Apr 2012 | 28 Apr 2012 | 22 Apr 2012 | 04 May 2012 | 31 May 2012 |
| 2 | 07 Apr 2012 | 29 Apr 2012 | 08 May 2012 | 03 May 2012 | 14 May 2012 | - |
| 3 | 19 Apr 2012 | 08 May 2012 | 14 May 2012 | 13 May 2012 | 31 May 2012 | - |
| 4 | 29 Apr 2012 | 14 May 2012 | - | 31 May 2012 | - | - |
| 5 | 14 May 2012 | - | - | - | - | - |
| Mean Square (MS) | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| S.O.V | DF | Plant Height (cm) | Capitol Diameter (mm) | Fresh Flower Weight (g) | Dried Flower Weight (g) | 1000 Grain Weight (g) | Total Chlorophyll (µg/g·dw) | Proline (µmol/g·dw) | Essential Oil% (w/w) |
| EC | 1 | 9098.47 ** | 10.51 ** | 241064.95 ** | 14175.22 ** | 0.0024 ** | 19337.71 ** | 15090.63 ** | 0.0008 ns |
| Block | 4 | 19.98 | 0.04 | 6602.1 | 328.69 | 0.00015 | 18.32 | 165.45 | 0.0001 |
| Cult | 2 | 5303.06 ** | 9.22 ** | 171402.77 ** | 9794.01 ** | 0.00022 ns | 130.21 ** | 3423.70 ** | 0.0006 ns |
| SA | 4 | 68.18 ns | 0.24 ns | 18051.77 * | 1080.04 ** | 0.00008 ns | 10.61 ns | 116.75 ns | 0.0004 ns |
| Cult × SA | 8 | 12.66 ns | 0.10 ns | 1750.12 ns | 71.85 ns | 0.0001 ns | 32.82 ns | 222.09 * | 0.0001 ns |
| EC × Cult | 2 | 1406.06 ** | 15.29 ** | 34884.14 ** | 1031.30 * | 0.00064 ** | 29.77 ns | 9070.7 ** | 0.0023 ** |
| EC × SA | 4 | 89.27 ns | 0.15 ns | 28412.84 ** | 1124.1 ** | 0.0001 ns | 1.46 ns | 10.60 ns | 0.0002 ns |
| EC × Cult × SA | 8 | 29.75 ns | 0.17 ns | 6359.83 ns | 323.79 ns | 0.0001 ns | 9.26 ns | 216.25 * | 0.0001 ns |
| Error | 56 | 35.41 | 6.79 | 6656.86 | 290.1 | 0.000085 | 23.91 | 105.41 | 0.0003 |
| C.V | 13.9 | 4.8 | 51.9 | 45.06 | 18.9 | 24.2 | 34.7 | 19.7 | |
| Characteristics | ||||||||
|---|---|---|---|---|---|---|---|---|
| Environmental Conditions | Cultivar | Plant Height (cm) | Capitol Diameter (mm) | Fresh Flower Weight (gr·m−2) | Dried Flower Weight (gr·m−2) | 1000 Grain Weight (g) | Proline (µmol/g·dw) | Essential Oil % (w/w) |
| Normal | Bushehr | 34.08c | 7.18b | 181.75ab | 42.06ab | 0.052a | 24.76b | 0.77d |
| Bona | 60.04a | 6.82bc | 260.42a | 65.63a | 0.056a | 10.96c | 0.82c | |
| Bodegold | 64.41a | 8.94a | 184.42ab | 43.33ab | 0.054a | 14.26c | 1.23a | |
| Heat stress | Bushehr | 22.01d | 6.43c | 79.99bc | 19.05bc | 0.049ab | 15.80c | 0.89b |
| Bona | 47.70b | 7.59ab | 224.22ab | 51.08a | 0.035b | 71.56a | 0.76de | |
| Bodegold | 28.49cd | 6.86bc | 11.85c | 5.60c | 0.046ab | 40.31b | 0.74e | |
| Traits | |||
|---|---|---|---|
| Environmental Conditions | SA Concentration (mg·L−1) | Fresh Flower Weight (g·m−2) | Dried Flower Weight (g·m−2) |
| Normal | 0 | 122.42bc | 30.82b |
| 1 | 287.97a | 67.66a | |
| 10 | 188.53abc | 45.67ab | |
| 25 | 176.31ab | 43.41ab | |
| 100 | 269.09ab | 64.14a | |
| Heat Stress | 0 | 140.10abc | 29.24b |
| 1 | 107.64c | 27.20b | |
| 10 | 96.86c | 23.18b | |
| 25 | 83.09c | 21.45b | |
| 100 | 99.09c | 25.13b | |
| SA Concentration (mg·L−1) | ||||||
|---|---|---|---|---|---|---|
| Environmental conditions | Cultivar | 0 | 1 | 10 | 25 | 100 |
| Normal | Bushehr | 34.73b | 29.30b | 20.30c | 20.06c | 26.56bc |
| Bona | 13.13c | 14.23b | 9.53c | 7.96c | 10.06c | |
| Bodegold | 11.70c | 11.93b | 16.13c | 13.76c | 17.80c | |
| Heat Stress | Bushehr | 13.30c | 22.96b | 18.90c | 11.70c | 12.16c |
| Bona | 91.96a | 64.40a | 67.70a | 75.40a | 58.36a | |
| Bodegold | 27.36bc | 47.80a | 41.50b | 42.06b | 42.83ab | |
| Cultivar | SA Concentration (mg·L−1) | ||||
|---|---|---|---|---|---|
| 0 | 1 | 10 | 25 | 100 | |
| Bushehr | 0.076b | 0.103ab | 0.086b | 0.100ab | 0.103ab |
| Bona | 0.100ab | 0.093ab | 0.093ab | 0.126a | 0.090b |
| Bodegold | 0.086b | 0.076b | 0.093ab | 0.096ab | 0.096ab |
| Cultivar | SA Concentration (mg·L−1) | ||||
|---|---|---|---|---|---|
| 0 | 1 | 10 | 25 | 100 | |
| Bushehr | 0.090a | 0.090a | 0.080a | 0.100a | 0.086a |
| Bona | 0.086a | 0.090a | 0.080a | 0.096a | 0.100a |
| Bodegold | 0.076a | 0.083a | 0.080a | 0.093a | 0.100a |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ghasemi, M.; Modarresi, M.; Babaeian Jelodar, N.; Bagheri, N.; Jamali, A. The Evaluation of Exogenous Application of Salicylic Acid on Physiological Characteristics, Proline and Essential Oil Content of Chamomile (Matricaria chamomilla L.) under Normal and Heat Stress Conditions. Agriculture 2016, 6, 31. https://doi.org/10.3390/agriculture6030031
Ghasemi M, Modarresi M, Babaeian Jelodar N, Bagheri N, Jamali A. The Evaluation of Exogenous Application of Salicylic Acid on Physiological Characteristics, Proline and Essential Oil Content of Chamomile (Matricaria chamomilla L.) under Normal and Heat Stress Conditions. Agriculture. 2016; 6(3):31. https://doi.org/10.3390/agriculture6030031
Chicago/Turabian StyleGhasemi, Mojtaba, Mohammad Modarresi, Nadali Babaeian Jelodar, Nadali Bagheri, and Abbas Jamali. 2016. "The Evaluation of Exogenous Application of Salicylic Acid on Physiological Characteristics, Proline and Essential Oil Content of Chamomile (Matricaria chamomilla L.) under Normal and Heat Stress Conditions" Agriculture 6, no. 3: 31. https://doi.org/10.3390/agriculture6030031
APA StyleGhasemi, M., Modarresi, M., Babaeian Jelodar, N., Bagheri, N., & Jamali, A. (2016). The Evaluation of Exogenous Application of Salicylic Acid on Physiological Characteristics, Proline and Essential Oil Content of Chamomile (Matricaria chamomilla L.) under Normal and Heat Stress Conditions. Agriculture, 6(3), 31. https://doi.org/10.3390/agriculture6030031