
Fertility of Herbivores Consuming Phytoestrogen-containing Medicago and Trifolium Species

Abstract
:
1. Introduction
2. Phytoestrogens
2.1. Phytoestrogenic Compounds
- Isoflavones (e.g., genistein, daidzein, glycitein, formononetin, and puerarin) are primarily found in soybean (Glycine max), chickpeas (Cicer arietinum), and some clovers, most notably subterranean (sub.) clover (Trifolium subterraneum) and red clover (T. pratense) but also white clover (Trifolium repens) in which, more importantly, coumestans may also be present [5]. Amongst the Trifolium (clovers), 14 of 100 species examined by Francis et al. (1967) [6] were found to have contents comparable with sub. clover. Isoflavones in sub. clover are mainly found in the leaf tissue; the availability of suitable carbon substrate is the major determinant of isoflavone content [7,8]. Elevated isoflavone levels are observed in phosphorous deficient, but not potassium deficient, red clover [8] and in phosphorous deficient sub. clover, where leaf concentration of formononetin may quadruple [7,9]. Rossiter (1969) [10] also noted that isoflavone content may double in nitrogen deficient sub clover. Their concentration increases under other stresses such as water deficit, water-logging and disease. Fungal infection can increase isoflavones in sub. clover [11]. While all varieties of isoflavone-containing clover species contain isoflavones, only some varieties have contents that result in economically significant oestrogenic potency. The variety Yarloop may contain an average of 4.8% dry weight as isoflavones [12]. The high potency of the Tallarook variety was noted early using a mice bioassay [13]. Lloyd Davies and Bennett (1962) [14] demonstrated the high potency of the cultivars Yarloop and Dwalganup by measuring the increased weight of the uterus and cervix of virgin and aged ewes. Isoflavones disappear when the clover wilts but are maintained by rapid drying and in well-made hay or silage [2,13,15]. Adams [2] summarised much of the research carried out on sub. clover-induced infertility of sheep in Western Australia.
- Flavones (e.g., luteolin, apigenin, querectin, chrysin, kaempherol, and wogonin). Numerous flavones are found in a diverse range of species including lucerne and white clover [16].
- Stilbenes (e.g., resveratrol) [16]
- Lignans (e.g., lariciresinol, matairesinol, and secoisolariciresinol can be metabolized by gastro-intestinal bacteria to the oestrogenic ‘mammalian lignins’ enterolactone and enterodiol) [16].
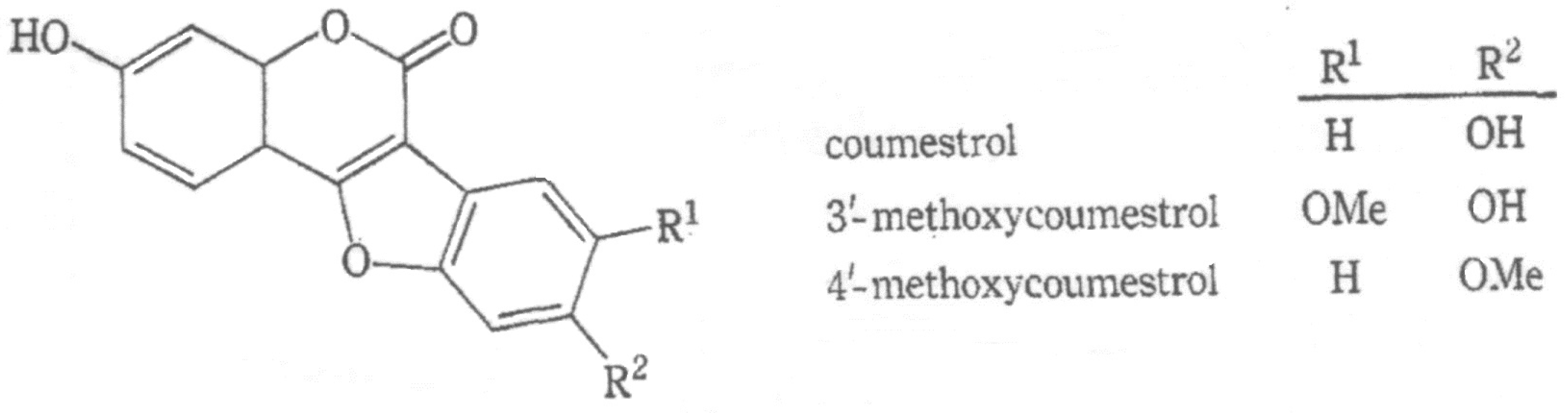
- Coumestans: At least 27 coumestans have been described [17,18], many of which have two or three synonyms. Coumestrol [syn 7’12’ dihydroxy coumestan] has been found in 58 plants [2], especially legumes, (e.g., lucerne), other perennial Medicago (e.g., M. falcata), annual ‘medics’ (Medicago spp.), peas (Pisum sativum), soybean, limabeans (Phaseolus lunatus), pinto beans (P. vulgaris), and some clovers (e.g., white clover [5,19,20] and strawberry clover (T. fragiferum) [6]). The relative abundance of particular coumestans and flavones that white clover produces in the field varies considerably depending on which plant pathogen stimulates their production [21,22]. The coumestrol content in white clover was considered insufficient to explain the oestrogenic activity it was responsible for [5,20]; other phytoestrogens may be involved. Although it is the most commonly measured coumestan, coumestrol’s concentration is likely to underestimate the oestrogenicity of the plant. Other coumestans include 4’-methoxy-coumestrol, 3’-methoxycoumestrol [syn 7,12-dihydroxy-11-methoxycoumestan], 11,12 dimethoxy-7-hydroxy coumestan, coumestrol dimethyl ether [syn 7’12’ dimethoxy coumestans], aureol, lucernol, medicagol, repensol, sativol, trifoliol, wairol, wedelactone [syn 7-methoxy-5,11,12-trihydroxy-coumestan], and wedelolactone [17,18].
2.2. Genetic Influence on Phytoestrogen Production
2.3. Anabolic Effects of Phytoestrogens
3. Phytoestrogen Effects on Fertility
3.1. Oestrogenic Activity
3.2. Mode of Action
3.3. Clinical Signs
4. Coumestans
4.1. Coumestans in Annual Medics
4.2. Coumestans in Lucerne
4.3. Coumestrol Production Stimuli
5. Coumestan Infertility
5.1 Sheep
5.1.1. Suppression of Oestrus
5.1.2. Ovulation
5.1.3. Advice to Industry
5.1.4. Tolerance Levels
5.2. Cattle
5.3. Horses
6. Conclusions
Acknowledgements
Author Contributions
Conflicts of Interest
References
- Irwin, J.A.G.; Lloyd, D.L.; Lowe, K.F. Lucerne biology and genetic improvement—an analysis of past activities and future goals in Australia. Aust. J. Agric. Res. 2001, 52, 699–712. [Google Scholar] [CrossRef]
- Adams, N.R. Phytoestrogens. In Toxicants of Plant Origin; Cheeke, P.R., Ed.; CRC Press: Boca Raton, FL, USA, 1989; Volume 4, pp. 25–32. [Google Scholar]
- Mostrom, M.; Evans, T.J. Phytoestrogens. In Reproductive and Developmental Toxicology; Gupta, R.C., Ed.; Academic Press—Medical: Amsterdam, The Netherlands, 2011; pp. 707–722. [Google Scholar]
- Clarke, D.; Wiseman, H. Phytoestrogens. In Bioactive Compounds in Foods; Gilbert, J., Senyuva, H., Eds.; John Wiley and Sons: Oxford, UK, 2009; pp. 173–198. [Google Scholar]
- Nykänen-Kurki, P.; Saloniemi, H.; Kallela, K.; Saastamoinen, I. Phyto-oestrogen content and oestrogenic effect of white clover. In White clover in Europe: State of the art; Frame, J., Ed.; Food and Agriculture Organisation of the United Nations: Rome, Italy, 1993. [Google Scholar]
- Francis, C.M.; Millington, A.J.; Bailey, E.T. The distribution of oestrogenic isoflavones in the genus Trifolium. Aust. J. Agric. Res. 1967, 18, 47–54. [Google Scholar] [CrossRef]
- Rossiter, R.C.; Beck, A.B. Physiological and ecological studies on the oestrogenic isoflavones in subterranean clover (T. subterraneum L.). II. Effects of phosphate supply. Aust. J. Agric. Res. 1966, 17, 447–456. [Google Scholar] [CrossRef]
- Butler, G.W.; Steemers, M.A.T.; Wong, E. The effect of nitrogen, phosphorus and potassium supply on the isoflavone content of the leaves of red clover. N. Z. J. Agric. Res. 1967, 10, 312–315. [Google Scholar] [CrossRef]
- Alexander, G.; Rossiter, R.C. The effects of fertilizer treatments on the oestrogenic potency of Trifolium subterraneum. Aust. J. Agric. Res. 1951, 3, 24–28. [Google Scholar] [CrossRef]
- Rossiter, R.C. Physiological and ecological studies on the oestrogenic isoflavones in subterranean clover (T. subterraneum L.) VII. Effects of nitrogen supply. Aust. J. Agric. Res. 1969, 20, 1043–1051. [Google Scholar] [CrossRef]
- Parbery, D.G.; Gardner, W.K.; Golebiowski, T. Stimulation of isoflavonoid content in subterranean clover by infection with a fungus. J. Aust. Inst. Agric. Sci. 1984, 50, 113–116. [Google Scholar]
- Rossiter, R.C. Evaluation of genotypes of subterranean clover in a Mediterranean environment; a personal and historical account. In Proceedings of the 5th Australian Agronomy Conference, Perth, Australia, 25–30 September 1989; pp. 13–23. Available online: http://www.regional.org.au/au/asa/1989/donald-oration/p.htm (accessed on 24 May 2016).
- Bickoff, E.M.; Booth, A.N.; Livingston, A.L.; Hendrikson, A.P. Estrogenic activity of fresh and dried red and subterranean clovers. J. Anim. Sci. 1961, 20, 133–136. [Google Scholar]
- Lloyd Davies, H.; Bennett, D. Studies on the oestrogenic activity of subterranean clover on sheep reproduction in south Western Australia. Aust. J. Agric. Res. 1962, 13, 1030–1040. [Google Scholar] [CrossRef]
- Shehata, M.N.; Hassan, A.B.; El-Shazly, K. Identification of the Oestrogenic Isoflavones in Fresh and Fermented Berseem Clover (Trifolium alexandrinum). Aust. J. Agric. Res. 1982, 33, 951–956. [Google Scholar] [CrossRef]
- Moutsatsou, P. The spectrum of phytoestrogens in nature: Our knowledge is expanding. Hormones 2007, 6, 173–193. [Google Scholar] [PubMed]
- The Netherlands Metabolomics Institute. 2015. Available online: http://www.metabolomicscentre.nl/ (accessed on 24 May 2016).
- Bickoff, E.M.; Spencer, R.R.; Witt, S.C.; Knuckles, B.E. Studies on the Chemical and Biological Properties of Coumestrol and Related Compounds; Technical Bulletin No. 1408; US Department of Agriculture - Agricultural Research Service: Washington, DC, USA, 1969; p. 95.
- Sanger, V.L.; Bell, D.S. Estrogenic activity in green forage crops and its effects on breeding ewes. J. Am. Vet. Med. Assoc. 1959, 134, 237–239. [Google Scholar] [PubMed]
- Saba, N.; Drane, H.M.; Herbert, C.N.; Newton, J.E.; Betts, J.E. Effect of disease on the oestrogenic activity and coumestrol content of white clover. J. Agric. Sci. 1972, 78, 471–475. [Google Scholar] [CrossRef]
- Wong, E.; Latch, G.C.M. Effect of fungal diseases on the phenolic contents of white clover. N. Z. J. Agric. Res. 1971, 14, 633–638. [Google Scholar] [CrossRef]
- Wong, E.; Flux, D.S.; Latch, G.C.M. The oestrogenic activity of white clover. N. Z. J. Agric. Res. 1971, 14, 639–645. [Google Scholar] [CrossRef]
- Shutt, D.A.; Braden, A.W.; Lindner, H.R. Plasma coumestrol levels in sheep following administration of synthetic coumestrol or ingestion of Medic hay (Medicago littoralis). Aust. J. Agric. Res. 1969, 20, 65–69. [Google Scholar] [CrossRef]
- Kelly, R.W.; Adams, N.R.; Lindsay, D.R. Effect of coumestans on reproduction in the ewe. Aust. J. Agric. Res. 1976, 27, 253–259. [Google Scholar] [CrossRef]
- Kelly, R.W.; Lindsay, D.R. Plasma coumestrol levels and cervical mucus responses in ewes ingesting coumestan-rich feeds. Aust. J. Agric. Res. 1978, 29, 115–121. [Google Scholar] [CrossRef]
- Coop, I.E. Depression of lambing performance from mating on lucerne. Proc. N. Z. Soc. Anim. Prod. 1977, 37, 149–151. [Google Scholar]
- Scales, G.H.; Moss, R.A.; Kelly, R.W. Reproductive performance of ewes mated on lucerne. Proc. N. Z. Soc. Anim. Prod. 1977, 37, 152–157. [Google Scholar]
- McLeod, B.J. An Investigation into the Mechanisms Involved in the Depression of Ovulation Rates in Ewes Grazing Oestrogenic Lucerne. Master’s Thesis, Massey University, Palmerston North, New Zealand, 1978; p. 103. Available online: http://mro.massey.ac.nz/bitstream/handle/10179/5010/02_whole.pdf?sequence=2 (accessed on 24 May 2016). [Google Scholar]
- Ferreira-Dias, G.; Botelho, M.; Zagrajczuk, A.; Rebordao, M.R.; Galvao, A.M.; Pinto Bravo, P.; Piotrowski-Tomala, K.; Szostek, A.Z.; Wiczkowski, W.; Piskul, M.; et al. Coumestrol and its metabolite in mares’ plasma after ingestion of phytoestrogen-rich plants: Potent endocrine disruptors inducing infertility. Theriogenology 2013, 80, 684–692. [Google Scholar] [CrossRef] [PubMed]
- Morley, F.H.W. A New Look at Phytoestrogens. In Proceedings New South Wales Divion, Australian Veterinary Association Annual General Meeting; Australian Veterinary Association: Sydney, Australia, 1966; p. 4. [Google Scholar]
- Reed, K.F.M.; Moore, D.D. A preliminary survey of zearalenone and other mycotoxins in Australian silage and pasture. Anim. Prod. Sci. 2009, 49, 696–703. [Google Scholar] [CrossRef]
- Cocks, P.S.; Craig, A.D.; Kenyon, R.V. Evolution of subterranean clover in South Australia. II. Change in genetic composition of a mixed population after 19 years grazing on a commercial farm. Aust. J. Agric. Res. 1982, 33, 679–695. [Google Scholar] [CrossRef]
- Day, H.R. A Comparative Production Study of Three Strains of Sub Clover; 1965–66 Report of the Kangaroo Island Research Centre; South Australian Department of Agriculture: Adelaide, Australia, 1966.
- Hume, I.D.; Somers, M.; McKeown, N.R. Nutritive evaluation of two strains of mature subterranean clover. Aust. J. Exp. Agric. Anim. Husb. 1968, 8, 295–300. [Google Scholar] [CrossRef]
- Stewart, G. Variation in Sub Clover in Southern New South Wales; Small Seeds Bulletin, New South Wales Department of Agriculture: Sydney, Australia, 1983.
- Dear, B.S.; Sandral, G.A. Subterranean Clover in NSW—Identification and Use; Agfact P2.5.16; New South Wales Agriculture: Dubbo, Australia, 1997; p. 36.
- Francis, C.M.; Millington, A.J. Varietal variation in the isoflavone content of subterranean clover: Its estimation by a microtechnique. Aust. J. Agric. Res. 1965, 16, 557–564. [Google Scholar] [CrossRef]
- Rumball, W.; Keogh, G.; Miller, J.E.; Claydon, R.B. Grasslands G27 red clover (Trifolium pratense L.). N. Z. J. Agric. Res. 1997, 40, 369–372. [Google Scholar] [CrossRef]
- Reed, K.F.M. A review of legume‑based vs. nitrogen‑fertilized pasture systems for sheep and beef cattle. In Forage Evaluation: Concepts and Techniques; Wheeler, J.L., Mochrie, R.D., Eds.; CSIRO and American Forage and Grassland Council: Melbourne, Australia, 1981; pp. 401–417. [Google Scholar]
- National Research Council. Hormonal Relationships and Applications in the Production of Meats, Milk and Eggs; National Research Council: Washington, DC, USA, 1959. [Google Scholar]
- Oldfield, J.E.; Fox, C.W.; Bahn, A.V.; Bickoff, E.M.; Kohler, G.O. Coumestrol in alfalfa as a factor in growth and carcass quality in lambs. J. Anim. Sci. 1966, 25, 167–174. [Google Scholar] [PubMed]
- Trenkle, A.; Burroughs, W. Physiological effects of estrogens in animal feeds with emphasis on growth in ruminants. In Nutrition and Drug Interrelationships; Hathcock, J.N., Coon, J., Eds.; Academic Press: New York, NY, USA, 1978; pp. 577–611. [Google Scholar]
- McClure, K.E.; Solomon, M.B.; Parrett, N.A.; Van Keuren, R.W. Growth and tissue accretion in lambs fed concentrate in drylot, grazed on alfalfa or ryegrass at weaning or after backgrounding on ryegrass. J. Anim. Sci. 1995, 73, 3437–3444. [Google Scholar] [PubMed]
- Moorby, J.M.; Fraser, M.D.; Theobald, V.J.; Wood, J.D.; Haresign, W. The effect of red clover formononetin content on liveweight gain, carcass characteristics and muscle equol content of finishing lambs. Anim. Sci. 2004, 79, 303–313. [Google Scholar]
- Pace, V.; Carbone, K.; Spirito, F.; Iacurto, M.; Terzano, M.G.; Verna, M. The effects of subterranean clover phytoestrogens on sheep growth, reproduction and carcass characteristics. Meat Sci. 2006, 74, 616–622. [Google Scholar] [CrossRef] [PubMed]
- Nogowski, L. Effects of phytoestrogen-coumestrol on lipid and carbohydrate metabolism in young ovariectomised rats may be independent of its estrogenicity. J. Nut. Biochem. 1999, 10, 664–669. [Google Scholar] [CrossRef]
- Little, D.A. Assessment of several pasture species, particularly tropical legumes, for oestrogenic activity. Aust. J. Agric. Res. 1976, 27, 681–686. [Google Scholar] [CrossRef]
- Boué, S.M.; Wiese, T.E.; Nehls, S.; Burow, M.E.; Elliott, S.; Carter-Wientjes, C.H.; Shih, B.Y.; McLachlan, J.A.; Cleveland, T.E. Evaluation of the estrogenic effects of legume extracts containing phytoestrogens. J. Agric. Food Chem. 2003, 51, 2193–2199. [Google Scholar] [CrossRef] [PubMed]
- Whitten, P.L.; Russell, E.; Naftolin, F. Effects of a normal, human-concentration, phytoestrogen diet on rat uterine growth. Steroids 1992, 57, 98–106. [Google Scholar] [CrossRef]
- Jefferson, W.N.; Patisaul, H.B.; Williams, C.J. Reproductive Consequences of Developmental Phytoestrogen Exposure. Reproduction 2012, 143, 247–260. [Google Scholar] [CrossRef] [PubMed]
- Smith, J.F.; Jagusch, K.T.; Brunswick, L.F.C.; Kelly, R.W. Coumestans in lucerne and ovulation in ewes. N. Z. J. Agric. Res. 1979, 22, 411–416. [Google Scholar] [CrossRef]
- Braden, A.W.H.; Hart, N.K.; Lamberton, J.A. The oestrogenic activity and metabolism of certain isoflavones in sheep. Aust. J. Agric. Res. 1967, 18, 335–348. [Google Scholar] [CrossRef]
- Cox, R.I.; Braden, A.W. The metabolism and physiological effects of phytoestrogens in Livestock. Proc. Aust. Soc. Anim. Prod. 1974, 10, 122–129. [Google Scholar]
- Folman, Y.; Pope, G.S. The interaction in the immature mouse of potent oestrogens with coumestrol, genistein and other utero-vagino trophic compounds of low potency. J. Endocrinol. 1966, 34, 215–225. [Google Scholar] [CrossRef] [PubMed]
- Woclawek-Potocka, I.; Acosta, T.J.; Korzekwa, A.; Bah, M.M.; Shibaya, M.; Okuda, K.; Skarzynski, D.J. Phytoestrogens modulate prostaglandin production in bovine endometrium: Cell type specificity and intracellular mechanisms. Exp. Biol. Med. 2005, 230, 326–333. [Google Scholar]
- Bickoff, E.M.; Livingston, A.L.; Hendrikson, A.P. and Booth, A.N. Relative potencies of several estrogen-like compounds found in forages. J. Agric. Food Chem. 1962, 10, 410–412. [Google Scholar] [CrossRef]
- Woclawek-Potocka, Y.; Okuda, K.; Acosta, T.J.; Korzekwa, A.; Pilawski, W.; Skarzynskia, D.J. Phytoestrogen metabolites are much more active than phytoestrogens themselves in increasing prostaglandin F2α synthesis via prostaglandin F2α synthase-like stimulation in bovine endometrium. Prostagladins Other Lipid Mediat. 2005, 78, 202–217. [Google Scholar] [CrossRef] [PubMed]
- Lindner, H.R. Study of the fate of phyto-oestrogens in the sheep by determination of isoflavones and coumestrol in the plasma and adipose tissue. Aust. J. Agric. Res. 1967, 18, 305–333. [Google Scholar] [CrossRef]
- Morley, S.A.; Murray, J.A. Effects of Body Condition Score on the Reproductive Physiology of the Broodmare: A Review. J. Eq. Vet. Sci. 2014, 34, 842–853. [Google Scholar] [CrossRef]
- Ashworth, C.J.; Toma, L.M.; Hunter, M.G. Nutritional effects on oocyte and embryo development in mammals: Implications for reproductive efficiency and environmental sustainability. Phil. Trans. R. Soc. London B Biol. Sci. 2009, 364, 3351–3361. [Google Scholar] [CrossRef] [PubMed]
- Bennetts, H.W.; Underwood, E.J.; Shier, F.L. A specific breeding problem of sheep on subterranean clover pastures in Western Australia. Aust. Vet. J. 1946, 22, 2–12. [Google Scholar] [CrossRef] [PubMed]
- Adams, N.R. Permanent infertility in ewes exposed to plant oestrogens. Aust. Vet. J. 1990, 67, 197–201. [Google Scholar] [CrossRef] [PubMed]
- Pugh, D.G. Theriogenology of Sheep and Goats. In Sheep and Goat Medicine; Schrefer, J.A., Ed.; Saunders: Philadelphia, PA, USA, 2002; p. 173. [Google Scholar]
- Croker, K.P.; Nichols, P.G.H.; Barbetti, M.J.; Adams, N.R. Sheep Infertility from Pasture Legumes; Farmnote. 41/2005; Western Australian Department of Agriculture: Perth, Australia, 2007.
- Dohle, L. Sheep in Clover May Give You Less Than You Bargained. Natural Resources, South Australia. 2015. Availlable online: http://www.naturalresources.sa.gov.au/kangarooisland/news/150928-sheep-in-clover (accessed on 6 November 2015). [Google Scholar]
- Lightfoot, R.J.; Wroth, R.H. The mechanism of temporary infertility in ewes grazing oestrogenic subterranean clover prior to and during joining. Proc. Aust. Soc. Anim. Prod. 1974, 10, 130–134. [Google Scholar]
- Lightfoot, R.J. A look at recommendations for the control of infertility due to clover disease in sheep. Proc. Aust. Soc. Anim. Prod. 1974, 10, 113–121. [Google Scholar]
- Austin, A.R.; Aston, K.; Drane, H.M.; Saba, N. The fertility of heifers consuming red clover silage. Grass Forage Sci. 1982, 37, 101–106. [Google Scholar] [CrossRef]
- Lundh, T.J.-O.; Pettersson, H.I.; Martinsson, K.A. Comparative levels of free and conjugated plant oestrogens in blood plasma of sheep and cattle fed oestrogenic silage. J. Agric. Food Chem. 1990, 38, 1530–1534. [Google Scholar] [CrossRef]
- Adams, N.R.; Sanders, M.R. Development of uterus-like redifferentiation in the cervix of the ewe after exposure to estradiol-17 beta. Biol. Reprod. 1993, 48, 357–362. [Google Scholar] [CrossRef] [PubMed]
- Lightfoot, R.J.; Croker, K.P.; Neil, H.G. Failure of sperm transport in relation to ewe infertility following prolonged grazing on oestrogenic pastures. Aust. J. Agric. Res. 1967, 18, 755–765. [Google Scholar] [CrossRef]
- Adams, N.R.; Sanders, M.R.; Ritar, A.J. Oestrogenic damage and reduced fertility in ewe flocks in south Western Australia. Aust. J. Agric. Res. 1988, 39, 71. [Google Scholar] [CrossRef]
- Newton, J.E.; Betts, J.E. The effects of red clover (Trifolium pratense var. Redhead), white clover (Trifolium repens var. S 100) or perennial ryegrass (Lolium perenne var. S 23) on the reproductive performance of sheep. J. Agric. Sci. 1973, 80, 323–327. [Google Scholar] [CrossRef]
- Shackell, G.H.; Wylie, J.G.; Kelly, R.W. Effects of prolonged exposure of ewes to oestrogenic pasture. 2. Occurrence of abnormalities of the external genitalia and altered mating performance. N. Z. J. Agric. Res. 1993, 36, 459–464. [Google Scholar] [CrossRef]
- Morley, F.H.W.; Bennett, D.; Axelsen, A. The effect of stilboestrol administered during an autumn mating on reproduction in Merino sheep. Aust. J. Agric. Res. 1963, 14, 660–669. [Google Scholar] [CrossRef]
- Morley, F.H.W.; Axelsen, A.; Bennett, D. Effects of grazing red clover during the mating season on ewe fertility. Proc. Aust. Soc. Anim. Prod. 1964, 5, 58–61. [Google Scholar]
- Adams, N.R. Detection of the effects of phytoestrogens on sheep and cattle. J. Anim. Sci. 1995, 73, 1509–1515. [Google Scholar] [PubMed]
- Lloyd Davies, H.; Rossiter, R.C.; Maller, R. The effects of different cultivars of subterranean clover (T. subterraneum L.) on sheep reproduction in the South-west of Western Australia. Aust. J. Agric. Res. 1970, 21, 359–369. [Google Scholar] [CrossRef]
- Lindsay, D.R.; Francis, C.M. Cervical mucus measurement in ovariectomised ewes as a bioassay of synthetic and phyto-oestrogens. Aust. J. Agric. Res. 1968, 19, 1069–1076. [Google Scholar] [CrossRef]
- Farkas, G.L.; Kiraly, Z. Role of phenolic compounds in the physiology of plant diseases and disease resistance. J. Phytopathol. 1962, 44, 105–150. [Google Scholar] [CrossRef]
- Bhattacharya, A.; Sood, P.; Citovsky, V. The roles of plant phenolics in defence and communication during Agrobacterium and Rhizobium infection. Mol. Plant Pathol. 2010, 11, 705–719. [Google Scholar] [CrossRef] [PubMed]
- Howie, J.; Ballard, R.; Peck, D.; Hill, J. A Strand Medic Resistant to Powdery Mildew for the Eyre Peninsula and Mallee; Minnepa Field Report; South Australia Research and Development Institute: Urrbrae, Australia, 2015; p. 4.
- Reid, R.L. Manual of Australian Agriculture, 5th ed.Australian Institute of Agricultural Science: Sydney, Australia, 1990.
- Nair, R.M.; Howie, J.; Ballard, R.; Hutton, R.; Charman, N.; Preston, C. Genetic improvement of strand medic (Medicago littoralis Rohde ex Lois.) for Australian farming systems. In Proceedings of the 12th Australian Agronomy Conference, Brisbane, Australia, 26–30 September 2004; Available online: http://www.regional.org.au/au/asa/2004/symposia/6/3/716_nairrm.htm#TopOfPage (accessed on 26 July 2016).
- Francis, C.M.; Millington, A.J. Wether bioassay of annual pasture legumes IV. The oestrogenic activity of annual medic pastures. Aust. J. Agric. Res. 1965, 16, 927–935. [Google Scholar] [CrossRef]
- Croker, K.P.; Barbetti, M.J.; Nichols, P.G.H. Incidence of coumestrol in medic pastures in Western Australia. Proc. Aust. Soc. Anim. Prod. 1994, 20, 416. [Google Scholar]
- Barbetti, M.J. Relative resistance, associated yield losses and phyto-oestrogen production from fungal foliar diseases in new and old annual Medicago cultivars. Aust. J. Agric. Res. 1995, 46, 441–450. [Google Scholar] [CrossRef]
- Francis, C.M.; Millington, A.J. The presence of methylated coumestans in annual Medicago species: Response to a fungal pathogen. Aust. J. Agric. Res. 1971, 22, 75–80. [Google Scholar] [CrossRef]
- Millington, A.J.; Francis, C.M.; McKeown, N.R. Wether bioassay of annual pasture legumes I. Oestrogenic activity in Medicago tribuloides var. Cyprus, relative to 4 strains of Trifolium subterraneum. Aust. J. Agric. Res. 1964, 15, 520–526. [Google Scholar] [CrossRef]
- Loper, G.M. Accumulation of coumestrol in barrel medic (Medicago littoralis) [sic]. Crop Sci. 1968, 8, 317–319. [Google Scholar] [CrossRef]
- Sormunen-Cristian, R.; Taponen, S.; Saastamoien, I.; Mela, T.; Saloniemi, H. Yellow-flowered lucerne: Properties and influence on performance and reproduction of ewes. Agric. Food Sci. Fin. 1998, 7, 437–446. [Google Scholar]
- Loper, G.M.; Hanson, C.H. Influence of controlled environmental factors and two foliar pathogens on coumestrol content of alfalfa. Crop Sci. 1964, 4, 480–482. [Google Scholar] [CrossRef]
- Loper, G.M.; Hanson, C.H.; Graham, J.H. Coumestrol content of alfalfa as affected by selection for resistance to foliar diseases. Crop Sci. 1967, 7, 189–192. [Google Scholar] [CrossRef]
- Hanson, C.H.; Loper, G.M.; Kohler, G.O.; Bickoff, E.M.; Taylor, K.W.; Kehr, W.R.; Stanford, E.H.; Dudley, J.W.; Pedersen, M.W.; Sorensen, E.L.; et al. Variation in Coumestrol Content of Alfalfa as Related to Location, Variety, Cutting, Year, Stage of Growth and Disease; Agricultural Technical Bulletin 1333; US Department of Agriculture: Washington, DC, USA, 1965; p. 72.
- Sherwood, R.T.; Olah, A.F.; Oleson, W.H.; Jones, E.E. Effect of disease and injury on accumulation of a flavonoid estrogen, coumestrol, in Alfalfa. J. Phytopathnol. 1970, 60, 684–688. [Google Scholar] [CrossRef]
- Kain, W.M.; Biggs, D.R. Effect of pea aphid and blue-green lucerne aphid (Acyrthosiphon spp.) on coumestrol levels in herbage of Lucerne (Medicago sativa). N. Z. J. Agric. Res. 1980, 23, 563–568. [Google Scholar] [CrossRef]
- Hall, D.G. Coumestrol content of lucerne in NSW. Proc. Aust. Soc. Anim. Prod. 1984, 15, 689. [Google Scholar]
- Hall, D.G.; Waterhouse, D.B. Coumestrol content of lucerne in the central west and Hunter valley of NSW. In Proceedings of the 3rd Australian Agronomy Conference, Hobart, Australia, January–February 1985; Available online: http://www.regional.org.au/au/asa/1985/concurrent/sub-clover-pasture-legumes/p-11.htm (accessed on 28 May 2016).
- Le Bars, J.; Le Bars, P. Recent acute and sub-acute mycotoxicoses recognized in France. Vet. Res. 1996, 27, 383–394. [Google Scholar] [PubMed]
- Seguin, P.; Zheng, W.; Souleimanov, A. Alfalfa Phytoestrogen Content: Impact of Plant Maturity and Herbage Components. J. Agron. Crop Sci. 2004, 190, 211–217. [Google Scholar] [CrossRef]
- Cheng, E.; Story, C.D.; Payne, L.C.; Yoder, L.; Burroughs, W. Detection of oestrogenic substances in alfalfa and clover hays fed to fattening lambs. J. Anim. Sci. 1953, 12, 507–514. [Google Scholar]
- Jansen, M.A.; Gaba, V.; Greenberg, B.M. Higher plants and UV-B radiation: Balancing damage, repair and acclimation. Trends Plant Sci. 1998, 3, 131–135. [Google Scholar] [CrossRef]
- Swinny, E.E.; Ryan, K.G. Red Clover Phytoestrogens: UV-B Radiation Increases Isoflavone Yield, and Postharvest Drying Methods Change the Glucoside Conjugate Profiles. J. Agric. Food Chem. 2005, 53, 8273–8278. [Google Scholar] [CrossRef] [PubMed]
- Seguin, P.; Wenju, Z. Phytoestrogen content of alfalfa cultivars grown in eastern Canada. J. Sci. Food Agric. 2006, 86, 765–771. [Google Scholar] [CrossRef]
- McLean, J.W. Lucerne and phytoestrogens. In The Lucerne Crop; Langer, R.H.M., Ed.; Reed: Wellington, New Zealand, 1967; pp. 276–282. [Google Scholar]
- Hanson, C.H. Varietal Improvement of Alfalfa as related to Product Quality and Utility. In Proceedings of 9th Technical Alfalfa Conference, Lincoln, NE, USA, 17 November 1965; pp. 5–9.
- Bickoff, E.M.; Livingston, A.L.; Booth, A.N.; Thomson, C.R.; Hollowell, E.A.; Beinhart, E.G. Some variation in estrogenic activity in fresh and dried white clover clones and the Ladino variety. J. Anim. Sci. 1960, 19, 1143–1149. [Google Scholar]
- Stob, M.; Davis, R.L.; Andrews, F.N. Strain differences in the oestrogenicity of alfalfa varieties. J. Anim. Sci. 1957, 16, 850–853. [Google Scholar]
- Weitzkin, G.; Marinov, U.; Roderig, H. Coumestrol in some alfalfa varieties in Israel. Refuah Vet. 1968, 25, 112–116. [Google Scholar]
- Purves, R.G.; Hood, N.D.; Dunbier, M.W. The effect of cutting management and fungicide application on coumestans levels in three lucerne cultivars. In Proceedings of 34th New Zealand Weed Pest Control Conference; New Zealand Weed Pest Control Society: Blenheim, New Zealand, 1981; pp. 25–28. [Google Scholar]
- Bickoff, E.M. Oestrogenic Constituents of Forage Plants; Commonwealth Bureau of Pastures and Field Crops: Hurley, Berkshire, UK, 1968; Review Series 1; p. 39. [Google Scholar]
- Shemesh, M.; Ayalon, N.; Lindner, H.R. Coumestrol and 4’-O-methyl coumestrol in alfalfa grown in Northern Israel. Possible effects of a foliar pathogen (Pseudopeziza medicaginis). Refuah Vet. 1969, 26, 1–7. [Google Scholar]
- Loper, G.M. Effect of aphid infestation on the coumestrol content of alfalfa varieties. Crop Sci. 1968, 8, 104–106. [Google Scholar] [CrossRef]
- Lloyd Davies, H. Limitations to livestock production associated with phytoestrogens and bloat. In Temperate Pastures, Their Production, Use and Management; Wheeler, J.L., Pearson, C.J., Robards, G.E., Eds.; CSIRO: Melbourne, Australia, 1987; pp. 446–456. [Google Scholar]
- Ruttle, J.L.; Goret, E.A. Effect of alfalfa on ewe fertility. J. Anim. Sci. 1968, 27, 1104. [Google Scholar]
- Morgan, W.C.; Parbery, D.G. Depressed Fodder quality and increased oestrogenic activity of lucerne infected with Pseudopeziza medicaginis. Aust. J. Agric. Res. 1980, 31, 1103–1110. [Google Scholar] [CrossRef]
- Saloniemi, H.; Wahala, K.; Nykanen-Kurki, P.; Kallela, K.; Saastamoinen, I. Phytoestrogen Content and Estrogenic Effect of Legume Fodder. Exp. Biol. Med. 1995, 208, 13–17. [Google Scholar] [CrossRef]
- Kara, E.; Tsanakalis, F.; Papaioannidou, P. Is coumestrol a natural beneficial compound or a hazardous agent? Available online: http://www.frontiersin.org/10.3389/conf.fphar.2010.60.00136/event_abstract (accessed on 24 May 2016).
- Cantero, A.; Sancha, J.L.; Flores, J.M.; Rodriguez, A.; Gonzalez, T. Histopathological changes in the reproductive organs of Manchego ewes grazing on lucerne. J. Vet. Med. Ser. A-Zentralblatt. 1996, 43, 325–330. [Google Scholar] [CrossRef]
- Oldfield, J.E.; Fox, C.W.; Bickoff, E.M. Effects of estrogenic activity in alfalfa on growing lambs. J. Anim. Sci. 1960, 19, 1281. [Google Scholar]
- Valderrábanoa, J.; Ramóna, J.P.; Barberána, M. Morphological alterations in the reproductive organs of ewe lambs reared on lucerne. Anim. Prod. 1988, 47, 271–274. [Google Scholar] [CrossRef]
- Morley, F.H.W.; Axelsen, A.; Bennett, D. Recovery of normal fertility after grazing on red clover. Aust. Vet. J. 1966, 42, 278. [Google Scholar] [CrossRef]
- Newsome, F.E.; Kitts, W.D. Effects of alfalfa consumption on estrogen levels in ewes. Can. J. Anim. Sci. 1977, 57, 531–535. [Google Scholar] [CrossRef]
- Amin, A.; Buratovich, M. The Anti-Cancer Charm of Flavonoids: A Cup-of-Tea Will Do! Recent Patents Anti-Cancer Drug Discov. 2007, 2. [Google Scholar] [CrossRef]
- Blomquist, C.H.; Lima, P.H.; Hotchkiss, J.R. Inhibition of 3α-hydroxysteoid dehydogenase (3α-HSD) activity of human lung microsomes by genistein, daidzein, coumestrol and C18-, C19- and C21 hydroxysteroids and ketosteroids. Steroids 2005, 70, 507–514. [Google Scholar] [CrossRef] [PubMed]
- Hettle, J.A.; Kitts, W.D. Effects of phytoestrogenic alfalfa consumption on plasma LH levels in cycling ewes. Anim. Reprod. Sci. 1983, 6, 233–238. [Google Scholar] [CrossRef]
- Engle, P.H.; Bell, D.S.; Davis, R.R. The effect of Ladino clover, birdsfoot trefoil and bluegrass pasture on the rate of conception among ewes. J. Anim. Sci. 1957, 16, 703–710. [Google Scholar]
- Coop, I.E.; Clark, V.R. The Reproductive Performance of Ewes mated on Lucerne. N. Z. J. Agric. Res. 1960, 3, 922–933. [Google Scholar] [CrossRef]
- Donnelly, J.R.; Morley, F.H.W.; McKinney, G.T. The productivity of breeding ewes grazing on lucerne or grass and clover pastures on the tablelands of Southern Australia. 1. Reproduction. Aust. J. Agric. Res. 1982, 33, 1085–1097. [Google Scholar] [CrossRef]
- Robertson, S.M.; Clayton, E.H.; King, B.J.; Knott, S.; Morgan, B.; Friend, M.A. Lucerne pasture ad libitum, after day 7 post insemination, may increase embryo mortality in ewes. Proc. Aust. Soc. Anim. Prod. 2014, 30, 66. [Google Scholar]
- King, B.J.; Robertson, S.M.; Wilkins, J.F.; Friend, M.A. Short-term grazing of lucerne and chicory increases ovulation rate in synchronised Merino ewes. J. Reprod. Sci. 2010, 121, 242–248. [Google Scholar] [CrossRef] [PubMed]
- Robertson, S.M.; Clayton, E.H.; Friend, M.A. Reproductive performance of ewes grazing lucerne during different periods around mating. Anim. Reprod. Sci. 2015, 162, 62–72. [Google Scholar] [CrossRef] [PubMed]
- Robertson, S.M.; Clayton, E.H.; Morgan, B.; Friend, M.A. Reproductive performance in ewes fed varying levels of cut lucerne pasture around conception. Anim. Reprod. Sci. 2015, 158, 75–85. [Google Scholar] [CrossRef] [PubMed]
- Cox, R.I. Plant oestrogens affecting livestock in Australia. In Effects of Poisonous Plants on Livestock; Keeler, R., Van Kempen, K.R., James, L.F., Eds.; Academic Press: New York, NY, USA, 1978; pp. 451–464. [Google Scholar]
- Venter, J.L.; Greyling, J.P.C. Effect of different periods of flushing and synchronized mating on body weight, blood glucose and reproductive performance in spring-mated ewes. Small Ruminant Res. 1994, 13, 257–261. [Google Scholar] [CrossRef]
- Ferguson, J.D.; Chalupa, W. Impact of Protein Nutrition on Reproduction in Dairy Cows. J. Dairy Sci. 1989, 72, 746–766. [Google Scholar] [CrossRef]
- Berardinelli, J.G.; Weng, J.; Burfening, P.J.; Adair, R. Effect of excess degradable intake protein on early embryonic development, ovarian steroids, and blood urea nitrogen on days 2, 3, 4, and 5 of the estrous cycle in mature ewes. J. Anim. Sci. 2001, 79, 193–199. [Google Scholar] [PubMed]
- Thomson, N.; Jagusch, K.T. Effect of lambing date on the utilisation of grass/clover and lucerne pastures during mating. Proc. N. Z. Soc. Anim. Prod. 1976, 36, 184–189. [Google Scholar]
- Jagusch, K.T.; Smith, J.F.; Kelly, R.W. Effect of feeding lucerne during mating on the fertility of ewes. Proc. N. Z. Nutr. Soc. 1977, 2, 161. [Google Scholar]
- Smith, J.F.; Jagusch, K.T.; Brunswick, L.E.C.; McGowan, L.T. The effect of lucerne feeding on the ovulation rate in ewes. Proc. N. Z. Soc. Anim. Prod. 1980, 40, 44–49. [Google Scholar]
- Bennett, D.; Morley, F.H.W.; Axelsen, A. Bioassay responses of ewes to legume swards. II Uterine weight results from swards. Aust. J. Agric. Res. 1967, 18, 495–500. [Google Scholar] [CrossRef]
- Shemesh, M.; Lindner, H.R.; Ayalon, N. Affinity of rabbit uterine oestradiol receptor for phyto-oestrogens and its use in a competitive binding radioassay for plasma coumestrol. J. Reprod. Fert. 1972, 29, 1–9. [Google Scholar] [CrossRef]
- Foltin, E. Observations on bovine infertility in the Jordan Valley. Refuah Vet. 1959, 16, 193. [Google Scholar]
- Adler, D.; Trainin, J.H. Apparent hyper-oestrogenism in dairy cows. Refuah Vet. 1959, 16, 40. [Google Scholar]
- Adler, D.; Trainin, J.H. Diet and bovine fertility. Vet. Rec. 1960, 72, 1171–1172. [Google Scholar]
- Adler, D.; Trainin, J.H. The apparent effect of alfalfa on the reproductive performance of dairy cattle. In Proceedings of the 4th International Congress of Animal Reproduction, The Hague, The Netherlands, 5–9 June 1961; pp. 451–456.
- Lotan, E.; Adler, J.H. Early effects of excessive alfalfa feeding on bovine fertility. Refuah Vet. 1966, 23, 110–112. [Google Scholar]
- Romero, R.C.M.; Tarrago, C.M.R.; Munoz, M.R.; Arista, R.R.; Rosado, G.R. Oestrogenic syndrome in dairy cows by alfalfa consumption with large amounts of coumestrol. Veterinaria Mexico. 1997, 28, 25–30. [Google Scholar]
- Lookhart, G.L. Analysis of coumestrol, a plant estrogen, in animal feeds by HPLC. J. Agric. Food Chem. 1980, 28, 666–667. [Google Scholar] [CrossRef] [PubMed]
- Shemesh, M.; Shore, L.S. Effects of Environmental Estrogens on Reproductive Parameters in Domestic Animals. Israel J. Vet. Med. 2012, 67, 6–10. [Google Scholar]
- Botelho, M.; Rebordao, M.R.; Balvao, A.M.; Pinto Bravo, P.; Piotrowska-Tomala, K.; Szostek, A.Z.; Wiczkowski, W.; Piscula, M.; Skarzynski, D.J.; Fradinho, M.J.; et al. Phytoestrogen coumestrol and its metabolites in mares’ plasma after clover mixed pasture and alfalfa pellets ingestion. In Forages and Grazing in Horse Nutrition; Saastamoinen, M., Fradinho, M.N.J., Santos, A.S., Miraglia, N., Eds.; European Federation of Animal Science: Wageningen, The Netherlands, 2012; Scientific Series 132; pp. 49–53. [Google Scholar]
- McCue, P.M.; Squires, E.L. Persistent anovulatory follicles in the mare. Theriogenology 2002, 58, 541–543. [Google Scholar]
- Mottershead, J. Anovulatory Hemorrhagic Follicles. 2007. Available online: http://www.equine-reproduction.com/articles/AHF.shtml (accessed on 24 May 2016).
- Pierson, R.A. Folliculogenesis and ovulation. In Equine Reproduction; McKinnon, A.O., Voss, J.L., Eds.; Lea and Febiger: Philadelphia, PA, USA, 1993; pp. 161–171. [Google Scholar]
- McKinnon, A.O. Ovarian abnormalities. In Equine Diagnostic Ultrasonography; Ranaten, N.W., McKinnon, A.O., Eds.; Williams and Wilkins: Baltimore, MD, USA, 1997; pp. 233–251. [Google Scholar]
- Ginther, O.J. Reproductive Biology of the Mare: Basic and Applied Aspects; Equiservices: Cross Plains, WI, USA, 1992. [Google Scholar]


{kind=link}
{kind=link}
| Cultivar, if Known | Stage of Growth | Plant Material Analysed | Disease 1/Pest Status | Coumestrol (mg/kg, DM Basis) 3 | Reference |
|---|---|---|---|---|---|
| Medicago truncatula | |||||
| Cyprus | fully podded | leaf | not stated | 40–180 | [89] |
| commercial | early podding | leaf | not stated | 45–210 | |
| Hannaford 2 | mature | stem | healthy | 218 (576) | [88] |
| Cyprus 2 | mature | stem | healthy | 232 (382) | |
| Jemalong 2 | mature | stem | healthy | 132 (272) | |
| Caliph | mature | stem & pod (no seed) | Phoma, Lepto., Pseudo. | 230 | [87] |
| Caliph | mature | stem & pod (no seed) | +fungicide | 80 | |
| Cyprus | mature | stem & pod (no seed) | Phoma, Lepto., Pseudo. | 350 | |
| Cyprus | mature | stem & pod (no seed) | +fungicide | 110 | |
| not stated | green, late spring | tops | Phoma (low) rated <3 (0–10) | 0–100 | [86] (Western Australia survey) |
| not stated | dry, mature | tops | Phoma (low) rated 0 (0–10) | 0–15 | |
| Medicago polymorpha | |||||
| var. denticulata | early burr | leaf | healthy | 9 (16) | [88] |
| var. denticulata | early burr | leaf | Uromyces, low | 24 (38) | |
| var. denticulata | early burr | leaf | Uromyces, medium | 39 (50) | |
| var. denticulata | early burr | leaf | Uromyces, high | 50 (80) | |
| Circle Valley | mature | stem | Phoma, Lepto., Pseudo. | 570 | [87] |
| Circle Valley | mature | stem | +fungicide | 470 | |
| Santiago | mature | stem | Phoma, Lepto., Pseudo. | 470 | |
| Santiago | mature | stem | +fungicide | 290 | |
| not stated | green, late spring | tops | Phoma (low) rated 0–10 | 0–250 | [86] (Western Australia survey) |
| not stated | dry (mature) | tops | Phoma (low) rated 0–10 | 0–800 | |
| not stated | dry (mature) | burr | Phoma (low) rated 0–10 | 0–200 | |
| not stated | dry (mature) | burr | Phoma (high) rated 0–10 | 100–200 | |
| Medicago littoralis | |||||
| Harbinger | emergence | cotlydons | not stated | 1 | [83] |
| Harbinger | dry (mature) | senescent | not stated | 335 | |
| Harbinger | hay | tops | not stated | 400 (810) | [23] |
| Harbinger 2 | mature | stem | healthy | 528 (737) | [88] |
| mature | leaf | spoted/physiogenic | 2362 | [90] | |
| Angel, Herald, Jaguar | mature | top 5 nodes—including pods | Phoma, Erysiphe | 1050 | [82] |
| Experimental var. | mature | top 5 nodes—including pods | Phoma, Erysiphe | 240 | |
| Angel, Herald, Jaguar | mature | top 5 nodes—including pods | Erysiphe | 383 | |
| Experimental var. | mature | top 5 nodes—including pods | Erysiphe | 37 | |
| Medicago murex | |||||
| Zodiac | mature | stem | Phoma, Lepto., Pseudo. | 880 | [87] |
| Zodiac | mature | stem | +fungicide | 270 | |
| Medicago scutellata | |||||
| wild type 2 | mature | stem | healthy | 66 (122) | [88] |
| Medicago sphaerocarpos | |||||
| Orion | mature | stem | Phoma, Lepto., Pseudo. | 470 | [87] |
| Orion | mature | stem | +fungicide | 170 | |
| Cultivar, if Known | Stage of Growth | Plant Material Analysed | Disease 1/Pest Status | Coumestrol (mg/kg, DM Basis) | Reference |
|---|---|---|---|---|---|
| Medicago falcata | |||||
| Karlu | summer | tops | not studied | 0–60 | [91] |
| Karlu | summer | silage | not studied | 26–44 | |
| Medicago sativa | |||||
| Ranger | leaves, 2 or >lesion/leaflet | Pseudo. | 184 | [92] | |
| Ranger | leaves, 1 lesion/leaflet | Pseudo | 40 | ||
| Ranger | leaves, 2 or >lesion/leaflet | Lepto. | 72 | ||
| Ranger | leaves, 1 lesion/leaflet | Lepto. | 29 | ||
| Uromyces | 400 | [93] | |||
| Ranger and others | 6 cuts/2 years | Tops | not stated | 6–429 | [94] (USA survey) |
| not stated | seed pod | not stated | 340–560 | EM Bickoff cited by [94] | |
| Ranger | 1st bud—bloom | leaf | diseased | 62–92 | [94] |
| Ranger | 1st bud—bloom | leaf | +fungicide | 29–35 | |
| Ranger | 1st bud—bloom | stem | diseased | 32–112 | |
| Ranger | 1st bud—bloom | stem | +fungicide | 30 | |
| Buffalo | prebud | leaf and stem | Phoma | 182–219 | |
| Buffalo | prebud | leaf and stem | healthy | 0–1 | |
| Buffalo | ¼ bloom | leaf and stem | Phoma | 60–74 | |
| Vernal | prebud | leaf and stem | Pseudopeziza | 33–48 | |
| Vernal | prebud | leaf and stem | healthy | 0 | |
| Clone R-5 | full bloom | leaf and stem | Pseudopeziza | 9 | |
| Clone R-5 | full bloom | leaf and stem | healthy | 3 | |
| Vernal | prebud | leaf and stem | Leptosphaerulina | 0 | |
| Clone R-5 | late bud | leaf and stem | Leptosphaerulina | 31 | |
| Clone R-5 | late bud | leaf and stem | Leptosphaerulina | 85 | |
| Clone R-5 | late bud | leaf and stem | healthy | 0 | |
| Buffalo | prebud | leaf and stem | Stemphylium | 30–45 | |
| Buffalo | prebud | leaf and stem | healthy | 0 | |
| Ranger | 1/10 bloom | leaf and stem | Yellow Mosaic virus | 30 | |
| Ranger | 1/10 bloom | leaf and stem | Yellow Mosaic virus | 33 | |
| Ranger | 1/10 bloom | leaf and stem | healthy | 19 | |
| Clone R-5 | ½ bloom | leaf and stem | Yellow Mosaic virus | 0 | |
| not stated | hay stage | leaf and stem | Foggy—1524 m ASL | 99 | [18] |
| not stated | hay stage | leaf and stem | Clear—610 m ASL | 32 | |
| not stated | hay stage | commercial meal | not stated | Usu. <100 | GO Kohler cited by [18] |
| Atlantic | vegetative | leaves | Ascochyta imperfecta | 132–542 | [95] |
| Atlantic | vegetative | leaves | Colletotrichum trifolii | 76 | |
| Atlantic | vegetative | leaves | Uromyces | 115 | |
| Atlantic | vegetative | Stem base | Cylindrocladium scoparium | 88 | |
| Atlantic | vegetative | roots | Cylindrocladium scoparium | 247–362 | |
| Atlantic | vegetative | leaves | Cylindrocladium scoparium | 0 | |
| Atlantic | vegetative | leaves | Xanthomonas alalfae | 22–40 | |
| not stated | sum.-autumn | tops | not stated | 25–190 | [26] |
| not stated | autumn grazed | tops | not stated | 51–157 | [27] |
| Wairau | autumn grazed | tops | not stated | 66–172 | [28] |
| Wairau | basal bud | tops | Blue green aphid + fungicide | 25 | [96] |
| Wairau | basal bud | tops | Pea aphid + fungicide | 13 | |
| Wairau | basal bud | tops | +fungicide | 2 | |
| Wairau | prebud | tops | Aphid infestation | 90 | |
| Wairau | prebud | tops | +aphicide | 23 | |
| not stated | not stated | leaf and stem | diseased | 0–159 | [97] (NSW survey) |
| not stated | not stated | leaf and stem | healthy | 0–19 | |
| various | @ 60 d intervals | leaf | not stated; coumestrol high after humid weather | 0–150 | [98] (NSW survey) |
| various | @ 60 d intervals | stem | not stated; coumestrol high after humid weather | 0–112 | |
| CUF 101 | @ 7 d intervals over spring, summer, aut. | leaf | rated 1–8; severity related to coumestrol | 0–150 | [98] |
| CUF 101 | stem | rated 1–8; severity related to coumestrol | 0–75 | ||
| not stated | summer/autumn | Pseudo., Lepto. | 100–350 | [99] (France survey) | |
| not stated | vege. to mature | whole tops | not stated | 15–225 | [100] |
| not stated | not stated | haylage | not stated | 32 | [29] |
| Pasture/Feed | Coumestan Concentration | Significant Results | Reference |
|---|---|---|---|
| Lambing studies | |||
| white clover vs. grass (Columbia premiparous and 2YO ewes over 3 year) | Clover positive to mouse uterine weight assay | 3% less lambs/ewe on clover cf. grass. Oestrous delayed; 41% conceived at 1st service cf. 66% for ewes on grass | [127] |
| lucerne vs. grass, white clover (3128 ewes over 3 year) | not assessed | 11% less lambs/ewe on lucerne due mainly to less multiple births | [128] |
| lucerne vs. grass, white clover (900 adult Border Leicester x Corriedale ewes over 10 weeks) | not assessed | 11% less lambs/ewe, and 2.65 cf. 0.3% barren, lucerne v grass | [26] |
| lucerne vs. grass, white clover (800 adult Border Leicester x Corriedale ewes over 7 weeks) | 60–150 mg/kg coumestrol (+0–40 mg/kg 4’-methyl–coumestrol) | 12% less lambs/ewe, and 3.0 cf. 1.0% barren, lucerne v grass | |
| lucerne vs. grass, white clover (Coopworth adult ewes, 2 year) | 51–104 mg/kg coumestrol (+9–91 mg/kg 4’-methyl–coumestrol) | 32% less lambs, lucerne v grass clover; 28% decrease in multiple births | [27] |
| lucerne vs. grass, white clover (Romney Marsh adult ewes, 2 year) | 82–157 mg/kg coumestrol (+41–154 mg/kg 4’-methyl–coumestrol) | 19% less lambs, lucernce v grass clover; 17% decrease in multiple births | |
| lucerne vs. grass, sub. clover (1800 Merino and crossbred ewes, mixed ages over 3 year) | not assessed | 8.5% less lambs, lucerne v grass clover. Fertile ewe % nsd. | [129] |
| M. falcata grazed then fed as silage vs. grass silage (34 ewes, 14 weeks) | 0—60 mg/kg coumestrol | conception and lambing both nsd; ewes conceived 5 days later on lucerne. | [91] |
| Case studies, uterus and ovulation observations | |||
| lucerne vs. grass, white clover | 66–172 mg/kg coumestrol (+33–145 mg/kg 4’-methyl–coumestrol) | ovulation depressed 29% following consumption during last half of oestrous cycle | [28] |
| varied lucerne treatments (1750 ewes over 2 year) | up to 600 mg/kg coumestrol in leaf. Fed coumestrol doses of 0–100 mg/kg | ovulation depressed 34%; lambing 14.6 %. dose linearly related to no. of ovulations from 1.44 to 0.98 | [51] |
| primiparous ewes | up to 350 mg/kg coumestrol. Summary of case studies, all involving diseased stands. | 60% barren at 45 days post insemination | [99] |
| primiparous ewes | >10% aborted 5 months into pregnancy | ||
| adult ewes | 5% aborted 5 months into pregnancy | ||
| lucerne vs. coumestrol-free diet (56 ewes over 10 months) | 25–30 mg/kg coumestrol | 43% lucerne ewes had macroscopic changes in the cervix and uterus | [119] |
| lucerne ad lib, day -7 to day 17 vs. maintenance diet of faba, oat hull pellet. 70 AI’d ewes | not assessed | 21% less foetuses/ewe, lucerne v pellets—nsd. lucerne ewes had less multiple ovulations (0.15 cf. 0.26)s | [130] |
| ‘flushing’ studies, viz. lucerne compared with low quality feed | |||
| lucerne vs. senescent grass, clover pasture | not assessed | lucerne ewes had greater multiple ovulations (0.36 cf. 0.27) | [131] |
| Lucerne vs. senescent pasture (300 ewes over 2 months) | not assessed | 19% more lambs/ewe, lucerne v senescent pasture. Barren ewes %—nsd | [132] |
| Species | Diet and Intake | Plasma Coumestrol | Plasma Methoxy-Coumestrol | Reference | ||
|---|---|---|---|---|---|---|
| Form | Concentration (µg/L) | Form | Concentration (µg/L) | |||
| Ewes (n = 5) | medic hay (M. littoralis) containing 300 mg/kg coumestrol and 340 mg/kg methoxy-coumestrol. Intake: 146 mg coumestrol and 124 mg 4’-methoxy-coumestrol/d | free | 5–7 | free | not detected | [23] |
| conjugate | 12–40 | conjugate | not detected | |||
| Ewes | not stated | free | 1.0–3.1 | [134] | ||
| sulphate conjugate | 1.8–5.0 | |||||
| glucuronide conjugate | 6.1–7.0 | |||||
| Ewes | fed 514 mg coumestrol/d for 16 d | free | 3.7 | [25] | ||
| conjugate | 14.5 | |||||
| fed 952 mg coumestrol/d for 16 d | free | 8.1 | ||||
| conjugate | 28.1 | |||||
| Goats | lucerne hay. 12 mg coumestrol/head/d | free | 2–3.9 | [142] | ||
| Mares—(Thoroughbred & Holstein, n = 16, 6–11YO, 540kg LW) | lucerne clover haylage (5–8 kg/d), concentrate + pasture and hay. Haylage contained 3 mg/kg coumestrol and 10 mg/kg methoxy-coumestrol | free | 0.03–0.24 1 | free | 0.06–0.18 1 | [29] |
| conjugate | 0.32–1.07 1 | conjugate | 0.45–1.04 1 | |||
© 2016 by the author; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Reed, K.F.M. Fertility of Herbivores Consuming Phytoestrogen-containing Medicago and Trifolium Species. Agriculture 2016, 6, 35. https://doi.org/10.3390/agriculture6030035
Reed KFM. Fertility of Herbivores Consuming Phytoestrogen-containing Medicago and Trifolium Species. Agriculture. 2016; 6(3):35. https://doi.org/10.3390/agriculture6030035
Chicago/Turabian StyleReed, K. F. M. 2016. "Fertility of Herbivores Consuming Phytoestrogen-containing Medicago and Trifolium Species" Agriculture 6, no. 3: 35. https://doi.org/10.3390/agriculture6030035
APA StyleReed, K. F. M. (2016). Fertility of Herbivores Consuming Phytoestrogen-containing Medicago and Trifolium Species. Agriculture, 6(3), 35. https://doi.org/10.3390/agriculture6030035